
Molecular Characterization of Secreted Factors and Extracellular Vesicles-Embedded miRNAs from Bone Marrow-Derived Mesenchymal Stromal Cells in Presence of Synovial Fluid from Osteoarthritis Patients

 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Synovial Fluid Collection
2.2. ELISA Characterization of Pooled SF
2.3. Bone Marrow Collection, BMSCs Isolation and Expansion
2.4. Flow Cytometry Characterization of SF-Treated BMSCs
2.5. ELISA Characterization of SF-Treated BMSCs Secretome
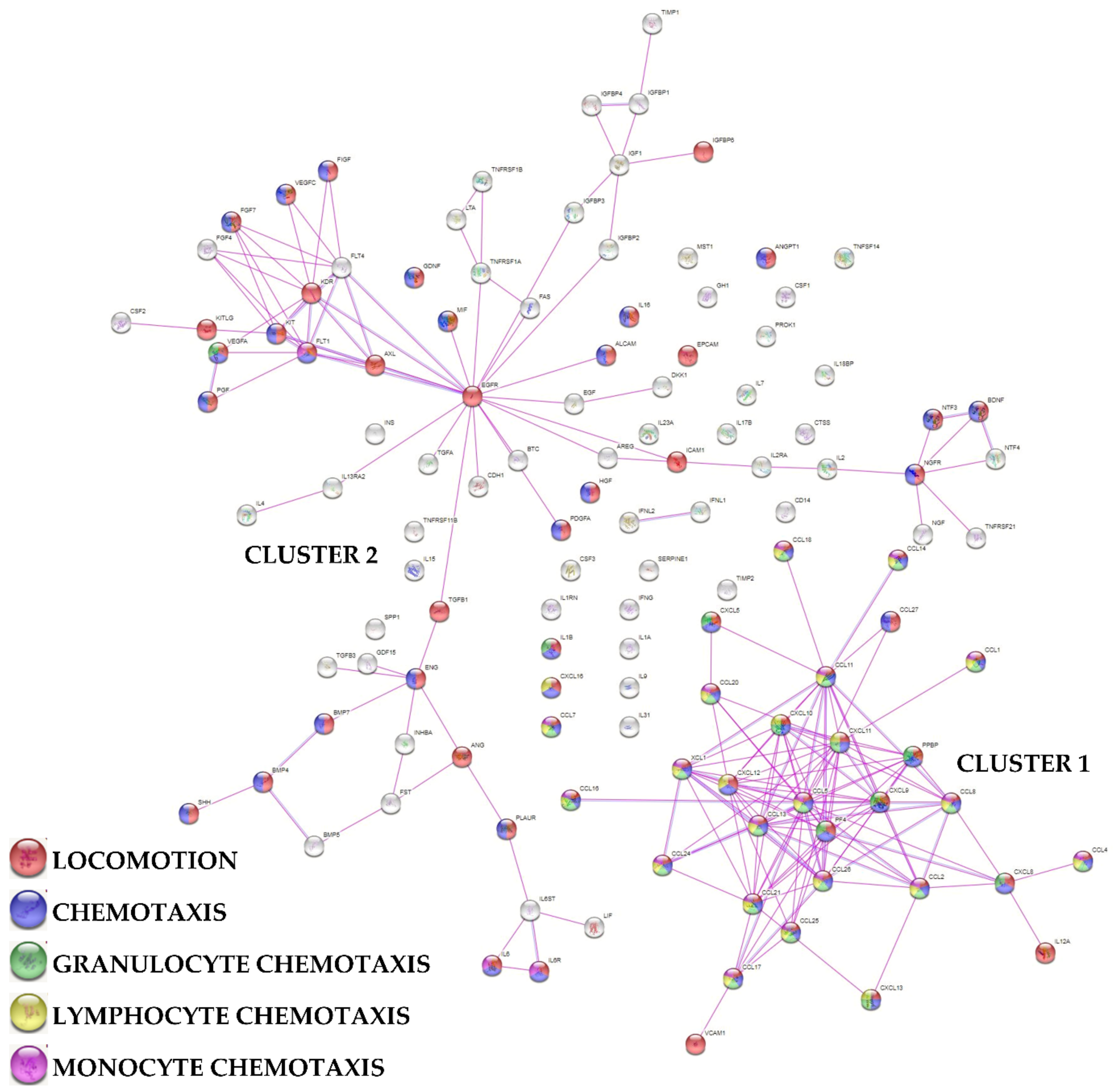
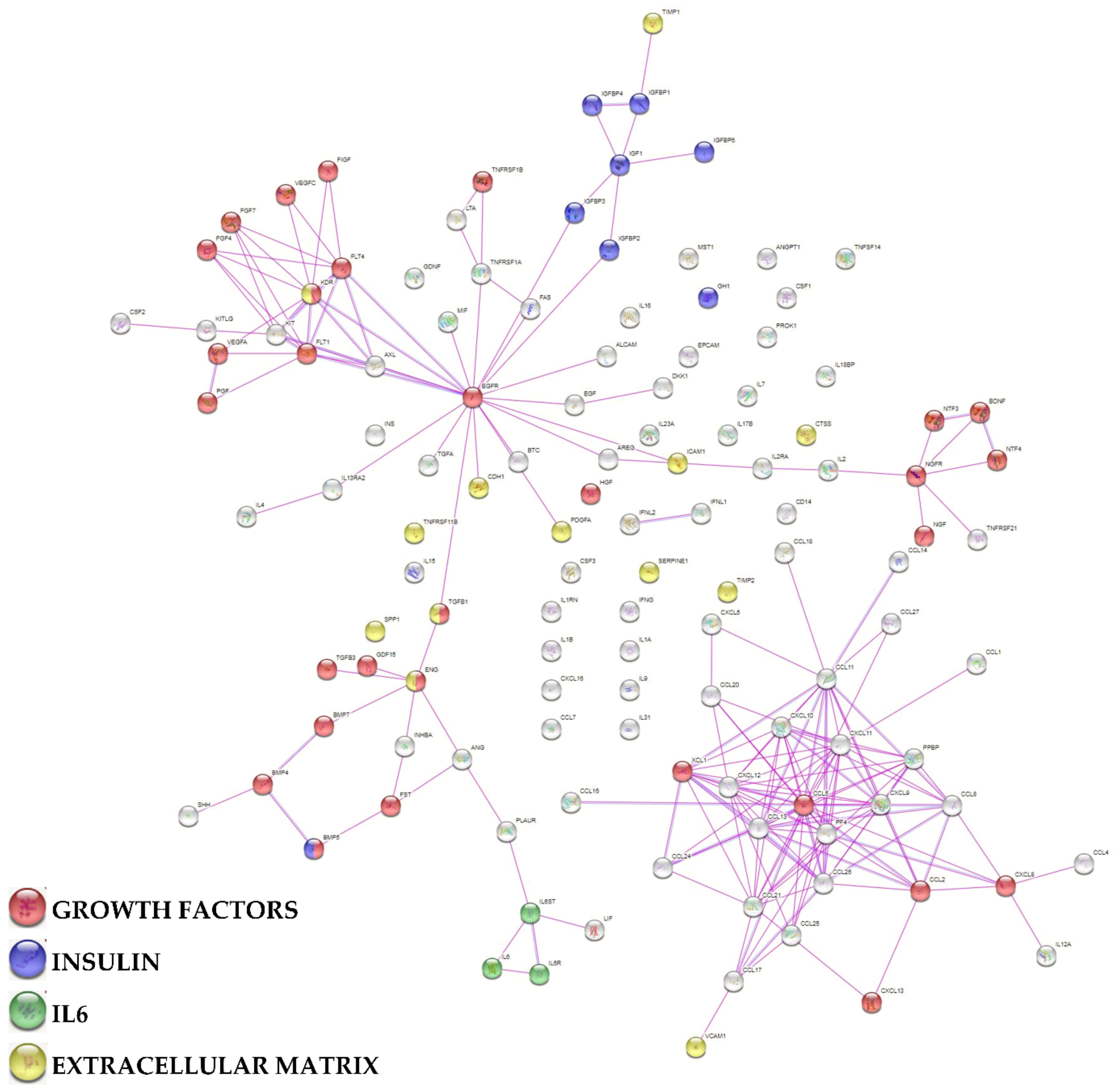
2.6. Protein-Protein Interaction Network
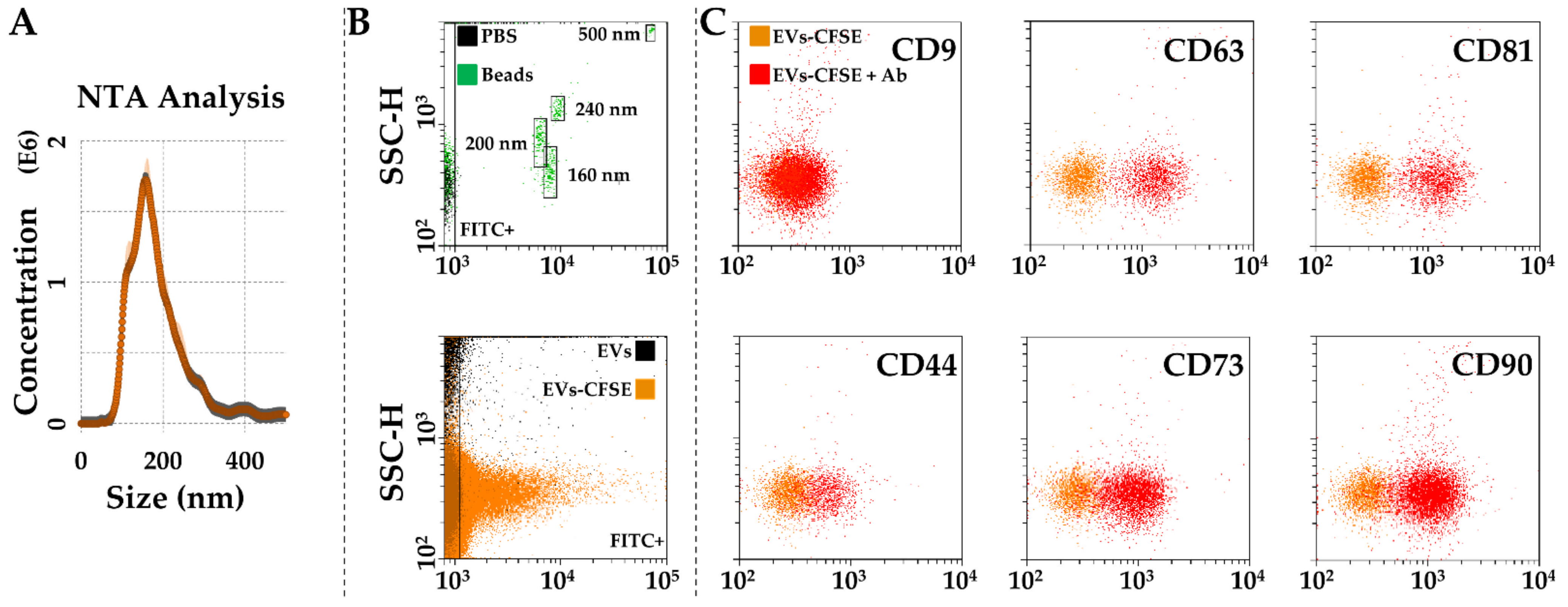
2.7. Characterization of EVs in SF-Treated BMSCs Secretomes
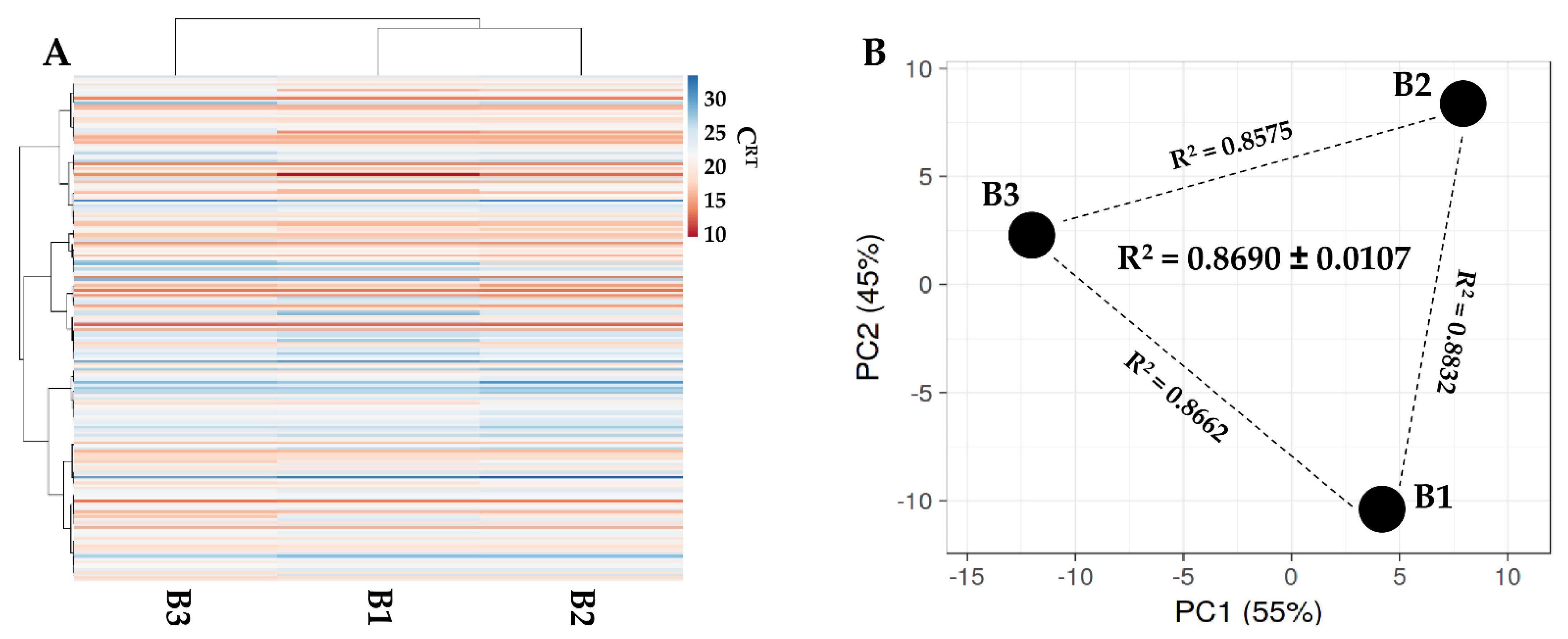
2.8. Total RNA Isolation from EVs and miRNAs Quantification
2.9. Identification of miRNAs Target
2.10. Statistical and Computational Analyses
3. Results
3.1. SF Characterization
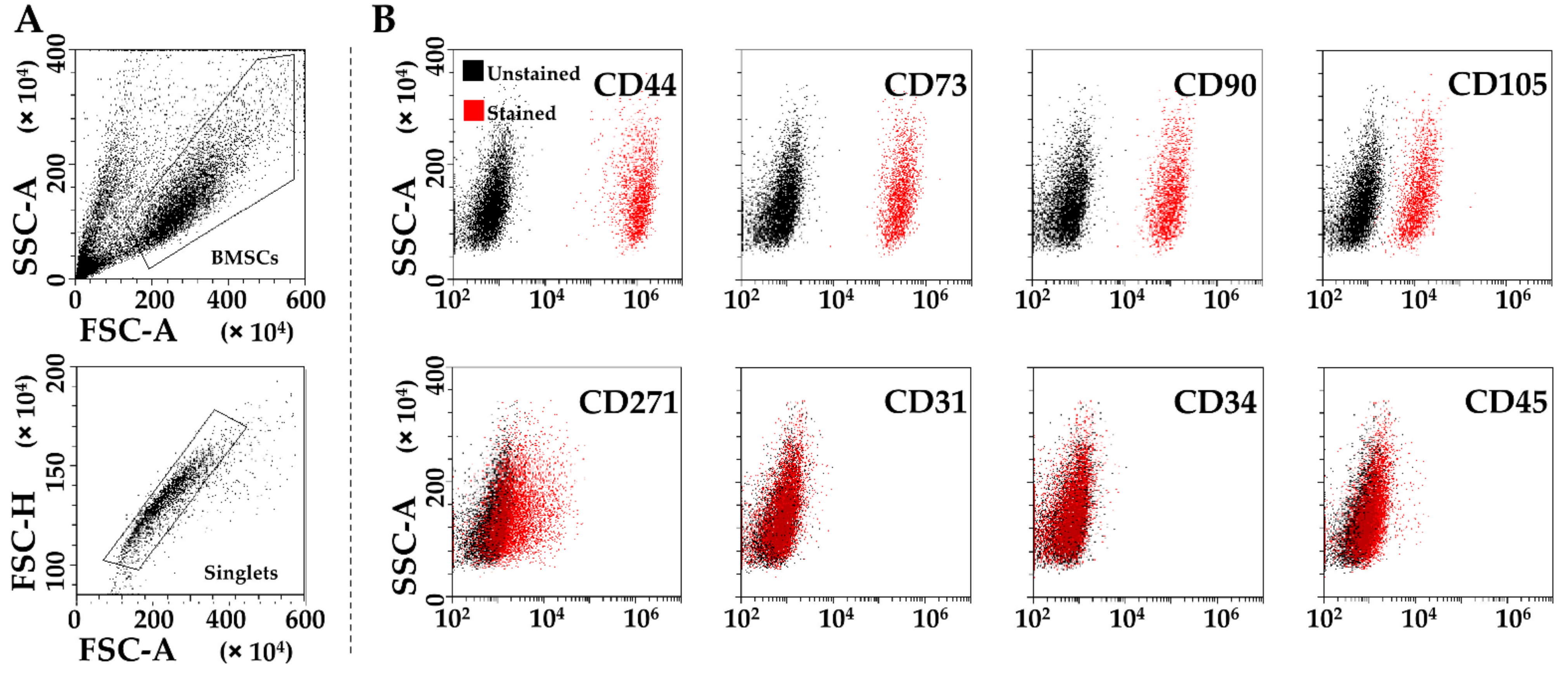
3.2. Immunophenotype of SF-Treated BMSCs
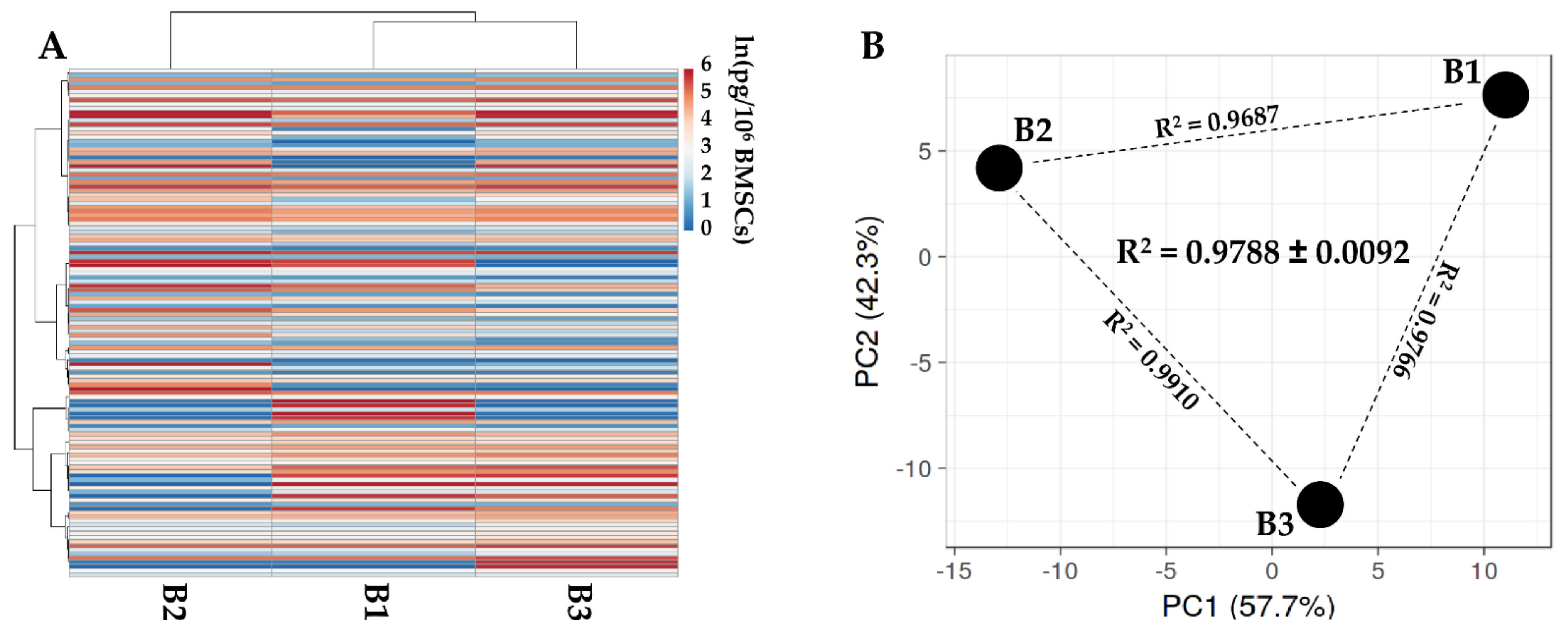
3.3. Characterization of SF-Treated BMSCs Secreted Factors
3.4. Characterization of SF-Treated BMSC-EVs
3.5. Identification of EV-Embedded miRNAs
3.6. EV-miRNAs Effect on OA-Related Molecules and Cell Types
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hunter, D.J.; Bierma-Zeinstra, S. Osteoarthritis. Lancet 2019, 393, 1745–1759. [Google Scholar] [CrossRef]
- Kleemann, R.U.; Krocker, D.; Cedraro, A.; Tuischer, J.; Duda, G.N. Altered cartilage mechanics and histology in knee osteoarthritis: Relation to clinical assessment (ICRS Grade). Osteoarthr. Cartil. 2005, 13, 958–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benito, M.J.; Veale, D.J.; FitzGerald, O.; van den Berg, W.B.; Bresnihan, B. Synovial tissue inflammation in early and late osteoarthritis. Ann. Rheum. Dis. 2005, 64, 1263–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martel-Pelletier, J.; Barr, A.J.; Cicuttini, F.M.; Conaghan, P.G.; Cooper, C.; Goldring, M.B.; Goldring, S.R.; Jones, G.; Teichtahl, A.J.; Pelletier, J. Osteoarthritis. Nat. Rev. Dis. Primers 2016, 2, 16072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, A.; Hilkens, C.M.U. Synovial Macrophages in Osteoarthritis: The Key to Understanding Pathogenesis? Front. Immunol. 2021, 12, 678757. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Luo, W.; Zhu, S.; Lei, G. T Cells in Osteoarthritis: Alterations and Beyond. Front. Immunol. 2017, 8, 356. [Google Scholar] [CrossRef] [Green Version]
- Kraus, V.B.; McDaniel, G.; Huebner, J.L.; Stabler, T.V.; Pieper, C.F.; Shipes, S.W.; Petry, N.A.; Low, P.S.; Shen, J.; McNearney, T.A.; et al. Direct in vivo evidence of activated macrophages in human osteoarthritis. Osteoarthr. Cartil. 2016, 24, 1613–1621. [Google Scholar] [CrossRef] [Green Version]
- Daghestani, H.N.; Pieper, C.F.; Kraus, V.B. Soluble macrophage biomarkers indicate inflammatory phenotypes in patients with knee osteoarthritis. Arthritis Rheumatol. 2015, 67, 956–965. [Google Scholar] [CrossRef] [Green Version]
- Huo, L.W.; Ye, Y.L.; Wang, G.W.; Ye, Y.G. Fractalkine (CX3CL1): A biomarker reflecting symptomatic severity in patients with knee osteoarthritis. J. Investig. Med. 2015, 63, 626–631. [Google Scholar] [CrossRef]
- Dolganiuc, A.; Stăvaru, C.; Anghel, M.; Georgescu, E.; Chichoş, B.; Olinescu, A. Shift toward T lymphocytes with Th1 and Tc1 cytokine-secterion profile in the joints of patients with osteoarthritis. Roum. Arch. Microbiol. Immunol. 1999, 58, 249–258. [Google Scholar]
- Haynes, M.K.; Hume, E.L.; Smith, J.B. Phenotypic characterization of inflammatory cells from osteoarthritic synovium and synovial fluids. Clin. Immunol. 2002, 105, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Grässel, S.; Muschter, D. Recent advances in the treatment of osteoarthritis. F1000Res. 2020, 9, 325. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhang, J.; Xu, H.; Lin, Z.; Chang, H.; Liu, W.; Kong, L. Mesenchymal stem cells in knee osteoarthritis treatment: A systematic review and meta-analysis. J. Orthop. Translat. 2020, 24, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Lopa, S.; Colombini, A.; Moretti, M.; de Girolamo, L. Injective mesenchymal stem cell-based treatments for knee osteoarthritis: From mechanisms of action to current clinical evidences. Knee Surg. Sports Traumatol. Arthrosc. 2019, 27, 2003–2020. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.; Andriolo, L.; Boffa, A.; Di Martino, A.; Reale, D.; Vara, G.; Miceli, M.; Cavallo, C.; Grigolo, B.; Zaffagnini, S.; et al. Prospective double-blind randomised controlled trial protocol comparing bone marrow aspirate concentrate intra-articular injection combined with subchondral injection versus intra-articular injection alone for the treatment of symptomatic knee osteoarthritis. BMJ Open 2022, 12, e062632. [Google Scholar] [CrossRef]
