
Projecting the Potential Distribution Areas of Ixodes scapularis (Acari: Ixodidae) Driven by Climate Change


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Software and Geographic Data
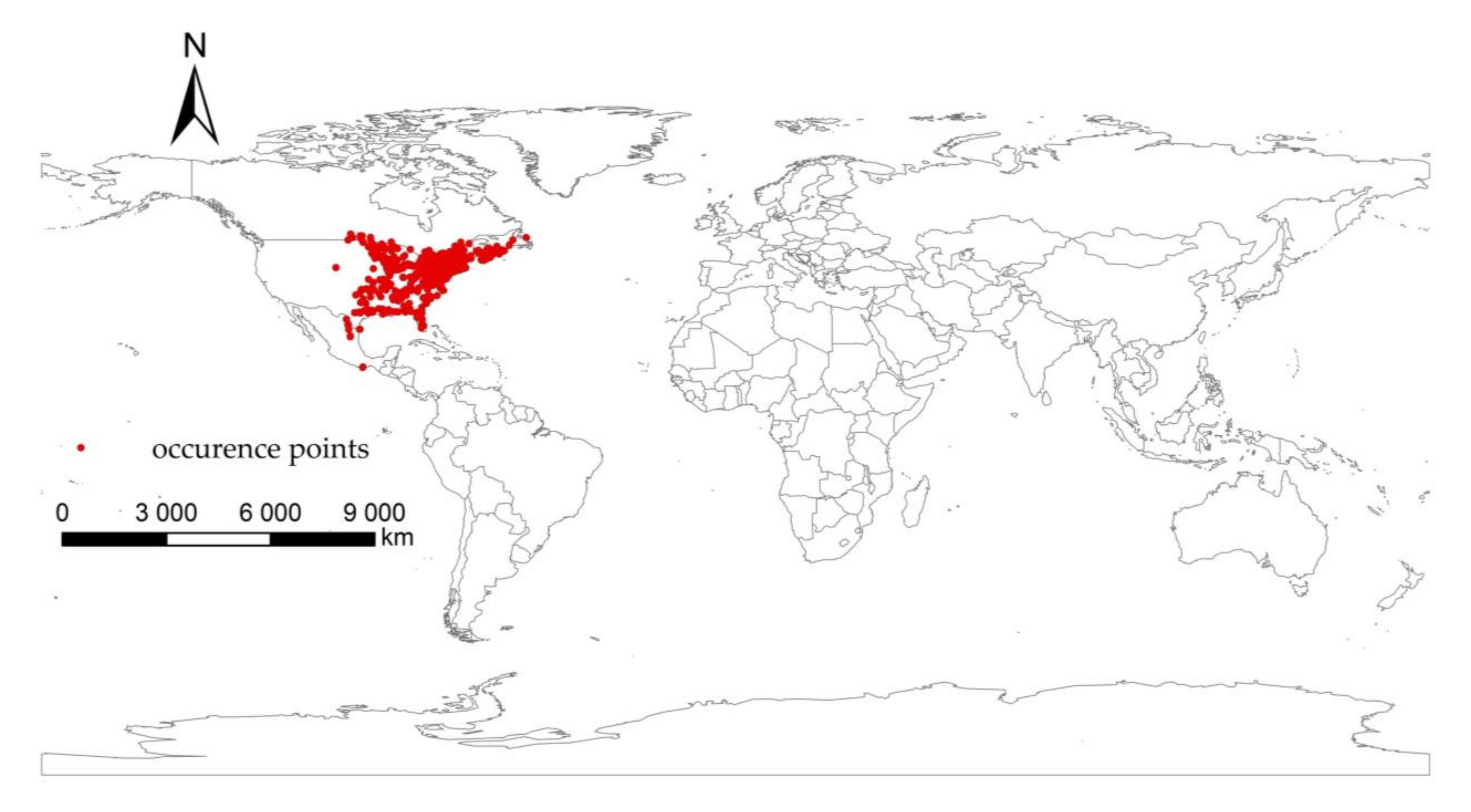
2.2. Distribution Data Collection and Processing of I. scapularis
2.3. Environmental Data
2.4. Selection of Model Parameters
2.5. Project the Potential Suitable Area
3. Results
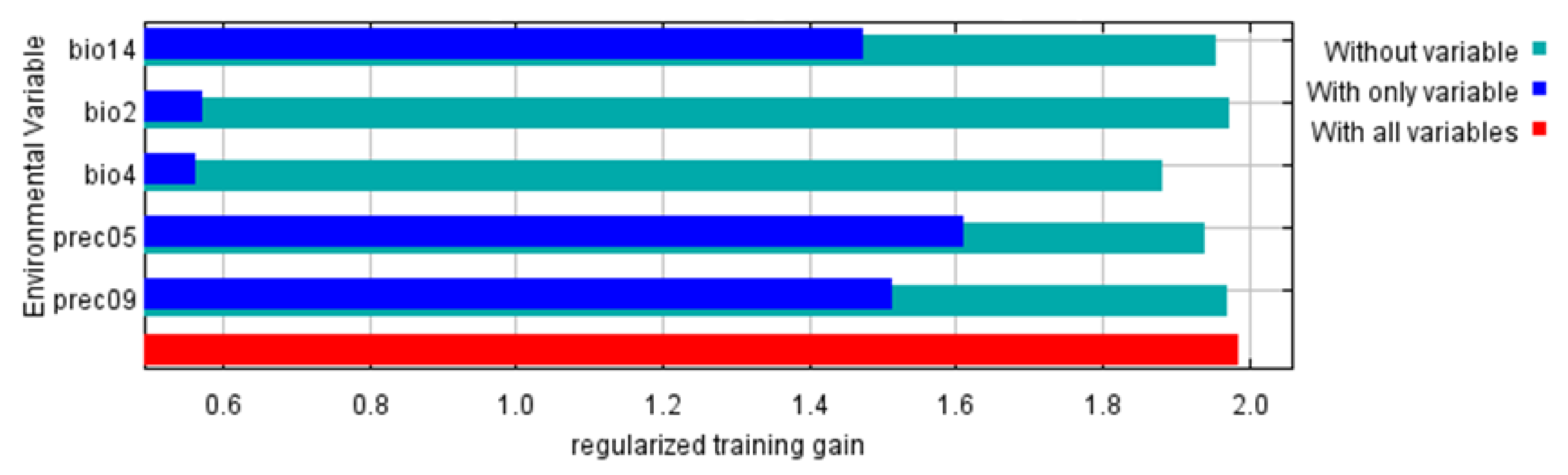
3.1. The Main Contributing Environmental Variables
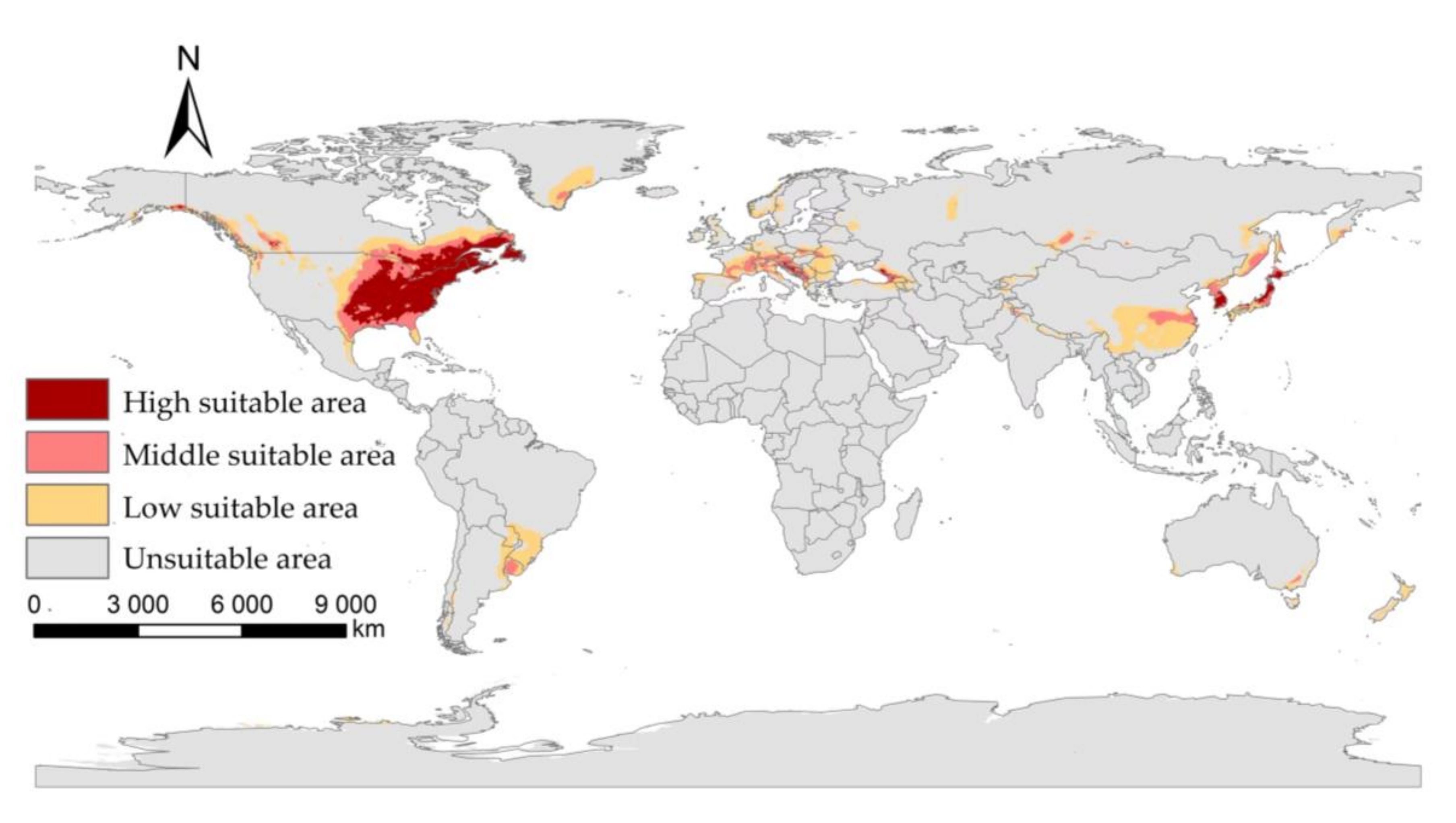
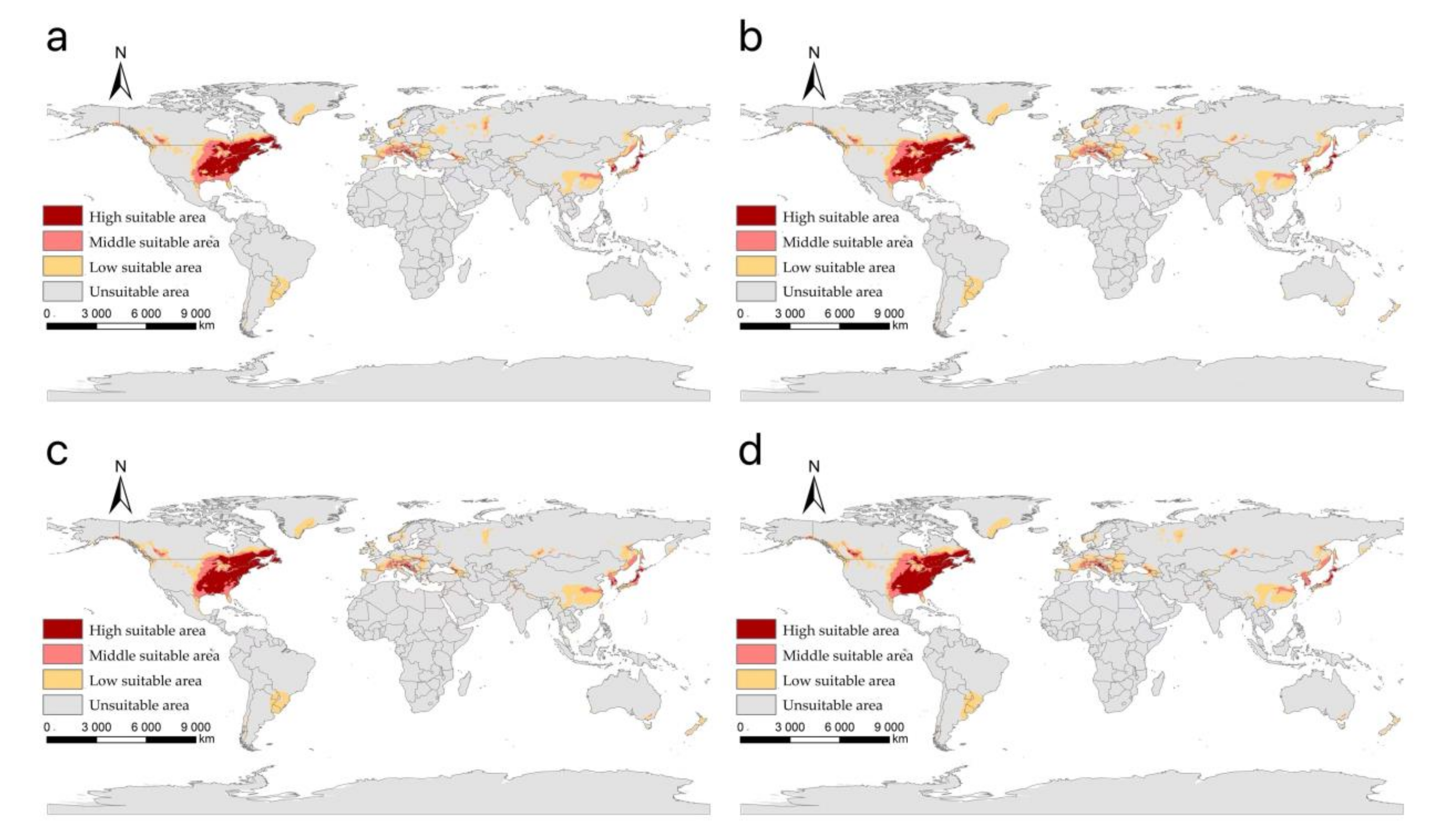
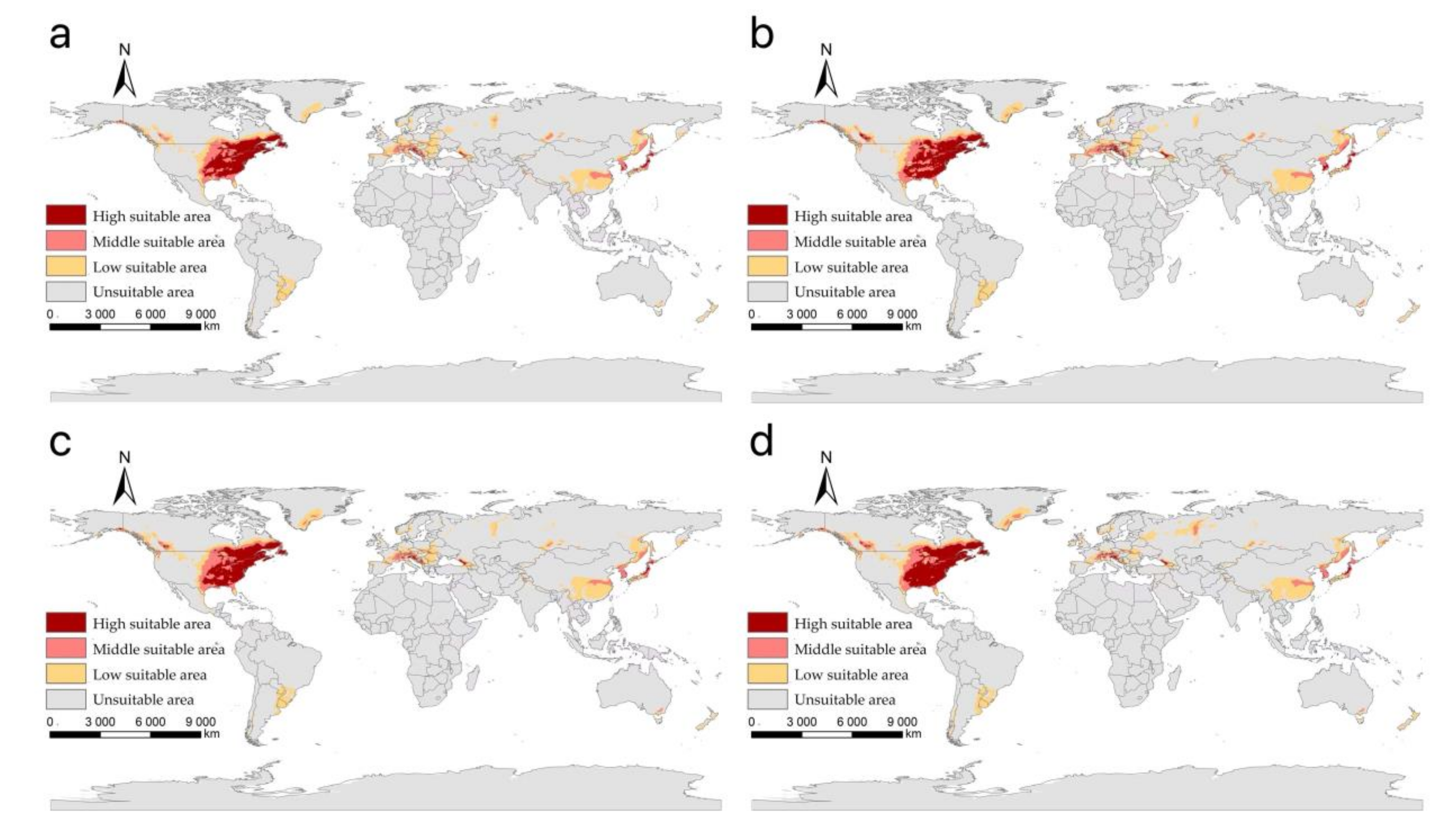
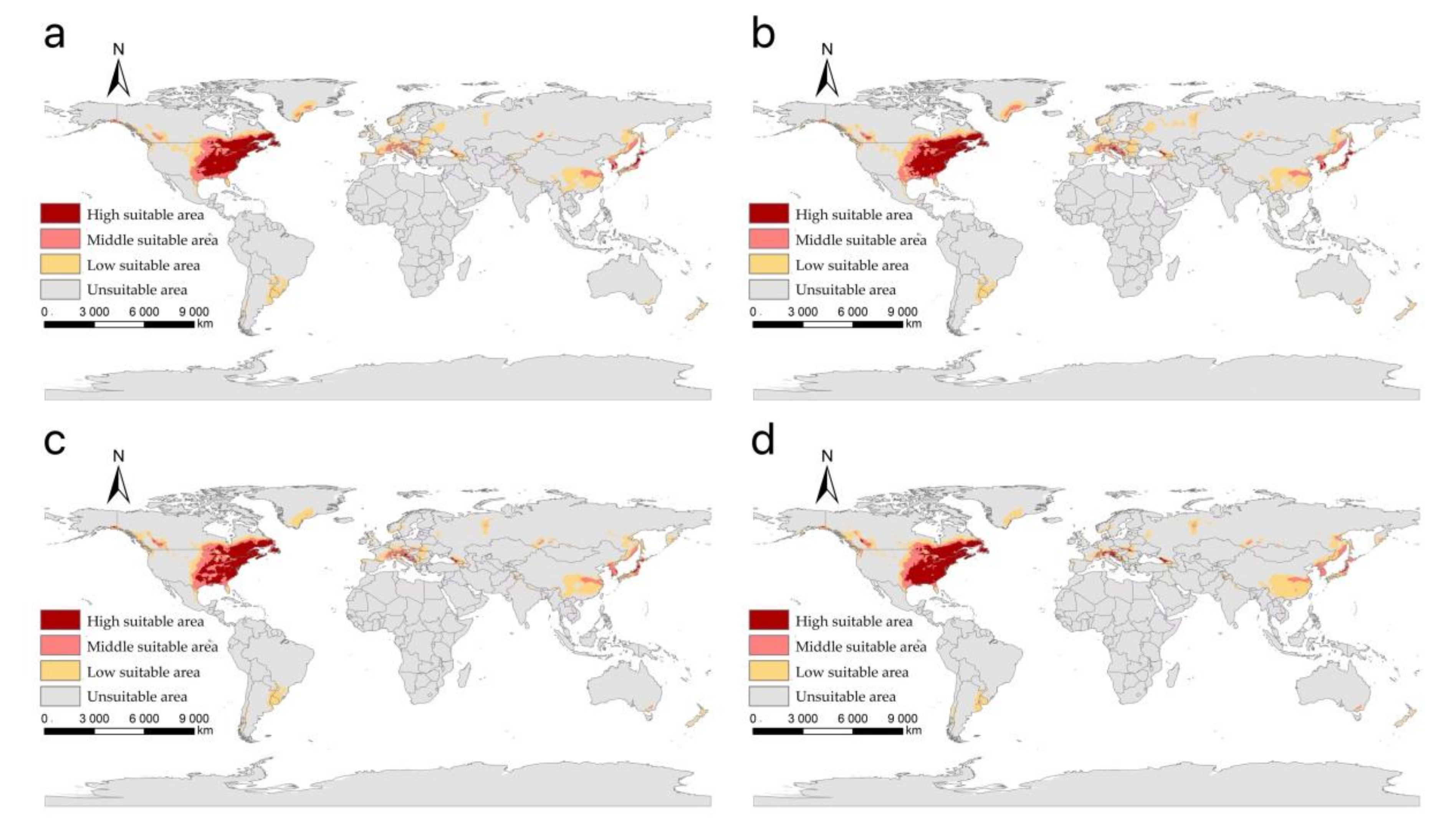
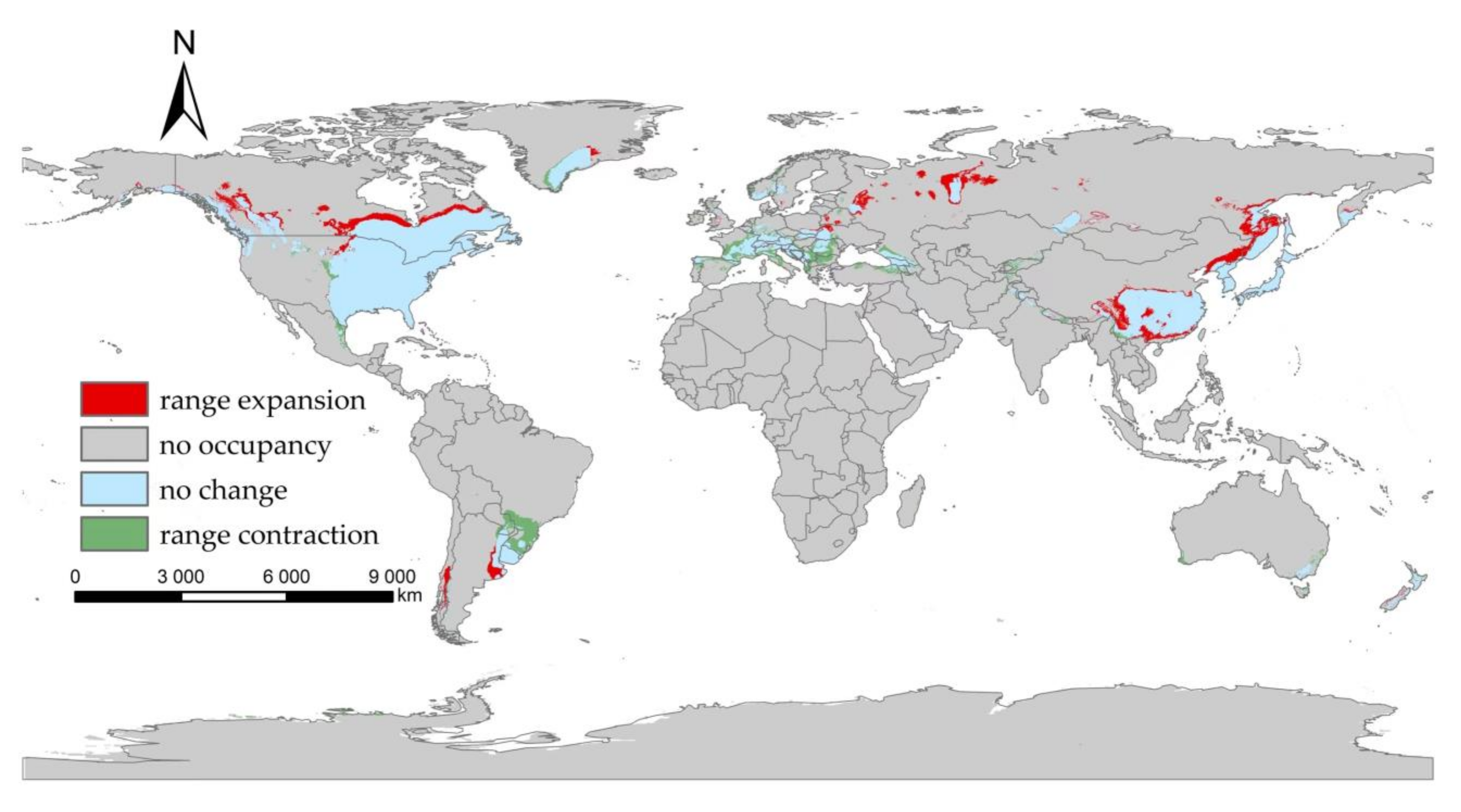
3.2. The Suitable Areas of I. scapularis under Near Current Climatic Condition
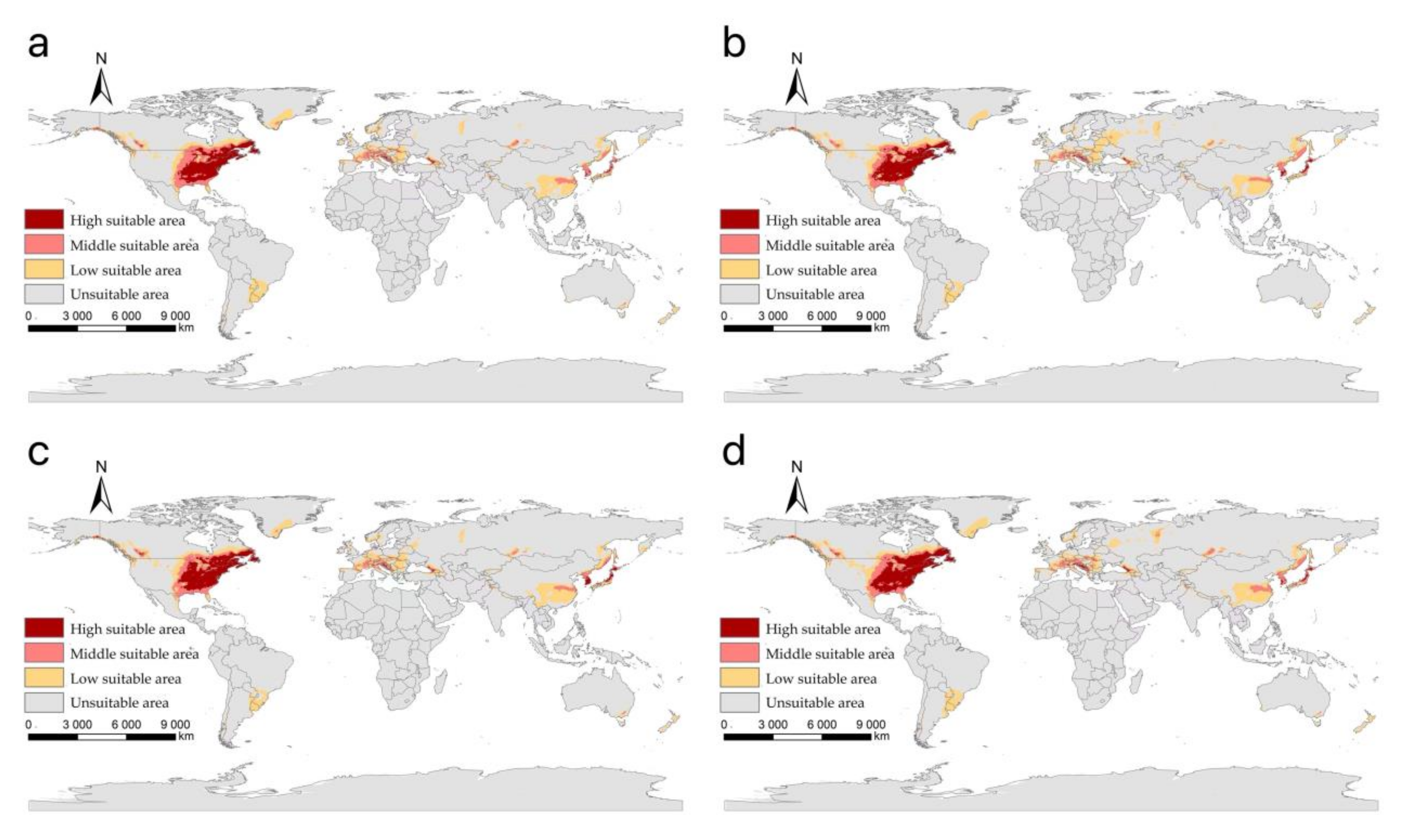
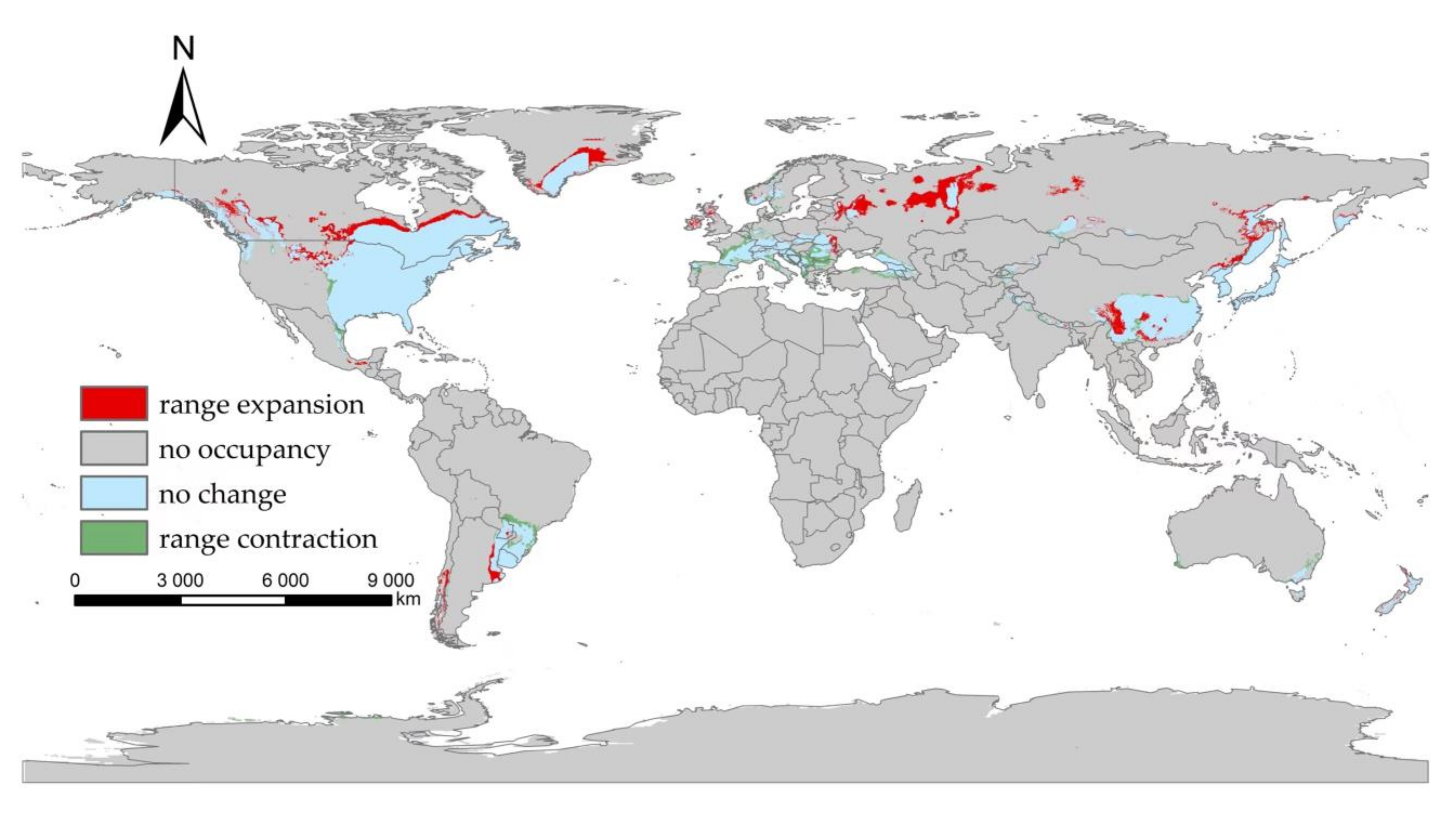
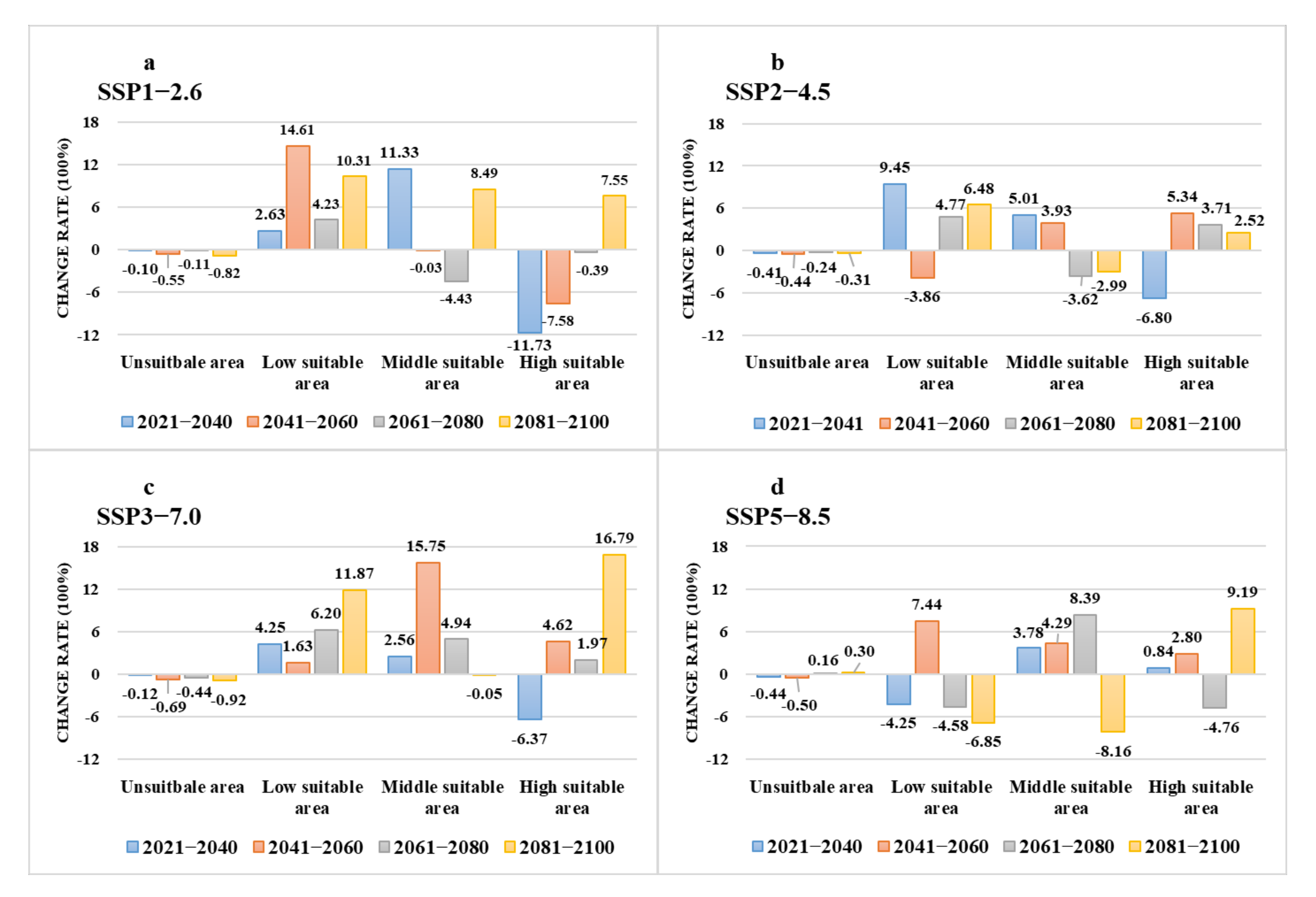
3.3. The Suitable Areas of I. scapularis under Future Climate Change Scenarios
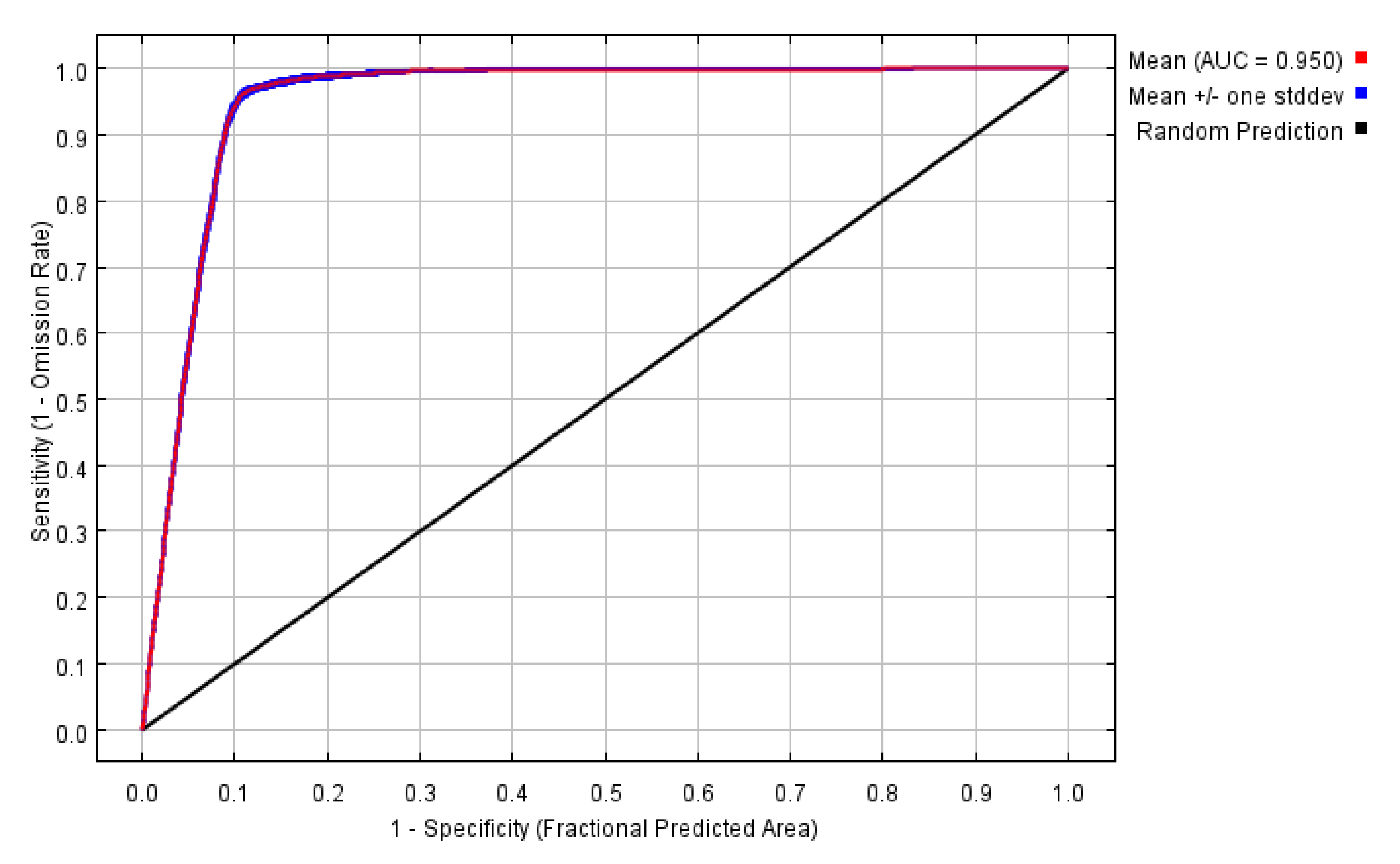
3.4. The Accuracy of the MaxEnt Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ogden, N.H.; Maarouf, A.; Barker, I.K.; Bigras-Poulin, M.; Lindsay, L.R.; Morshed, M.G.; O’Callaghan, C.J.; Ramay, F.; Waltner-Toews, D.; Charron, D.F. Climate change and the potential for range expansion of the Lyme disease vector Ixodes scapularis in Canada. Int. J. Parasitol. 2006, 36, 63–70. [Google Scholar] [CrossRef]
- Eisen, R.J.; Eisen, L. The Blacklegged Tick, Ixodes scapularis: An Increasing Public Health Concern. Trends Parasitol. 2018, 34, 295–309. [Google Scholar] [CrossRef]
