
The Ultrastructural Analysis of Human Colorectal Cancer Stem Cell-Derived Spheroids and Their Mouse Xenograft Shows That the Same Cells Types Have Different Ratios

 , , ,
, , , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. CSC Isolation and Culture
2.2. Animal Procedures
2.3. Scanning Electron Microscopy Protocol for Spheroids
2.4. Flow Cytometry
2.5. Lentiviral Infection
2.6. Transmission Electron Microscopy Protocol for Spheroids and Xenograft
2.7. Evaluation of Spheroids Shape and Size Parameters on SEM Images
2.8. Ultrastructural Characterization of Spheroids and Xenograft Cell Population by Transmission Electron Microscopy
2.9. Statistical Analysis
3. Results
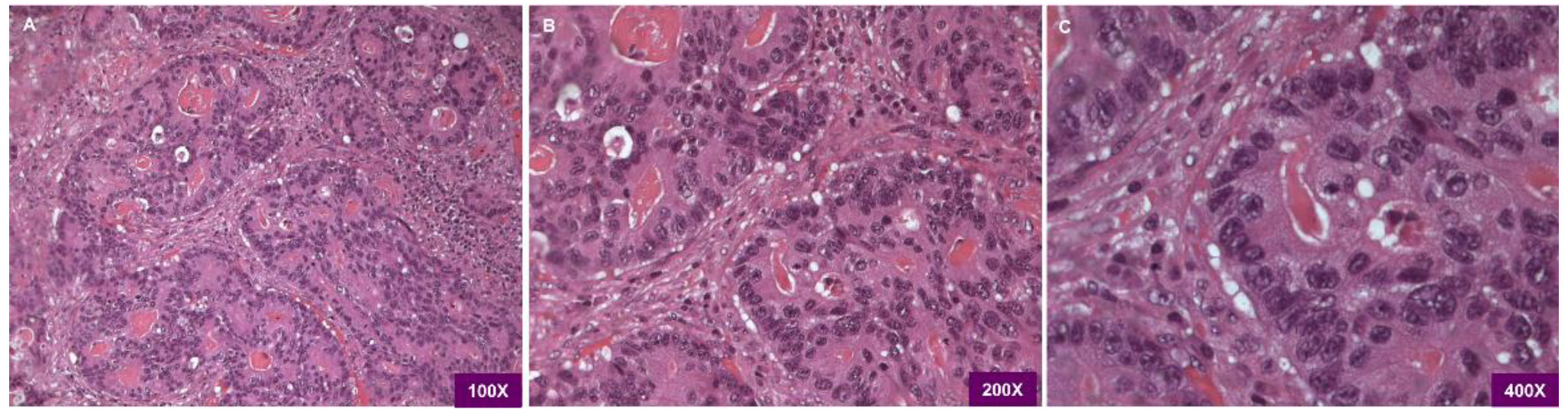
3.1. Histological Examination of the Patient Colorectal Cancer
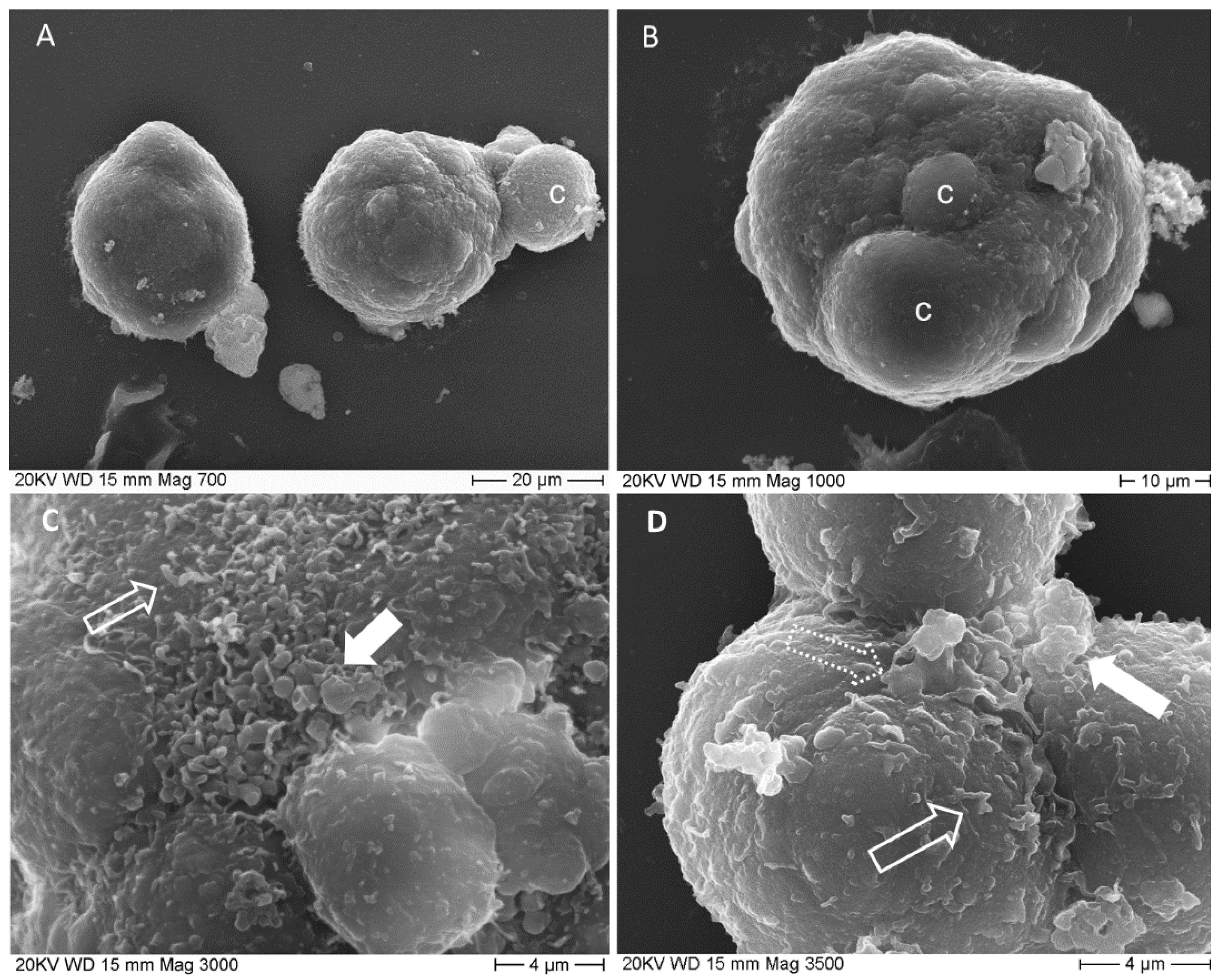
3.2. Evaluation of Patient Colorectal Cancer-Derived Spheroids Morphology by SEM
3.3. Evaluation of Patient Colorectal Cancer-Derived Spheroids by Light Microscopy
3.4. Evaluation by Light Microscopy of the Tumor Resulted from Colorectal Cancer-Derived Spheroids Xenograft in Mouse
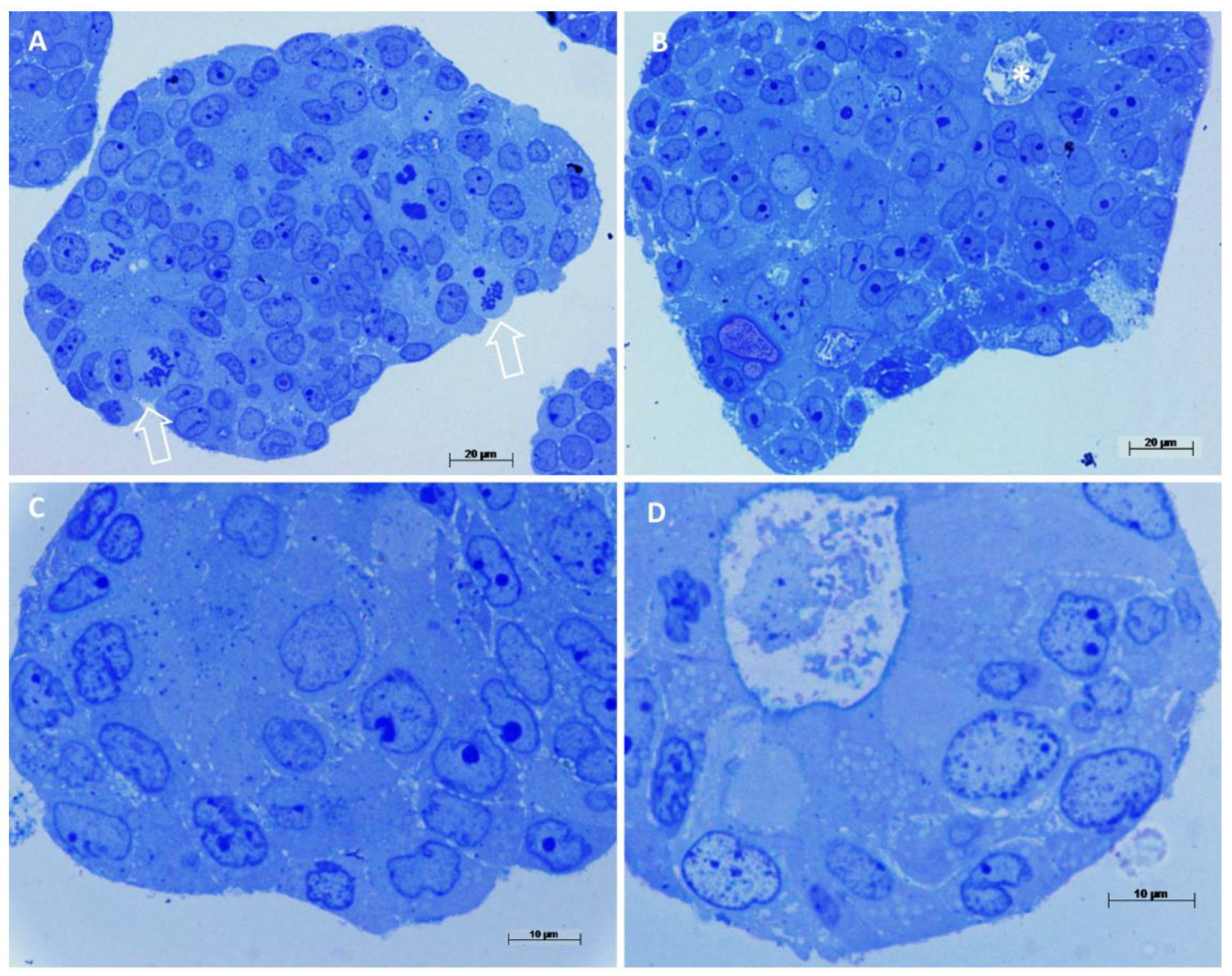
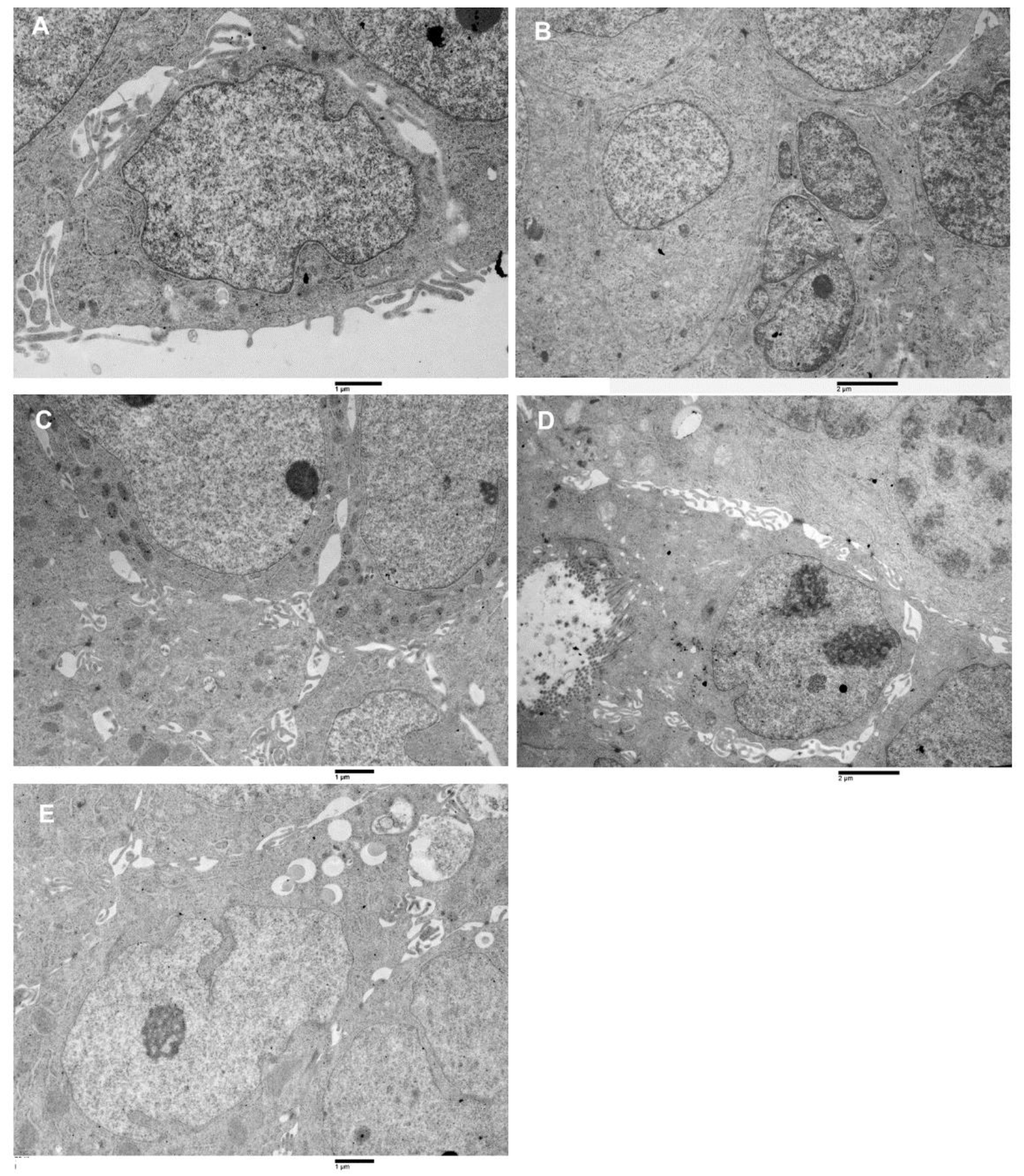
3.5. Ultrastructural Characterization of Patient Colorectal Cancer-Derived Spheroids’ Cell Population by Transmission Electron Microscopy
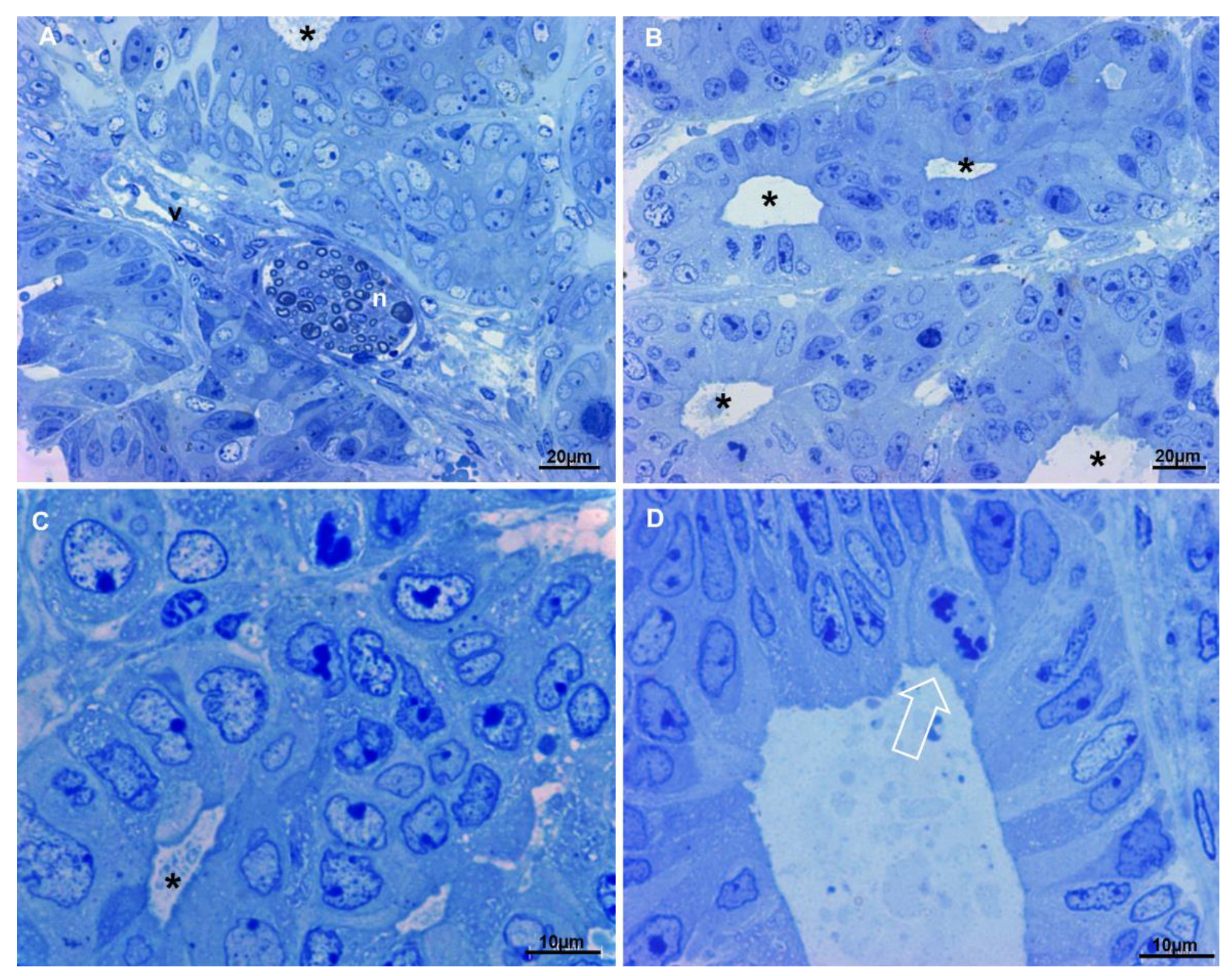
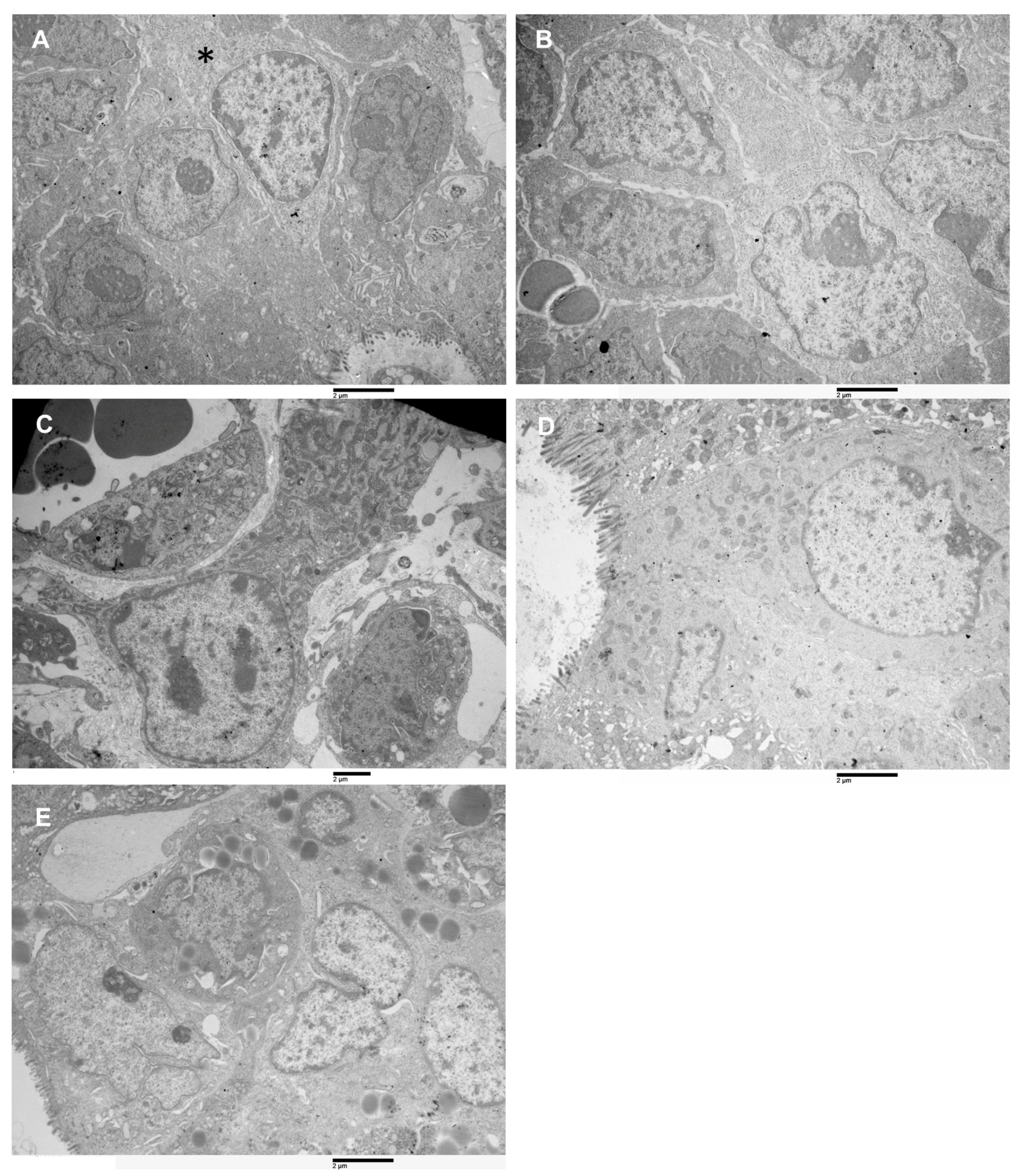
3.6. Ultrastructural Characterization of Xenograft Cell Population by Transmission Electron Microscopy
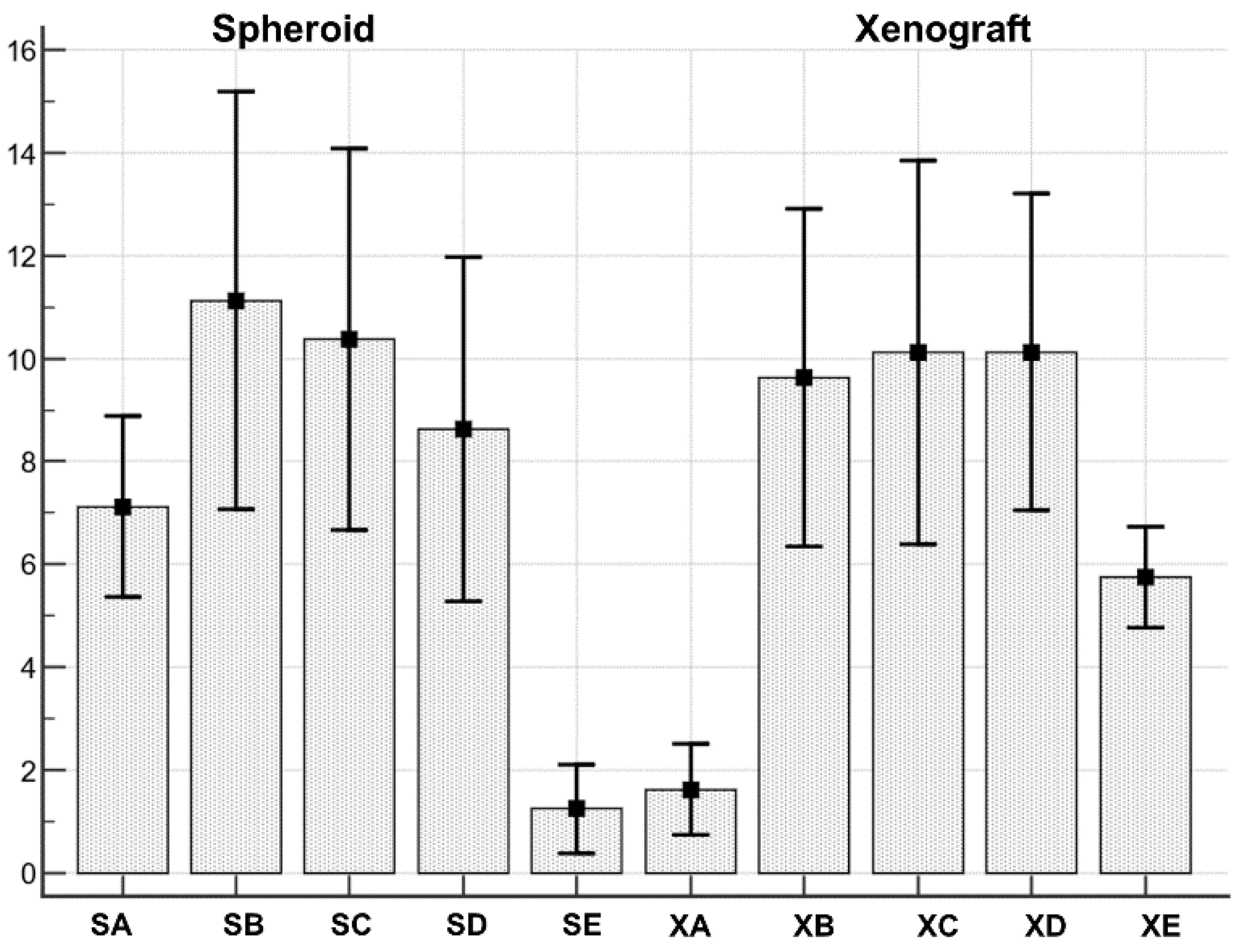
3.7. Data Statistical Analysis