- Heidari, N.; Borg, T.; Olgiati, S.; Slevin, M.; Danovi, A.; Fish, B.; Wilson, A.; Noorani, A. Microfragmented Adipose Tissue Injection (MFAT) May Be a Solution to the Rationing of Total Knee Replacement: A Prospective, Gender-Bias Mitigated, Reproducible Analysis at Two Years. Stem Cells Int. 2021, 2021, 9921015. [Google Scholar] [CrossRef]
- Zhang, S.; Xu, H.; He, B.; Fan, M.; Xiao, M.; Zhang, J.; Chen, D.; Tong, P.; Mao, Q. Mid-term prognosis of the stromal vascular fraction for knee osteoarthritis: A minimum 5-year follow-up study. Stem Cell Res. Ther. 2022, 13, 105. [Google Scholar] [CrossRef]
- Zhu, C.; Wu, W.; Qu, X. Mesenchymal stem cells in osteoarthritis therapy: A review. Am. J. Transl. Res. 2021, 13, 448–461. [Google Scholar]
- Van Buul, G.M.; Villafuertes, E.; Bos, P.K.; Waarsing, J.H.; Kops, N.; Narcisi, R.; Weinans, H.; Verhaar, J.A.N.; Bernsen, M.R.; van Osch, G.J.V.M. Mesenchymal stem cells secrete factors that inhibit inflammatory processes in short-term osteoarthritic synovium and cartilage explant culture. Osteoarthr. Cartil. 2012, 20, 1186–1196. [Google Scholar] [CrossRef] [Green Version]
- Cosenza, S.; Ruiz, M.; Toupet, K.; Jorgensen, C.; Noël, D. Mesenchymal stem cells derived exosomes and microparticles protect cartilage and bone from degradation in osteoarthritis. Sci. Rep. 2017, 7, 16214. [Google Scholar] [CrossRef] [Green Version]
- González-González, A.; García-Sánchez, D.; Dotta, M.; Rodríguez-Rey, J.C.; Pérez-Campo, F.M. Mesenchymal stem cells secretome: The cornerstone of cell-free regenerative medicine. World J. Stem Cells 2020, 12, 1529–1552. [Google Scholar] [CrossRef] [PubMed]
- Baberg, F.; Geyh, S.; Waldera-Lupa, D.; Stefanski, A.; Zilkens, C.; Haas, R.; Schroeder, T.; Stühler, K. Secretome analysis of human bone marrow derived mesenchymal stromal cells. Biochim. Biophys. Acta Proteins Proteom. 2019, 1867, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Wangler, S.; Kamali, A.; Wapp, C.; Wuertz-Kozak, K.; Häckel, S.; Fortes, C.; Benneker, L.M.; Haglund, L.; Richards, R.G.; Alini, M.; et al. Uncovering the secretome of mesenchymal stromal cells exposed to healthy, traumatic, and degenerative intervertebral discs: A proteomic analysis. Stem Cell Res. Ther. 2021, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Vega, A.; Martín-Ferrero, M.A.; Del Canto, F.; Alberca, M.; García, V.; Munar, A.; Orozco, L.; Soler, R.; Fuertes, J.J.; Huguet, M.; et al. Treatment of Knee Osteoarthritis with Allogeneic Bone Marrow Mesenchymal Stem Cells: A Randomized Controlled Trial. Transplantation 2015, 99, 1681–1690. [Google Scholar] [CrossRef]
- Al-Najar, M.; Khalil, H.; Al-Ajlouni, J.; Al-Antary, E.; Hamdan, M.; Rahmeh, R.; Alhattab, D.; Samara, O.; Yasin, M.; Al Abdullah, A.; et al. Intra-articular injection of expanded autologous bone marrow mesenchymal cells in moderate and severe knee osteoarthritis is safe: A phase I/II study. J. Orthop. Surg. Res. 2017, 12, 190. [Google Scholar] [CrossRef] [Green Version]
- Davatchi, F.; Abdollahi, B.S.; Mohyeddin, M.; Shahram, F.; Nikbin, B. Mesenchymal stem cell therapy for knee osteoarthritis. Preliminary report of four patients. Int. J. Rheum. Dis. 2011, 14, 211–215. [Google Scholar] [CrossRef]
- Kraus, V.B.; Stabler, T.V.; Kong, S.Y.; Varju, G.; McDaniel, G. Measurement of synovial fluid volume using urea. Osteoarthr. Cartil. 2007, 15, 1217–1220. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Abram, F.; Pelletier, J.; Raynauld, J.; Dorais, M.; d’Anjou, M.; Martel-Pelletier, J. Fully automated system for the quantification of human osteoarthritic knee joint effusion volume using magnetic resonance imaging. Arthritis Res. Ther. 2010, 12, R173. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Lin, Y.; Li, J.; Huang, K.; Shrestha, S.; Hong, H.; Tang, Y.; Chen, Y.; Jin, C.; Yu, Y.; et al. miRTarBase 2020: Updates to the experimentally validated microRNA-target interaction database. Nucleic Acids Res. 2020, 48, D148–D154. [Google Scholar] [CrossRef] [Green Version]
- Akoglu, H. User’s guide to correlation coefficients. Turk. J. Emerg. Med. 2018, 18, 91–93. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Barilani, M.; Banfi, F.; Sironi, S.; Ragni, E.; Guillaumin, S.; Polveraccio, F.; Rosso, L.; Moro, M.; Astori, G.; Pozzobon, M.; et al. Low-affinity Nerve Growth Factor Receptor (CD271) Heterogeneous Expression in Adult and Fetal Mesenchymal Stromal Cells. Sci. Rep. 2018, 8, 9321. [Google Scholar] [CrossRef] [PubMed]