- Sanchez-Vicente, S.; Tagliafierro, T.; Coleman, J.L.; Benach, J.L.; Tokarz, R. Polymicrobial Nature of Tick-Borne Diseases. mBio 2019, 10, e02055-19. [Google Scholar] [CrossRef] [Green Version]
- Ogden, N.H.; Radojevic, M.; Wu, X.; Duvvuri, V.R.; Leighton, P.A.; Wu, J. Estimated effects of projected climate change on the basic reproductive number of the Lyme disease vector Ixodes scapularis. Environ. Health Perspect. 2014, 122, 631–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacon, R.M.; Kugeler, K.J.; Mead, P.S. Surveillance for Lyme disease—United States, 1992–2006. MMWR Surveill. Summ. 2008, 57, 1–9. [Google Scholar] [PubMed]
- Semenza, J.C.; Suk, J.E. Vector-borne diseases and climate change: A European perspective. FEMS Microbiol. Lett. 2018, 365, fnx244. [Google Scholar] [CrossRef]
- Couper, L.I.; MacDonald, A.J.; Mordecai, E.A. Impact of prior and projected climate change on US Lyme disease incidence. Glob. Chang. Biol. 2021, 27, 738–754. [Google Scholar] [CrossRef] [PubMed]
- Levy, S. Ticking time bomb? Climate change and Ixodes scapularis. Environ. Health Perspect. 2014, 122, A168. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, B.; Kriegler, E.; Ebi, K.L.; Kemp-Benedict, E.; Riahi, K.; Rothman, D.S.; Ruijven, B.V.; Vuuren, D.V.; Birkmann, J.; Kok, K. The roads ahead: Narratives for shared socioeconomic pathways describing world futures in the 21st century. Glob. Environ. Change 2017, 42, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Duan, R.Y.; Kong, X.Q.; Huang, M.Y.; Varela, S.; Ji, X. The potential effects of climate change on amphibian distribution, range fragmentation and turnover in China. PeerJ 2016, 4, e2185. [Google Scholar] [CrossRef] [Green Version]
- Alkishe, A.A.; Peterson, A.T.; Samy, A.M. Climate change influences on the potential geographic distribution of the disease vector tick Ixodes ricinus. PLoS ONE 2017, 12, e0189092. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Lun, X.; Li, C.; Zhou, R.; Zhao, Z.; Wang, J.; Zhang, Q.; Liu, Q. Predicting the Potential Global Distribution of (Acari: Ixodidae) under Near Current and Future Climatic Conditions, Using the Maximum Entropy Model. Biology 2021, 10, 1057. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Gao, Y.; Chang, N.; Gao, T.; Ma, D.; Li, C.; Liu, Q. Projecting the Potential Distribution of (Diptera: Glossinidae) under Climate Change Using the MaxEnt Model. Biology 2021, 10, 1150. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Shi, J. The Potential Global Distribution of Sirex juvencus (Hymenoptera: Siricidae) under Near Current and Future Climatic Conditions as Predicted by the Maximum Entropy Model. Insects 2021, 12, 222. [Google Scholar] [CrossRef] [PubMed]
- Cauwer, V.D.; Muys, B.; Revermann, R.; Trabucco, A. Potential, realised, future distribution and environmental suitability for Pterocarpus angolensis DC in southern Africa. For. Ecol. Manag. 2014, 315, 211–226. [Google Scholar] [CrossRef]