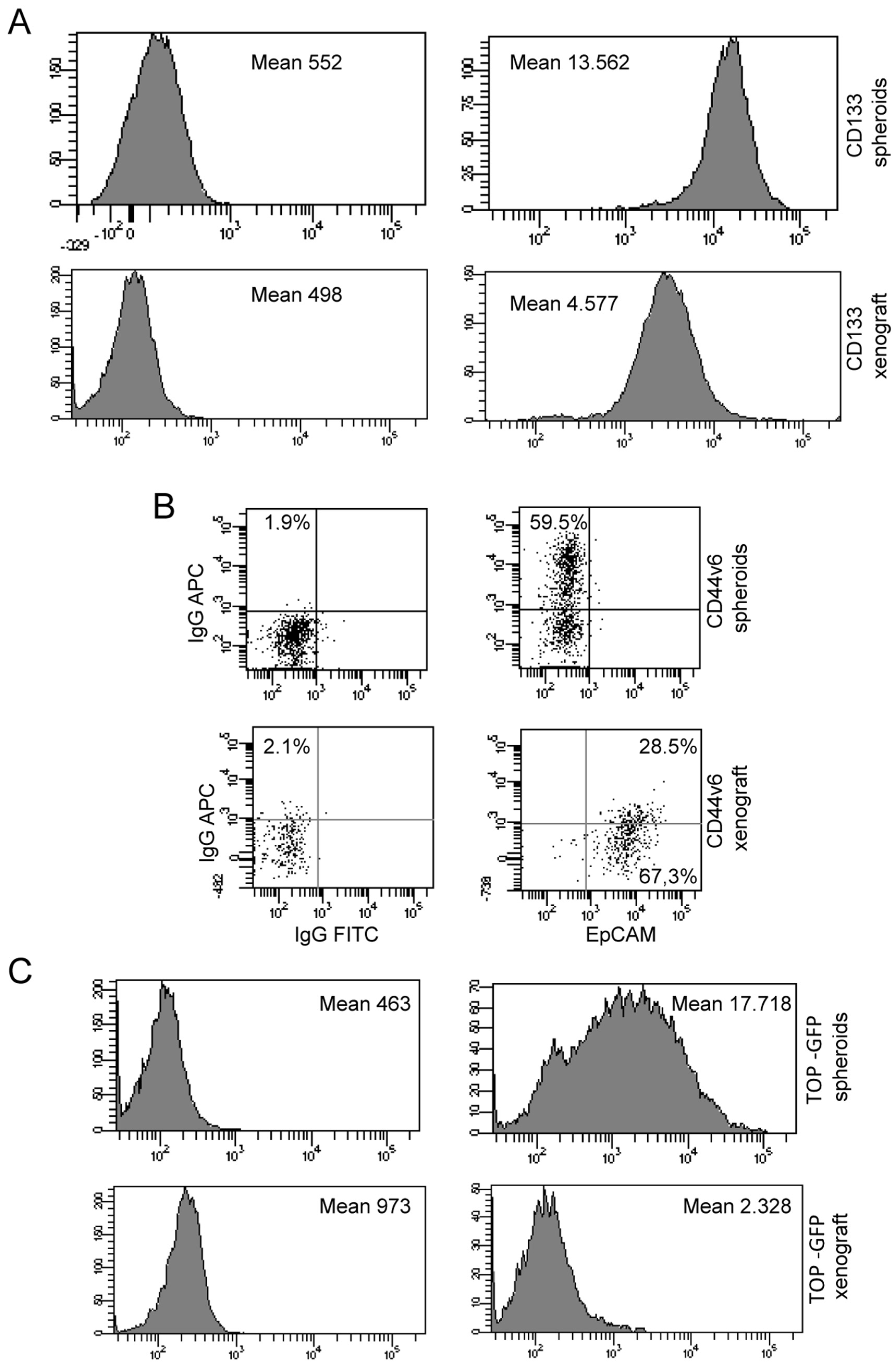
3.8. Phenotypic Analysis by Flow Cytometer of Spheroids and Xenograft Cells Reveal a Decrease in Stem Cell Number and Activity in Ex Vivo Sample
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Offit, K. Personalized medicine: New genomics, old lessons. Hum. Genet. 2011, 130, 3–14. [Google Scholar] [CrossRef]
- Mingaleeva, R.N.; Solovieva, V.V.; Blatt, N.L.; Rizvanov, A.A. Application of cell and tissue cultures for potential anti-cancer/oncology drugs screening in vitro. Cell. Transplant. Tissue Eng. 2013, 8, 20–28. [Google Scholar]
- Kitaeva, K.V.; Prudnikov, T.S.; Gomzikova, M.O.; Kletukhina, S.K.; James, V.; Rizvanov, A.A.; Solovyeva, V.V. Analysis of the interaction and proliferative activity of adenocarcinoma, peripheral blood mononuclear and mesenchymal stromal cells after co-cultivation in vitro. BioNanoScience 2019, 9, 502–509. [Google Scholar] [CrossRef]
- Ham, S.L.; Joshi, R.; Thakuri, P.S.; Tavana, H. Liquid-based three-dimensional tumor models for cancer research and drug discovery. Exp. Biol. Med. 2016, 241, 939–954. [Google Scholar] [CrossRef]
- Xu, J.; Meng, Q.; Li, X.; Yang, H.; Xu, J.; Gao, N.; Sun, H.; Wu, S.; Familiari, G.; Relucenti, M.; et al. Long Noncoding RNA MIR17HG Promotes Colorectal Cancer Progression via miR-17-5p. Cancer Res. 2019, 79, 4882–4895. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Su, J.; Fu, X.; Zheng, L.; Yin, Z. Microfluidic device for primary tumor spheroid isolation. Exp. Hematol. Oncol. 2017, 6, 22. [Google Scholar] [CrossRef] [PubMed]