- Barilani, M.; Peli, V.; Cherubini, A.; Dossena, M.; Dolo, V.; Lazzari, L. NG2 as an Identity and Quality Marker of Mesenchymal Stem Cell Extracellular Vesicles. Cells 2019, 8, 1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toh, W.S.; Lai, R.C.; Hui, J.H.P.; Lim, S.K. MSC exosome as a cell-free MSC therapy for cartilage regeneration: Implications for osteoarthritis treatment. Semin. Cell Dev. Biol. 2017, 67, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Chevillet, J.R.; Kang, Q.; Ruf, I.K.; Briggs, H.A.; Vojtech, L.N.; Hughes, S.M.; Cheng, H.H.; Arroyo, J.D.; Meredith, E.K.; Gallichotte, E.N.; et al. Quantitative and stoichiometric analysis of the microRNA content of exosomes. Proc. Natl. Acad. Sci. USA 2014, 111, 14888–14893. [Google Scholar] [CrossRef] [Green Version]
- Schopman, N.C.T.; Heynen, S.; Haasnoot, J.; Berkhout, B. A miRNA-tRNA mix-up: tRNA origin of proposed miRNA. RNA Biol. 2010, 7, 573–576. [Google Scholar] [CrossRef]
- Chou, C.; Jain, V.; Gibson, J.; Attarian, D.E.; Haraden, C.A.; Yohn, C.B.; Laberge, R.; Gregory, S.; Kraus, V.B. Synovial cell cross-talk with cartilage plays a major role in the pathogenesis of osteoarthritis. Sci. Rep. 2020, 10, 10868. [Google Scholar] [CrossRef]
- Endisha, H.; Rockel, J.; Jurisica, I.; Kapoor, M. The complex landscape of microRNAs in articular cartilage: Biology, pathology, and therapeutic targets. JCI Insight 2018, 3, e121630. [Google Scholar] [CrossRef] [Green Version]
- Tavallaee, G.; Rockel, J.S.; Lively, S.; Kapoor, M. MicroRNAs in Synovial Pathology Associated with Osteoarthritis. Front. Med. 2020, 7, 376. [Google Scholar] [CrossRef]
- Xu, S.J.; Hu, H.T.; Li, H.L.; Chang, S. The Role of miRNAs in Immune Cell Development, Immune Cell Activation, and Tumor Immunity: With a Focus on Macrophages and Natural Killer Cells. Cells 2019, 8, 1140. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Galán, A.; Fernández-Messina, L.; Sánchez-Madrid, F. Control of Immunoregulatory Molecules by miRNAs in T Cell Activation. Front. Immunol. 2018, 9, 2148. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Zhu, S.; He, H.; Yu, X.; Xu, Y.; He, C. Mesenchymal stromal cell-based therapy for cartilage regeneration in knee osteoarthritis. Stem Cell Res. Ther. 2022, 13, 14. [Google Scholar] [CrossRef] [PubMed]
- Orozco, L.; Munar, A.; Soler, R.; Alberca, M.; Soler, F.; Huguet, M.; Sentís, J.; Sánchez, A.; García-Sancho, J. Treatment of knee osteoarthritis with autologous mesenchymal stem cells: A pilot study. Transplantation 2013, 95, 1535–1541. [Google Scholar] [CrossRef] [PubMed]
- Soler, R.; Orozco, L.; Munar, A.; Huguet, M.; López, R.; Vives, J.; Coll, R.; Codinach, M.; Garcia-Lopez, J. Final results of a phase I-II trial using ex vivo expanded autologous Mesenchymal Stromal Cells for the treatment of osteoarthritis of the knee confirming safety and suggesting cartilage regeneration. Knee 2016, 23, 647–654. [Google Scholar] [CrossRef]
- Emadedin, M.; Labibzadeh, N.; Liastani, M.G.; Karimi, A.; Jaroughi, N.; Bolurieh, T.; Hosseini, S.; Baharvand, H.; Aghdami, N. Intra-articular implantation of autologous bone marrow-derived mesenchymal stromal cells to treat knee osteoarthritis: A randomized, triple-blind, placebo-controlled phase 1/2 clinical trial. Cytotherapy 2018, 20, 1238–1246. [Google Scholar] [CrossRef]
- Bastos, R.; Mathias, M.; Andrade, R.; Bastos, R.; Balduino, A.; Schott, V.; Rodeo, S.; Espregueira-Mendes, J. Intra-articular injections of expanded mesenchymal stem cells with and without addition of platelet-rich plasma are safe and effective for knee osteoarthritis. Knee Surg. Sports Traumatol. Arthrosc. 2018, 26, 3342–3350. [Google Scholar] [CrossRef]
- Chahal, I.; Gómez-Aristizábal, A.; Shestopaloff, K.; Bhatt, S.; Chaboureau, A.; Fazio, A.; Chisholm, J.; Weston, A.; Chiovitti, J.; Keating, A.; et al. Bone Marrow Mesenchymal Stromal Cell Treatment in Patients with Osteoarthritis Results in Overall Improvement in Pain and Symptoms and Reduces Synovial Inflammation. Stem Cells Transl. Med. 2019, 8, 746–757. [Google Scholar] [CrossRef] [Green Version]
- Hernigou, P.; Delambre, J.; Quiennec, S.; Poignard, A. Human bone marrow mesenchymal stem cell injection in subchondral lesions of knee osteoarthritis: A prospective randomized study versus contralateral arthroplasty at a mean fifteen year follow-up. Int. Orthop. 2021, 45, 365–373. [Google Scholar] [CrossRef]
- Zheng, Y.H.; Deng, Y.Y.; Lai, W.; Zheng, S.Y.; Bian, H.N.; Liu, Z.A.; Huang, Z.H.; Sun, C.W.; Li, H.H.; Luo, H.M.; et al. Effect of bone marrow mesenchymal stem cells on the polarization of macrophages. Mol. Med. Rep. 2018, 17, 4449–4459. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, X.; Cao, W.; Shi, Y. Plasticity of mesenchymal stem cells in immunomodulation: Pathological and therapeutic implications. Nat. Immunol. 2014, 15, 1009–1016. [Google Scholar] [CrossRef]