- Gastón, A.; Garcia-Vinas, J.I.; Bravo-Fernandez, A.J.; López-Leiva, C.; Oliet, J.A.; Roig, S.; Serrada, R. Species distribution models applied to plant species selection in forest restoration: Are model predictions comparable to expert opinion? New Forest 2014, 45, 641–653. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, B.; Wan, F.; Xiao, Q.; Dai, L. Application of ROC curve analysis in evaluating the performance of alien species’ potential distribution models. Biodivers. Sci. 2007, 15, 365–372. [Google Scholar]
- Xiang, L.; Fucheng, Z.; Shuai, H.; Yuqing, W.; Lin, M. Research Progress Analysison the Comprehensive Application of MaxEnt Model. J. Green Sci. Technol. 2020, 18, 14–17. [Google Scholar]
- Soucy, J.R.; Slatculescu, A.M.; Nyiraneza, C.; Ogden, N.H.; Leighton, P.A.; Kerr, J.T.; Kulkarni, M.A. High-Resolution Ecological Niche Modeling of Ixodes scapularis Ticks Based on Passive Surveillance Data at the Northern Frontier of Lyme Disease Emergence in North America. Vector Borne Zoonotic Dis. 2018, 18, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Kearney, M.; Phillips, S. The art of modelling range-shifting species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Shcheglovitova, M.; Anderson, R.P. Estimating optimal complexity for ecological niche models: A jackknife approach for species with small sample sizes. Ecol. Model. 2013, 269, 9–17. [Google Scholar] [CrossRef]
- Morales, N.S.; Fernández, I.C.; Baca-González, V. MaxEnt’s parameter configuration and small samples: Are we paying attention to recommendations? A systematic review. PeerJ 2017, 5, e3093. [Google Scholar] [CrossRef]