- Horvath, P.; Aulner, N.; Bickle, M.; Davies, A.M.; Nery, E.D.; Ebner, D.; Montoya, M.C.; Östling, P.; Pietiäinen, V.; Price, L.S.; et al. Screening out irrelevant cell-based models of disease.Nature reviews. Drug Discov. 2016, 15, 751–769. [Google Scholar] [CrossRef] [PubMed]
- Capp, J.P. Cancer stem cells: From historical roots to a new perspective. J. Oncol. 2019, 2019. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.L.; Zeuner, A.; Policicchio, E.; Russo, G.; Bruselles, A.; Signore, M.; Vitale, S.; De Luca, G.; Pilozzi, E.; Boe, A.; et al. Cancer Stem Cell-Based Models of Colorectal Cancer Reveal Molecular Determinants of Therapy Resistance. Stem Cells Transl. Med. 2016, 5, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Katsiampoura, A.; Raghav, K.; Jiang, Z.-Q.; Menter, D.G.; Varkaris, A.; Morelli, M.P.; Manuel, S.; Wu, J.; Sorokin, A.V.; Rizi, B.S.; et al. Modeling of Patient-Derived Xenografts in Colorectal Cancer. Mol. Cancer Ther. 2017, 16, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Viqueira, B.; Jimeno, A.; Cusatis, G.; Zhang, X.; Iacobuzio-Donahue, C.; Karikari, C.; Shi, C.; Danenberg, K.; Danenberg, P.V.; Kuramochi, H.; et al. An In vivo Platform for Translational DrugDevelopment in Pancreatic Cancer. Clin. Cancer Res. 2006, 12, 4652–4661. [Google Scholar] [CrossRef]
- Hidalgo, M.; Bruckheimer, E.; Rajesh Kumar, N.V.; Garrido-Laguna, I.; De Oliveira, E.; Rubio-Viqueira, B.; Strawn, S.; Wick, M.J.; Martell, J.; Sidransky, D. A pilot clinical study of treatment guided by personalized tumor grafts in patients with advanced cancer. Mol. Cancer Ther. 2011, 10, 1311–1316. [Google Scholar] [CrossRef]