- Luz-Crawford, P.; Kurte, M.; Bravo-Alegría, J.; Contreras, R.; Nova-Lamperti, E.; Tejedor, G.; Noël, D.; Jorgensen, C.; Figueroa, F.; Djouad, F.; et al. Mesenchymal stem cells generate a CD4+CD25+Foxp3+ regulatory T cell population during the differentiation process of Th1 and Th17 cells. Stem Cell Res. Ther. 2013, 4, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kean, T.J.; Lin, P.; Caplan, A.I.; Dennis, J.E. MSCs: Delivery Routes and Engraftment, Cell-Targeting Strategies, and Immune Modulation. Stem Cells Int. 2013, 2013, 732742. [Google Scholar] [CrossRef] [PubMed]
- Luz-Crawford, P.; Djouad, F.; Toupet, K.; Bony, C.; Franquesa, M.; Hoogduijn, M.J.; Jorgensen, C.; Noël, D. Mesenchymal Stem Cell-Derived Interleukin 1 Receptor Antagonist Promotes Macrophage Polarization and Inhibits B Cell Differentiation. Stem Cells 2016, 34, 483–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melief, S.M.; Schrama, E.; Brugman, M.H.; Tiemessen, M.M.; Hoogduijn, M.J.; Fibbe, W.E.; Roelofs, H. Multipotent stromal cells induce human regulatory T cells through a novel pathway involving skewing of monocytes toward anti-inflammatory macrophages. Stem Cells 2013, 31, 1980–1991. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Huang, S.; Yuan, X.; Liang, J.; Xu, R.; Yao, G.; Feng, X.; Sun, L. The regulation of the Treg/Th17 balance by mesenchymal stem cells in human systemic lupus erythematosus. Cell Mol. Immunol. 2017, 14, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.; Zhang, X.M.; Joshi, R.N.; Iqbal, S.; Wahlund, C.; Gabrielsson, S.; Harris, R.A.; Tegnér, J. Human macrophages induce CD4(+)Foxp3(+) regulatory T cells via binding and re-release of TGF-β. Immunol. Cell Biol. 2016, 94, 747–762. [Google Scholar] [CrossRef] [Green Version]
- Di Nicola, M.; Carlo-Stella, C.; Magni, M.; Milanesi, M.; Longoni, P.D.; Matteucci, P.; Grisanti, S.; Gianni, A.M. Human bone marrow stromal cells suppress T-lymphocyte proliferation induced by cellular or nonspecific mitogenic stimuli. Blood 2002, 99, 3838–3843. [Google Scholar] [CrossRef]
- Aoki, C.A.; Borchers, A.T.; Li, M.; Flavell, R.A.; Bowlus, C.L.; Ansari, A.; Gershwin, M.E. Transforming growth factor beta (TGF-beta) and autoimmunity. Autoimmun. Rev. 2005, 4, 450–459. [Google Scholar] [CrossRef]
- Zheng, S.; Huang, K.; Xia, W.; Shi, J.; Liu, Q.; Zhang, X.; Li, G.; Chen, J.; Wang, T.; Chen, X.; et al. Mesenchymal Stromal Cells Rapidly Suppress TCR Signaling-Mediated Cytokine Transcription in Activated T Cells Through the ICAM-1/CD43 Interaction. Front. Immunol. 2021, 12, 609544. [Google Scholar] [CrossRef]
- Xu, Q.; Sun, X.; Shang, X.; Jiang, H. Association of CXCL12 levels in synovial fluid with the radiographic severity of knee osteoarthritis. J. Investig. Med. 2012, 60, 898–901. [Google Scholar] [CrossRef]
- Monasterio, G.; Castillo, F.; Rojas, L.; Cafferata, E.A.; Alvarez, C.; Carvajal, P.; Núñez, C.; Flores, G.; Díaz, W.; Vernal, R. Th1/Th17/Th22 immune response and their association with joint pain, imagenological bone loss, RANKL expression and osteoclast activity in temporomandibular joint osteoarthritis: A preliminary report. J. Oral Rehabil. 2018, 45, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Bollmann, M.; Lokau, J.; Garbers, C.; Bertrand, J. Interleukin-11—A new cytokine in osteoarthritis? Osteoarthr. Cartil. 2022, 30, S168–S169. [Google Scholar] [CrossRef]
- Fordham, J.B.; Naqvi, A.R.; Nares, S. miR-24 Regulates Macrophage Polarization and Plasticity. J. Clin. Cell Immunol. 2015, 6, 362. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Liu, Y.; Wang, J.; Zhang, M.; Wang, M. Cardioprotection of M2 macrophages-derived exosomal microRNA-24-3p/Tnfsf10 axis against myocardial injury after sepsis. Mol. Immunol. 2022, 141, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Fayyad-Kazan, H.; Hamade, E.; Rouas, R.; Najar, M.; Fayyad-Kazan, M.; El Zein, N.; ElDirani, R.; Hussein, N.; Fakhry, M.; Al-Akoum, C.; et al. Downregulation of microRNA-24 and -181 parallels the upregulation of IFN-γ secreted by activated human CD4 lymphocytes. Hum. Immunol. 2014, 75, 677–685. [Google Scholar] [CrossRef]
- Chandran, P.A.; Keller, A.; Weinmann, L.; Seida, A.A.; Braun, M.; Andreev, K.; Fischer, B.; Horn, E.; Schwinn, S.; Junker, M.; et al. The TGF-β-inducible miR-23a cluster attenuates IFN-γ levels and antigen-specific cytotoxicity in human CD8⁺ T cells. J. Leukoc. Biol. 2014, 96, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.-B.; Zhang, H.; Cai, T.-T.; Liu, Y.-N.; Ni, J.-J.; He, J.; Peng, J.-Y.; Chen, Q.-Y.; Mo, H.-Y.; Jun-Cui; et al. Exosomal miR-24-3p impedes T-cell function by targeting FGF11 and serves as a potential prognostic biomarker for nasopharyngeal carcinoma. J. Pathol. 2016, 240, 329–340. [Google Scholar] [CrossRef]