- Dan, L.W.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar]
- Raghavan, R.K.; Peterson, A.T.; Cobos, M.E.; Ganta, R.; Foley, D. Current and Future Distribution of the Lone Star Tick, Amblyomma americanum (L.) (Acari: Ixodidae) in North America. PLoS ONE 2019, 14, e0209082. [Google Scholar] [CrossRef]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef] [Green Version]
- Franklin, J. Ecological understanding of species distributions. In Mapping Species Distributions (Spatial Inference and Prediction); Cambridge University Press: Cambridge, UK, 2010; pp. 34–52. [Google Scholar]
- Franklin, J. Mapping Species Distributions (Spatial Inference and Prediction) || Ecological Understanding of Species Distributions; Cambridge University Press: Cambridge, UK, 2010; pp. 34–52. [Google Scholar]
- Ostfeld, R.S.; Brunner, J.L. Climate change and Ixodes tick-borne diseases of humans. Philos. Trans. R Soc. Lond. B Biol. Sci. 2015, 370, 20140051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, K.A.; Ginsberg, H.S.; Gonzalez, L.; Mather, T.N. Relative humidity and activity patterns of Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 2014, 51, 769–776. [Google Scholar] [CrossRef]
- Hahn, M.B.; Jarnevich, C.S.; Monaghan, A.J.; Eisen, R.J. Modeling the Geographic Distribution of Ixodes scapularis and Ixodes pacificus (Acari: Ixodidae) in the Contiguous United States. J. Med. Entomol. 