- Stebbing, J.; Paz, K.; Schwartz, G.K.; Wexler, L.H.; Maki, R.G.; Pollock, R.E.; Morris, R.; Cohen, R.; Shankar, A.; Blackman, G.; et al. Patient-derived xenografts for individualized care in advanced sarcoma. Cancer 2014, 120, 2006–2015. [Google Scholar] [CrossRef] [PubMed]
- Fiore, D.; Di Giacomo, F.; Kyriakides, P.; Inghirami, G. Patient-Derived-Tumor-Xenograft: Modeling cancer for basic and translational cancer research. Clin. Diagn. Pathol. 2017, 1, 1. [Google Scholar] [CrossRef][Green Version]
- Pauli, C.; Hopkins, B.D.; Prandi, D.; Shaw, R.; Fedrizzi, T.; Sboner, A.; Sailer, V.; Augello, M.; Puca, L.; Rosati, R.; et al. Personalized In Vitro and In vivo Cancer Models to Guide Precision Medicine. Cancer Discov. 2017, 7, 462–477. [Google Scholar] [CrossRef]
- Rajcevic, U.; Knol, J.C.; Piersma, S.; Bougnaud, S.; Fack, F.; Sundlisaeter, E.; Søndenaa, K.; Myklebust, R.; Pham, T.V.; Niclou, S.P.; et al. Colorectal cancer derived organotypic spheroids maintain essential tissue characteristics but adapt their metabolism in culture. Proteome Sci. 2014, 12, 39. [Google Scholar] [CrossRef] [PubMed]
- Park, J.I.; Lee, J.; Kwon, J.L.; Park, H.B.; Lee, S.Y.; Kim, J.Y.; Sung, J.; Kim, J.M.; Song, K.S.; Kim, K.H. Scaffold-Free Coculture Spheroids of Human Colonic Adenocarcinoma Cells and Normal Colonic Fibroblasts Promote Tumorigenicity in Nude Mice. Transl. Oncol. 2016, 9, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Bauman, E.; Feijão, T.; Carvalho, D.; Granja, P.L.; Barrias, C.C. Xeno-free pre-vascularized spheroids for therapeutic applications. Sci. Rep. 2018, 8, 230. [Google Scholar] [CrossRef]