- Graff, J.W.; Dickson, A.M.; Clay, G.; McCaffrey, A.P.; Wilson, M.E. Identifying functional microRNAs in macrophages with polarized phenotypes. J. Biol. Chem. 2012, 287, 21816–21825. [Google Scholar] [CrossRef] [Green Version]
- Ragni, E.; Perucca Orfei, C.; De Luca, P.; Lugano, G.; Viganò, M.; Colombini, A.; Valli, F.; Zacchetti, D.; Bollati, V.; de Girolamo, L. Interaction with hyaluronan matrix and miRNA cargo as contributors for in vitro potential of mesenchymal stem cell-derived extracellular vesicles in a model of human osteoarthritic synoviocytes. Stem Cell Res. Ther. 2019, 10, 109. [Google Scholar] [CrossRef] [Green Version]
- De Lange-Brokaar, B.J.E.; Ioan-Facsinay, A.; van Osch, G.J.V.M.; Zuurmond, A.; Schoones, J.; Toes, R.E.M.; Huizinga, T.W.J.; Kloppenburg, M. Synovial inflammation, immune cells and their cytokines in osteoarthritis: A review. Osteoarthr. Cartil. 2012, 20, 1484–1499. [Google Scholar] [CrossRef] [Green Version]
- Brew, K.; Nagase, H. The tissue inhibitors of metalloproteinases (TIMPs): An ancient family with structural and functional diversity. Biochim. Biophys. Acta 2010, 1803, 55–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, A.J.; Curry, V.A.; Powell, E.K.; Cawston, T.E. The prevention of collagen breakdown in bovine nasal cartilage by TIMP, TIMP-2 and a low molecular weight synthetic inhibitor. Biochem. Biophys. Res. Commun. 1994, 201, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Sun, F.; Jiang, L.; Bao, J.; Wu, L. Paeoniflorin inhibits IL-1β-induced MMP secretion via the NF-κB pathway in chondrocytes. Exp. Ther. Med. 2018, 16, 1513–1519. [Google Scholar] [CrossRef] [PubMed]
- Qu, H.; Li, J.; Wu, L.; Chen, W. Trichostatin A increases the TIMP-1/MMP ratio to protect against osteoarthritis in an animal model of the disease. Mol. Med. Rep. 2016, 14, 2423–2430. [Google Scholar] [CrossRef] [Green Version]
- Blaney Davidson, E.N.; van der Kraan, P.M.; van den Berg, W.B. TGF-beta and osteoarthritis. Osteoarthr. Cartil. 2007, 15, 597–604. [Google Scholar] [CrossRef] [Green Version]
- Sanrattana, W.; Maas, C.; de Maat, S. SERPINs-From Trap to Treatment. Front. Med. 2019, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Martel-Pelletier, J.; Faure, M.P.; McCollum, R.; Mineau, F.; Cloutier, J.M.; Pelletier, J.P. Plasmin, plasminogen activators and inhibitor in human osteoarthritic cartilage. J. Rheumatol. 1991, 18, 1863–1871. [Google Scholar]
- Deng, Z.H.; Li, Y.S.; Gao, X.; Lei, G.H.; Huard, J. Bone morphogenetic proteins for articular cartilage regeneration. Osteoarthr. Cartil. 2018, 26, 1153–1161. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, T.; Cooper, G.M.; Gharaibeh, B.; Meszaros, L.B.; Li, G.; Usas, A.; Fu, F.H.; Huard, J. Cartilage repair in a rat model of osteoarthritis through intraarticular transplantation of muscle-derived stem cells expressing bone morphogenetic protein 4 and soluble Flt-1. Arthritis Rheum. 2009, 60, 1390–1405. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Zhang, X.; Zhu, J.; Pi, Y.; Hu, X.; Zhou, C.; Ao, Y. Nanoparticle delivery of the bone morphogenetic protein 4 gene to adipose-derived stem cells promotes articular cartilage repair in vitro and in vivo. Arthroscopy 2013, 29, 2001–2011.e2. [Google Scholar] [CrossRef]
- Tanaka, N.; Tsuno, H.; Ohashi, S.; Iwasawa, M.; Furukawa, H.; Kato, T.; Fukui, N. The attenuation of insulin-like growth factor signaling may be responsible for relative reduction in matrix synthesis in degenerated areas of osteoarthritic cartilage. BMC Musculoskelet Disord. 2021, 22, 231. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.A.; Scherb, M.B.; Lembke, L.A.; Buckwalter, J.A. Damage control mechanisms in articular cartilage: The role of the insulin-like growth factor I axis. Iowa Orthop. J. 2000, 20, 1–10. [Google Scholar] [PubMed]
- Pérez-García, S.; Carrión, M.; Gutiérrez-Cañas, I.; Villanueva-Romero, R.; Castro, D.; Martínez, C.; González-Álvaro, I.; Blanco, F.J.; Juarranz, Y.; Gomariz, R.P. Profile of Matrix-Remodeling Proteinases in Osteoarthritis: Impact of Fibronectin. Cells 2019, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Qian, X.; Ding, R. MiR-24-3p attenuates IL-1β-induced chondrocyte injury associated with osteoarthritis by targeting BCL2L12. J. Orthop. Surg. Res. 2021, 16, 371. [Google Scholar] [CrossRef]
- Meng, F.; Li, Z.; Zhang, Z.; Long, D.; He, A.; Liao, W. Microrna-193B modulates chondrogenesis and cartilage degeneration via histone deacetylase 3. Osteoarthr. Cartil. 2018, 26, S93–S94. [Google Scholar] [CrossRef] [Green Version]