2016, 53, 1176–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, T.L.; Bjork, J.K.H.; Neitzel, D.F.; Dorr, F.M.; Schiffman, E.K.; Eisen, R.J. Habitat Suitability Model for the Distribution of Ixodes scapularis (Acari: Ixodidae) in Minnesota. J. Med. Entomol. 2016, 53, 598–606. [Google Scholar] [CrossRef] [Green Version]
- Mcpherson, M.Y.; Garcíagarcía, A.; Cuestavalero, F.J.; Beltrami, H.; Hansenketchum, P.; Macdougall, D.; Ogden, N.H. Expansion of the Lyme Disease Vector Ixodes scapularis in Canada inferred from CMIP5 Climate Projections. In Proceedings of the EGU General Assembly Conference, Vienna, Austria, 23–28 April 2017. [Google Scholar]
- Hoegh-Guldberg, O.; Jacob, D.; Taylor, M.; Guillén Bolaños, T.; Bindi, M.; Brown, S.; Camilloni, I.A.; Diedhiou, A.; Djalante, R.; Ebi, K.; et al. The human imperative of stabilizing global climate change at 1.5 °C. Science 2019, 365, eaaw6974. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Liu, C.; Xie, F.; Lu, J.; Chen, B. Precipitation Characteristic Changes due to Global Warming in a High-Resolution (16-km) ECMWF Simulation. Q. J. R. Meteorol. Soc. 2018, 145, 303–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada-PeñA, A.; José, D. The ecology of ticks and epidemiology of tick-borne viral diseases. Antiviral Res. 2014, 108, 104–128. [Google Scholar] [CrossRef] [PubMed]