- Gheytanchi, E.; Naseri, M.; Karimi-Busheri, F.; Atyabi, F.; Mirsharif, E.S.; Bozorgmehr, M.; Ghods, R.; Madjd, Z. Morphological and molecular characteristics of spheroid formation in HT-29 and Caco-2 colorectal cancer cell lines. Cancer Cell Int. 2021, 21, 204. [Google Scholar] [CrossRef]
- Francescangeli, F.; Patrizii, M.; Signore, M.; Federici, G.; Di Franco, S.; Pagliuca, A.; Baiocchi, M.; Biffoni, M.; Ricci Vitiani, L.; Todaro, M.; et al. Proliferation state and polo-like kinase1 dependence of tumorigenic colon cancer cells. Stem Cells 2012, 30, 1819–1830. [Google Scholar] [CrossRef]
- Correr, S.; Makabe, S.; Heyn, R.; Relucenti, M.; Naguro, T.; Familiari, G. Microplicae-like structures of the fallopian tube in postmenopausal women as shown by electron microscopy. Histol. Histopathol. 2006, 21, 219–226. [Google Scholar]
- Cottignoli, V.; Relucenti, M.; Agrosì, G.; Cavarretta, E.; Familiari, G.; Salvador, L.; Maras, A. Biological Niches within Human Calcified Aortic Valves: Towards Understanding of the Pathological Biomineralization Process. BioMed Res. Int. 2015, 542687. [Google Scholar] [CrossRef] [PubMed]
- Lo Torto, F.; Relucenti, M.; Familiari, G.; Vaia, N.; Casella, D.; Matassa, R.; Miglietta, S.; Marinozzi, F.; Bini, F.; Fratoddi, I.; et al. The Effect of Postmastectomy Radiation Therapy on Breast Implants: Material Analysis on Silicone and Polyurethane Prosthesis. Ann. Plast. Surg. 2018, 81, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Forte, M.; Bianchi, F.; Cotugno, M.; Marchitti, S.; De Falco, E.; Raffa, S.; Stanzione, R.; Di Nonno, F.; Chimenti, I.; Palmerio, S.; et al. Pharmacological restoration of autophagy reduces hypertension-related stroke occurrence. Autophagy 2020, 16, 1468–1481. [Google Scholar] [CrossRef]