- Kopańska, M.; Szala, D.; Czech, J.; Gabło, N.; Gargasz, K.; Trzeciak, M.; Zawlik, I.; Snela, S. MiRNA expression in the cartilage of patients with osteoarthritis. J. Orthop. Surg. Res. 2017, 12, 51. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Jin, E.; Kim, D.; Kim, K.Y.; Chun, C.; Jin, E. MicroRNA-222 regulates MMP-13 via targeting HDAC-4 during osteoarthritis pathogenesis. BBA Clin. 2014, 3, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.; Zhang, Z.; Chen, W.; Huang, G.; He, A.; Hou, C.; Long, Y.; Yang, Z.; Zhang, Z.; Liao, W. MicroRNA-320 regulates matrix metalloproteinase-13 expression in chondrogenesis and interleukin-1β-induced chondrocyte responses. Osteoarthr. Cartil. 2016, 24, 932–941. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TYPE | FACTOR | (pg/mL) | |
|---|---|---|---|
| GF | IGFBP4 | 248,050 | Insulin-like growth factor-binding protein 4 |
| REC | VCAM1 | 109,797 | Vascular cell adhesion protein 1 |
| CYT | ICAM2 | 109,414 | Intercellular adhesion molecule 2 |
| GF | IGFBP2 | 44,358 | Insulin-like growth factor-binding protein 2 |
| CYT | PLG | 42,376 | Plasminogen |
| GF | IGFBP3 | 42,150 | Insulin-like growth factor-binding protein 3 |
| CYT | SIGLEC5 | 32,371 | Sialic acid-binding Ig-like lectin 5 |
| GF | TGFB1 | 29,154 | Transforming growth factor beta-1 |
| INF | TIMP2 | 25,220 | Metalloproteinase inhibitor 2 |
| REC | SELL | 20,525 | L-selectin |
| INF | IL6R | 19,933 | Interleukin-6 receptor subunit alpha |
| GF | BMP4 | 19,550 | Bone morphogenetic protein 4 |
| INF | TNFRSF1A | 19,497 | Tumor necrosis factor receptor superfamily member 1A |
| GF | CSF1R | 19,117 | Macrophage colony-stimulating factor 1 receptor |
| CYT | IL6ST | 15,002 | Interleukin-6 receptor subunit beta |
| CHE | IFNL1 | 13,977 | Interferon lambda-1 |
| REC | TNFRSF17 | 12,355 | Tumor necrosis factor receptor superfamily member 17 |
| REC | PLAUR | 11,597 | Urokinase plasminogen activator surface receptor |
| INF | TNFRSF1B | 10,977 | Tumor necrosis factor receptor superfamily member 1B |
| pg/Million BMSCs | |||||||
|---|---|---|---|---|---|---|---|
| TYPE | FACTOR | B1 | B2 | B3 | MEAN | SD | FUNCTION |
| GF | IGFBP4 | 109,869 | 131,569 | 124,587 | 122,009 | 9045 | Insulin-like growth factor-binding protein 4 |
| GF | IGFBP3 | 84,091 | 117,198 | 102,025 | 101,105 | 13,532 | Insulin-like growth factor-binding protein 3 |
| GF | TGFB1 | 39,339 | 32,615 | 30,189 | 34,048 | 3870 | Transforming growth factor beta-1 |
| INF | TIMP2 | 22,096 | 28,331 | 30,462 | 26,963 | 3550 | Metalloproteinase inhibitor 2 |
| CHE | IFNL1 | 19,399 | 20,613 | 22,381 | 20,798 | 1225 | Interferon lambda-1 |
| CHE | IL9 | 1770 | 24,217 | 16,270 | 14,086 | 9293 | Interleukin-9 |
| INF | TIMP1 | 12,116 | 14,563 | 14,503 | 13,727 | 1140 | Metalloproteinase inhibitor 1 |
| GF | BMP4 | 7872 | 10,892 | 20,075 | 12,946 | 5190 | Bone morphogenetic protein 4 |
| CYT | SERPINE1 | 10,110 | 14,584 | 13,253 | 12,649 | 1876 | Plasminogen activator inhibitor 1 |
| GF | IGFBP2 | 6439 | 14,130 | 10,661 | 10,410 | 3145 | Insulin-like growth factor-binding protein 2 |
| GF | VEGFA | 7794 | 12,442 | 10,732 | 10,323 | 1919 | Vascular endothelial growth factor A |
| CHE | PF4 | 9652 | 10,111 | 8745 | 9503 | 567 | Platelet factor 4 |
| GF | IGFBP6 | 6466 | 10,314 | 9509 | 8763 | 1657 | Insulin-like growth factor-binding protein 6 |
| CHE | MIF | 3604 | 4526 | 4973 | 4368 | 570 | Macrophage migration inhibitory factor |
| REC | VCAM1 | 4131 | 5719 | 2852 | 4234 | 1173 | Vascular cell adhesion protein 1 |
| CYT | INHBA | 4344 | 4350 | 3770 | 4155 | 272 | Inhibin beta A chain |
| CHE | XCL1 | 2921 | 4047 | 5177 | 4048 | 921 | Lymphotactin |
| CHE | CCL27 | 4283 | 2370 | 3979 | 3544 | 840 | C-C motif chemokine 27 |
| CHE | CXCL16 | 2374 | 3787 | 3932 | 3364 | 702 | C-X-C motif chemokine 16 |
| INF | TNFRSF1A | 2640 | 3403 | 3410 | 3151 | 361 | Tumor necrosis factor receptor superfamily member 1A |
| CHE | MST1 | 3356 | 1822 | 2150 | 2443 | 659 | Hepatocyte growth factor-like protein |
| CHE | CCL26 | 2212 | 932 | 4108 | 2417 | 1305 | C-C motif chemokine 26 |
| CHE | CCL21 | 2552 | 3078 | 1594 | 2408 | 614 | C-C motif chemokine 21 |
| CHE | CXCL11 | 2165 | 3168 | 1466 | 2266 | 698 | C-X-C motif chemokine 11 |
| REC | PLAUR | 1515 | 2482 | 1641 | 1879 | 429 | Urokinase plasminogen activator surface receptor |
| CYT | ANGPT1 | 1553 | 2005 | 1837 | 1798 | 187 | Angiopoietin-1 |
| GF | BMP7 | 2239 | 327 | 1766 | 1444 | 813 | Bone morphogenetic protein 7 |
| CYT | IL6ST | 1239 | 1842 | 1090 | 1390 | 325 | Interleukin-6 receptor subunit beta |
| GF | HGF | 1058 | 1462 | 1443 | 1321 | 186 | Hepatocyte growth factor |
| GF | FGF4 | 1536 | 1484 | 729 | 1250 | 369 | Fibroblast growth factor 4 |
| CYT | ANG | 971 | 1427 | 1301 | 1233 | 193 | Angiogenin |