- Clow, K.M.; Ogden, N.H.; Lindsay, L.R.; Michel, P.; Pearl, D.L.; Jardine, C.M. The influence of abiotic and biotic factors on the invasion of Ixodes scapularis in Ontario, Canada. Ticks Tick Borne Dis. 2017, 8, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Lubelczyk, C.B.; Elias, S.P.; Rand, P.W.; Holman, M.S.; Lacombe, E.H.; Smith, R.P. Habitat Associations of Ixodes scapularis (Acari: Ixodidae) in Maine. Environ. Entomol. 2004, 33, 900–906. [Google Scholar] [CrossRef]
- Guoping, Z. Spatial Distribution of Ticks and Their Hazard Prediction in China. Ph.D. Thesis, Academy of Military Sciences, Beijing, China, 2018. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Description | Unit | Contribution (%) |
|---|---|---|---|
| Prec05 | Precipitation in May | mm | 39.6 |
| Prec09 | Precipitation in September | mm | 22.0 |
| Bio14 | Precipitation of the driest month | mm | 19.6 |
| Bio4 | Temperature seasonality (standard deviation × 100) | \ | 17.0 |
| Bio2 | Mean diurnal range (mean of monthly (max temp—min temp)) | °C | 1.80 |
| Climate Scenario | Period | Low Suitable Area | Middle Suitable Area | High Suitable Area | Total Area | Area Change | Area Change Ratio (%) |
|---|---|---|---|---|---|---|---|
| current | 1970–2000 | 6.63 | 2.64 | 2.83 | 12.10 | 0.00 | |
| SSP1-2.6 | 2021–2040 | 6.80 | 2.94 | 2.50 | 12.24 | 0.14 | 1.17 |
| 2041–2060 | 7.59 | 2.64 | 2.62 | 12.86 | 0.75 | 6.21 | |
| 2061–2080 | 6.91 | 2.53 | 2.82 | 12.26 | 0.15 | 1.25 | |
| 2081–2100 | 7.31 | 2.87 | 3.05 | 13.22 | 1.12 | 9.26 | |
| SSP2-4.5 | 2021–2040 | 7.25 | 2.78 | 2.64 | 12.67 | 0.57 | 4.67 |
| 2041–2060 | 6.97 | 2.75 | 2.98 | 12.70 | 0.60 | 4.97 | |
| 2061–2080 | 6.94 | 2.55 | 2.94 | 12.43 | 0.33 | 2.69 | |