- Relucenti, M.; Heyn, R.; Petruzziello, L.; Pugliese, G.; Taurino, M.; Familiari, G. Detecting microcalcifications in atherosclerotic plaques by a simple trichromic staining method for epoxy embedded carotid endarterectomies. Eur. J. Histochem. 2010, 54, e33. [Google Scholar] [CrossRef]
- Relucenti, M.; Miglietta, S.; Bove, G.; Donfrancesco, O.; Battaglione, E.; Familiari, P.; Barbaranelli, C.; Covelli, E.; Barbara, M.; Familiari, G. SEM BSE 3D Image Analysis of Human Incus Bone Affected by Cholesteatoma Ascribes to Osteoclasts the Bone Erosion and VpSEM dEDX Analysis Reveals New Bone Formation. Scanning 2020, 9371516. [Google Scholar] [CrossRef] [PubMed]
- Dolznig, H.; Rupp, C.; Puri, C.; Haslinger, C.; Schweifer, N.; Wieser, E.; Kerjaschki, D.; Garin-Chesa, P. Modeling colon adenocarcinomas in vitro a 3D co-culture system induces cancer-relevant pathways upon tumor cell and stromal fibroblast interaction. Am. J. Pathol. 2011, 179, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Han, S.J.; Kwon, S.; Kim, K.S. Challenges of applying multicellular tumor spheroids in preclinical phase. Cancer Cell Int. 2021, 21, 152. [Google Scholar] [CrossRef]
- Olzmann, J.A.; Carvalho, P. Dynamics and functions of lipid droplets. Nat. Rev. Mol. Cell Biol. 2019, 20, 137–155. [Google Scholar] [CrossRef]