| REC | ALCAM | 956 | 1209 | 1239 | 1135 | 127 | CD166 antigen |
| CYT | FST | 690 | 1260 | 1284 | 1078 | 274 | Follistatin |
| CYT | CTSS | 808 | 1256 | 1150 | 1071 | 191 | Cathepsin S |
| GF | KDR | 1078 | 887 | 1102 | 1022 | 96 | Vascular endothelial growth factor receptor 2 |
| miRBase ID | Crt | |||||
|---|---|---|---|---|---|---|
| B1 | B2 | B3 | Mean | SD | Weight % | |
| hsa-miR-518f-3p | 9.26 | 12.46 | 13.80 | 11.84 | 1.90 | 17.75728 |
| hsa-miR-24-3p | 12.47 | 12.14 | 12.20 | 12.27 | 0.15 | 13.16838 |
| hsa-miR-193b-3p | 13.04 | 12.78 | 12.95 | 12.92 | 0.11 | 8.388070 |
| hsa-miR-222-3p | 13.26 | 13.12 | 12.81 | 13.06 | 0.19 | 7.591257 |
| hsa-miR-1274B | 13.12 | 13.18 | 13.33 | 13.21 | 0.09 | 6.863791 |
| hsa-miR-574-3p | 13.50 | 13.24 | 13.55 | 13.43 | 0.14 | 5.882132 |
| hsa-miR-191-5p | 13.43 | 13.53 | 13.56 | 13.51 | 0.06 | 5.582864 |
| hsa-miR-484 | 14.17 | 13.74 | 13.90 | 13.94 | 0.18 | 4.142033 |
| hsa-miR-320a-3p | 14.34 | 13.94 | 14.30 | 14.19 | 0.18 | 3.468564 |
| hsa-miR-197-3p | 15.20 | 14.51 | 14.89 | 14.86 | 0.28 | 2.181023 |
| hsa-miR-19b-3p | 15.44 | 15.55 | 15.68 | 15.55 | 0.10 | 1.352228 |
| hsa-miR-214-3p | 15.47 | 15.49 | 15.81 | 15.59 | 0.16 | 1.317989 |
| hsa-miR-99a-5p | 15.69 | 15.67 | 15.63 | 15.66 | 0.03 | 1.254117 |
| hsa-miR-145-5p | 15.61 | 15.59 | 15.80 | 15.67 | 0.09 | 1.250356 |
| hsa-miR-125b-5p | 15.76 | 15.67 | 15.65 | 15.69 | 0.05 | 1.226040 |
| hsa-miR-1274A | 15.80 | 15.93 | 16.14 | 15.96 | 0.14 | 1.021016 |
| hsa-miR-627-5p | 17.05 | 14.73 | 16.22 | 16.00 | 0.96 | 0.992408 |
| hsa-miR-342-3p | 16.22 | 15.80 | 16.38 | 16.13 | 0.25 | 0.904174 |
| hsa-miR-409-3p | 16.19 | 16.33 | 15.94 | 16.15 | 0.16 | 0.891932 |
| hsa-miR-21-5p | 15.80 | 16.24 | 16.46 | 16.17 | 0.28 | 0.884135 |
| hsa-miR-106a-5p | 16.02 | 16.42 | 16.33 | 16.25 | 0.17 | 0.832009 |
| hsa-miR-16-5p | 16.37 | 16.29 | 16.19 | 16.28 | 0.07 | 0.816961 |
| hsa-miR-17-5p | 16.10 | 16.36 | 16.39 | 16.28 | 0.13 | 0.815264 |
| hsa-let-7b-5p | 16.39 | 16.40 | 16.66 | 16.48 | 0.12 | 0.710056 |
| hsa-miR-29a-3p | 16.71 | 16.83 | 16.39 | 16.64 | 0.19 | 0.635518 |
| hsa-miR-30c-5p | 16.48 | 16.81 | 16.81 | 16.70 | 0.16 | 0.610052 |
| hsa-miR-221-3p | 16.79 | 16.75 | 16.86 | 16.80 | 0.05 | 0.569594 |
| hsa-miR-92a-3p | 17.12 | 16.93 | 17.10 | 17.05 | 0.08 | 0.478969 |
| hsa-miR-30b-5p | 16.93 | 17.20 | 17.35 | 17.16 | 0.17 | 0.444629 |
| hsa-miR-20a-5p | 16.92 | 17.28 | 17.43 | 17.21 | 0.21 | 0.428492 |
| hsa-miR-132-3p | 17.31 | 17.13 | 17.47 | 17.30 | 0.14 | 0.402206 |
| hsa-miR-618 | 13.41 | 14.78 | 24.32 | 17.50 | 4.85 | 0.350464 |
| hsa-miR-138-5p | 17.39 | 17.43 | 17.87 | 17.56 | 0.22 | 0.335722 |
| hsa-miR-382-5p | 18.55 | 17.53 | 17.41 | 17.83 | 0.51 | 0.279323 |
| hsa-miR-663b | 17.74 | 17.68 | 18.19 | 17.87 | 0.23 | 0.271308 |
| hsa-miR-483-5 | 18.29 | 17.46 | 17.99 | 17.91 | 0.34 | 0.264011 |
| hsa-miR-199a-3p | 18.14 | 18.30 | 17.86 | 18.10 | 0.18 | 0.231327 |
| hsa-miR-520e-3p | 15.88 | 17.10 | 21.67 | 18.22 | 2.49 | 0.213307 |
| hsa-miR-31-5p | 18.39 | 18.44 | 17.85 | 18.23 | 0.27 | 0.211883 |
| hsa-miR-28-3p | 18.39 | 18.51 | 18.18 | 18.36 | 0.14 | 0.193402 |
| hsa-miR-146a-5p | 18.47 | 18.77 | 18.02 | 18.42 | 0.31 | 0.185395 |
| hsa-miR-720 | 18.69 | 18.31 | 18.31 | 18.44 | 0.18 | 0.183435 |
| hsa-miR-193a-5p | 18.27 | 18.37 | 18.71 | 18.45 | 0.19 | 0.181957 |
| hsa-miR-34a-5p | 19.15 | 18.58 | 17.84 | 18.52 | 0.54 | 0.172740 |
| hsa-miR-376a-3p | 18.51 | 18.59 | 18.58 | 18.56 | 0.03 | 0.168094 |
| hsa-miR-186-5p | 19.01 | 18.74 | 18.40 | 18.72 | 0.25 | 0.151005 |
| CHO | SYN | HLA-DR+ | T CELL | % WEIGHT | MAIN miRNA | |
|---|---|---|---|---|---|---|
| CYTOKINES | ||||||
| TNF | X | X | 2.05 | hsa-miR-125-5p (1.23%) | ||
| IL6 | X | X | 0.19 | hsa-miR-146a-5p (0.19%) | ||
| IL1B | X | X | 0.88 | hsa-miR-21-5p (0.88%) | ||
| IL1A | X | X | 5.58 | hsa-miR-191-5p (5.58%) | ||
| CXCL12 | X | X | 0.97 | hsa-miR-221-3p (0.57%) | ||
| CCL5 | X | X | X | 1.51 | hsa-miR-214-3p (1.32%) | |
| IL11 | X | X | X | 0.61 | hsa-miR-30c-5p (0.61%) | |
| GROWTH FACTORS | ||||||
| TGFB1 | X | X | X | X | 19.24 | hsa-miR-24-3p (13.17%) |
| IGF1 | X | X | 0.87 | hsa-miR-29a-3p (0.64%) | ||
| FGF2 | X | X | 0.97 | hsa-miR-16-5p (0.82%) | ||
| BMP2 | X | X | X | 1.65 | hsa-miR-106a-5p (0.83%) | |
| VEGFA | X | X | X | 6.22 | hsa-miR-145-5p (1.25%) | |
| HGF | X | X | 1.05 | hsa-miR-16-5p (0.82%) | ||
| ANGPT2 | X | X | 2.48 | hsa-miR-145-5p (1.25%) | ||
| CTGF | X | X | X | 1.86 | hsa-miR-145-5p (1.25%) | |
| KITLG | X | X | X | 3.47 | hsa-miR-320a-3p (3.47%) | |
| TGFB2 | X | X | X | 2.13 | hsa-miR-145-5p (1.25%) | |
| INHBB | X | 0.17 | hsa-miR-34a-5p (0.17%) | |||
| IGF2 | X | X | 1.5 | hsa-miR-125b-5p (1.23%) | ||
| BDNF | X | 1.22 | hsa-miR-16-5p (0.82%) | |||