| 2081–2100 | 7.06 | 2.56 | 2.90 | 12.53 | 0.42 | 3.48 | |
| SSP3-7.0 | 2021–2040 | 6.91 | 2.71 | 2.65 | 12.27 | 0.17 | 1.39 |
| 2041–2060 | 7.02 | 3.06 | 2.96 | 13.04 | 0.94 | 7.77 | |
| 2061–2080 | 7.04 | 2.77 | 2.89 | 12.70 | 0.60 | 4.93 | |
| 2081–2100 | 7.41 | 2.64 | 3.31 | 13.36 | 1.26 | 10.41 | |
| SSP5-8.5 | 2021–2040 | 7.10 | 2.74 | 2.86 | 12.70 | 0.60 | 4.92 |
| 2041–2060 | 7.12 | 2.76 | 2.91 | 12.79 | 0.69 | 5.66 | |
| 2061–2080 | 6.32 | 2.87 | 2.70 | 11.89 | −0.22 | −1.79 | |
| 2081–2100 | 6.17 | 2.43 | 3.09 | 11.69 | −0.41 | −3.38 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Ma, D.; Li, C.; Zhou, R.; Wang, J.; Liu, Q. Projecting the Potential Distribution Areas of Ixodes scapularis (Acari: Ixodidae) Driven by Climate Change. Biology 2022, 11, 107. https://doi.org/10.3390/biology11010107
Zhang L, Ma D, Li C, Zhou R, Wang J, Liu Q. Projecting the Potential Distribution Areas of Ixodes scapularis (Acari: Ixodidae) Driven by Climate Change. Biology. 2022; 11(1):107. https://doi.org/10.3390/biology11010107
Chicago/Turabian StyleZhang, Lu, Delong Ma, Chao Li, Ruobing Zhou, Jun Wang, and Qiyong Liu. 2022. "Projecting the Potential Distribution Areas of Ixodes scapularis (Acari: Ixodidae) Driven by Climate Change" Biology 11, no. 1: 107. https://doi.org/10.3390/biology11010107
APA StyleZhang, L., Ma, D., Li, C., Zhou, R., Wang, J., & Liu, Q. (2022). Projecting the Potential Distribution Areas of Ixodes scapularis (Acari: Ixodidae) Driven by Climate Change. Biology, 11(1), 107. https://doi.org/10.3390/biology11010107