- Cruz, A.L.S.; Barreto, E.d.A.; Fazolini, N.; Viola, J.; Bozza, P.B. Lipid droplets: Platforms with multiple functions in cancer hallmarks. Cell Death Dis. 2020, 11, 105. [Google Scholar] [CrossRef]
- Ricci-Vitiani, L.; Lombardi, D.G.; Pilozzi, E.; Biffoni, M.; Todaro, M.; Peschle, C.; De Maria, R. Identification and expansion of human colon-cancer-initiating cells. Nature 2007, 445, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, L.; Todaro, M.; de Sousa Mello, F.; Sprick, M.R.; Kemper, K.; Perez Alea, M.; Richel, D.J.; Stassi, G.; Medema, J.P. Single-cell cloning of colon cancer stem cells reveals a multi-lineage differentiation capacity. In Proceedings of the National Academy of Sciences of the United States of America, 12 August 2008; Volume 105, pp. 13427–13432. [Google Scholar]
- Dieter, S.M.; Ball, C.R.; Hoffmann, C.M.; Nowrouzi, A.; Herbst, F.; Zavidij, O.; Abel, U.; Arens, A.; Weichert, W.; Brand, K.; et al. Distinct types of tumor-initiating cells form human colon cancer tumors and metastases. Cell Stem Cell 2011, 9, 357–365. [Google Scholar] [CrossRef]
- Vermeulen, L.; De Sousa, E.; Melo, F.; van der Heijden, M.; Cameron, K.; de Jong, J.H.; Borovski, T.; Tuynman, J.B.; Todaro, M.; Merz, C.; et al. Wnt activity defines colon cancer stem cells and is regulated by the microenvironment. Nature Cell Biol. 2010, 12, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Todaro, M.; Gaggianesi, M.; Catalano, V.; Benfante, A.; Iovino, F.; Biffoni, M.; Apuzzo, T.; Sperduti, I.; Volpe, S.; Cocorullo, G.; et al. CD44v6 is a marker of constitutive and reprogrammed cancer stem cells driving colon cancer metastasis. Cell Stem Cell 2014, 14, 342–356. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Compartment | Morphological Parameters |