| PROTEASES | ||||||
| ADAM12 | X | X | 0.64 | hsa-miR-29a-3p (0.64%) | ||
| ADAM17 | X | X | X | 1.25 | hsa-miR-145-5p (1.25%) | |
| ADAMTS9 | X | 0.64 | hsa-miR-29a-3p (0.64%) | |||
| MMP1 | X | 8.84 | hsa-miR-222-3p (7.59%) | |||
| MMP2 | X | X | 3.26 | hsa-miR-125b-5p (1.23%) | ||
| MMP9 | X | X | 0.4 | hsa-miR-132-3p (0.40%) | ||
| MMP13 | 1.23 | hsa-miR-125b-5p (1.23%) | ||||
| MMP14 | X | X | 14.42 | hsa-miR-24-3p (13.17%) | ||
| PLAU | X | X | 8.39 | hsa-miR-193b-3p (8.39%) | ||
| PLAT | X | X | 0.88 | hsa-miR-21-5p (0.88%) | ||
| APC | X | X | 2.06 | hsa-miR-125b-5p (1.23%) | ||
| TIMP2 | X | X | 1.26 | hsa-miR-106a-5p (0.83%) | ||
| TIMP3 | X | X | 9.86 | hsa-miR-222-3p (7.59%) |
| TISSUE/CELLS | % WEIGHT | ROLE |
|---|---|---|
| CARTILAGE | ||
| Protective | ||
| hsa-miR-24-3p | 13.17 | Prevents ECM degradation, increases chondrocyte viability |
| hsa-miR-193b-3p | 8.39 | Reduces cartilage degradation |
| hsa-miR-222-3p | 7.59 | Reduces cartilage degradation |
| hsa-miR-320a-3p | 3.47 | Increases chondrocyte viability |
| hsa-miR-125b-5p | 1.23 | Prevents aggrecan loss |
| hsa-miR-17-5p | 0.82 | Induces autophagy |
| hsa-miR-221-3p | 0.57 | Prevents ECM degradation |
| hsa-miR-92a-3p | 0.48 | Increases collagen deposition |
| hsa-miR-199a-3p | 0.23 | Anti-catabolic |
| Total | 35.94 | |
| Destructive | ||
| hsa-miR-21-5p | 0.88 | Negatively regulates chondrogenesis |
| hsa-miR-16-5p | 0.82 | Cartilage degradation |
| hsa-miR-30b-5p | 0.44 | Pro-apoptotic, ECM degradation |
| hsa-miR-138-5p | 0.34 | Cartilage degradation |
| hsa-miR-483-5 | 0.26 | Chondrocyte hypertrophy, ECM degradation and cartilage angiogenesis |
| hsa-miR-146a-5p | 0.19 | Activator in early OA |
| hsa-miR-34a-5p | 0.17 | Apoptosis |
| Total | 3.10 | |
| Dual | ||
| hsa-miR-145-5p | 1.25 | Regulates chondrocyte proliferation and fibrosis |
| SYNOVIUM | ||
| Protective | ||
| hsa-miR-29a-3p | 0.64 | Anti-fibrotic effects |
| Destructive | ||
| hsa-miR-34a-5p | 0.17 | Enhances synovial inflammation |
| Dual | ||
| hsa-miR-146a-5p | 0.19 | Enhances/Suppresses synovial inflammation |
| MACROPHAGE | ||
| M1 | ||
| hsa-miR-145-5p | 1.25 | Pro-M1 |
| hsa-miR-125b-5p | 1.23 | Pro-M1 |
| Total | 2.48 | |
| M2 | ||
| hsa-miR-24-3p | 13.17 | Pro M2, blocks M1 |
| hsa-miR-222-3p | 7.59 | Pro M2 |
| hsa-let-7b-5p | 0.71 | Pro M2 |
| hsa-miR-146a-5p | 0.19 | Pro M2, blocks M1 |
| hsa-miR-34a-5p | 0.17 | Pro M2 |
| Total | 21.83 | |
| T CELL | ||
| Pro-Activation | ||
| hsa-miR-19b-3p | 1.35 | Reduces PTEN repressor |
| hsa-miR-214-3p | 1.32 | Reduces PTEN repressor |
| hsa-miR-21-5p | 0.88 | Reduces PTEN repressor |
| hsa-miR-106a-5p | 0.83 | Represses IL10 |
| hsa-miR-17-5p | 0.82 | Reduces PTEN repressor and promotes IFNγ |
| hsa-let-7b-5p | 0.71 | Represses IL10 |
| hsa-miR-221-3p | 0.57 | Downregulates PIK3R1 |
| hsa-miR-132-3p | 0.40 | Downregulates PIK3R1 |
| Total | 6.88 | |
| Anti-activation | ||
| hsa-miR-24-3p | 13.17 | Represses IFNγ in activated CD4+ and CD8+ |
| hsa-miR-125b-5p | 1.23 | Maintains T cell naïve state |
| hsa-miR-342-3p | 0.90 | Downregulated upon activation |
| hsa-miR-146a-5p | 0.19 | Represses activation markers |
| Total | 15.48 | |
| Dual | ||
| hsa-miR-31-5p | 0.21 | Upregulates IL2, downregulated with activation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ragni, E.; Perucca Orfei, C.; Valli, F.; Zagra, L.; de Girolamo, L. Molecular Characterization of Secreted Factors and Extracellular Vesicles-Embedded miRNAs from Bone Marrow-Derived Mesenchymal Stromal Cells in Presence of Synovial Fluid from Osteoarthritis Patients. Biology 2022, 11, 1632. https://doi.org/10.3390/biology11111632
Ragni E, Perucca Orfei C, Valli F, Zagra L, de Girolamo L. Molecular Characterization of Secreted Factors and Extracellular Vesicles-Embedded miRNAs from Bone Marrow-Derived Mesenchymal Stromal Cells in Presence of Synovial Fluid from Osteoarthritis Patients. Biology. 2022; 11(11):1632. https://doi.org/10.3390/biology11111632
Chicago/Turabian StyleRagni, Enrico, Carlotta Perucca Orfei, Federico Valli, Luigi Zagra, and Laura de Girolamo. 2022. "Molecular Characterization of Secreted Factors and Extracellular Vesicles-Embedded miRNAs from Bone Marrow-Derived Mesenchymal Stromal Cells in Presence of Synovial Fluid from Osteoarthritis Patients" Biology 11, no. 11: 1632. https://doi.org/10.3390/biology11111632
APA StyleRagni, E., Perucca Orfei, C., Valli, F., Zagra, L., & de Girolamo, L. (2022). Molecular Characterization of Secreted Factors and Extracellular Vesicles-Embedded miRNAs from Bone Marrow-Derived Mesenchymal Stromal Cells in Presence of Synovial Fluid from Osteoarthritis Patients. Biology, 11(11), 1632. https://doi.org/10.3390/biology11111632