|---|---|
| Membrane | Microvilli, apical binding complex; basal and lateral domains |
| Cytoplasm | Types and morphology of organelles, filaments and inclusions. |
| Nucleus | Shape, number of nucleoli; chromatin aspect |
| Descriptors | Mean | Std. Error | 95% CI |
|---|---|---|---|
| Circularity | 0.94 | 0.006 | 0.92 to 0.95 |
| Roundness | 0.87 | 0.008 | 0.86 to 0.89 |
| Aspect Ratio | 1.17 | 0.02 | 1.12 to 1.22 |
| Solidity | 0.99 | 0.001 | 0.98 to 0.99 |
| Parameter | Mean | Std. Error | 95% CI |
|---|---|---|---|
| Area | 967.75 µm2 | 59.95 µm | 844.26 to 1091.24 µm |
| Perimeter | 112.21 µm | 3.46 µm | 105.07 to 119.35 µm |
| Feret diameter | 39.21 µm | 1.51 µm | 36.10 to 42.32 µm |
| Min Feret diameter | 32.84 µm | 1.11 µm | 30.55 to 35.14 µm |
| Factor | Mean | Std. Error | 95% CI |
|---|---|---|---|
| Spheroid cell type A | 7.1250 | 0.7425 | 5.3692 to 8.8808 |
| Spheroid cell type B | 11.1250 | 1.7159 | 7.0676 to 15.1824 |
| Spheroid cell type C | 10.3750 | 1.5691 | 6.6647 to 14.0853 |
| Spheroid cell type D | 8.6250 | 1.4134 | 5.2828 to 11.9672 |
| Spheroid cell type E | 1.2500 | 0.3660 | 0.3846 to 2.1154 |
| Xenograft cell type A | 1.6250 | 0.3750 | 0.7383 to 2.5117 |
| Xenograft cell type B | 9.6250 | 1.3879 | 6.3431 to 12.9069 |
| Xenograft cell type C | 10.1250 | 1.5748 | 6.4013 to 13.8487 |
| Xenograft cell type D | 10.1250 | 1.3016 | 7.0472 to 13.2028 |
| Xenograft cell type E | 5.7500 | 0.4119 | 4.7761 to 6.7239 |
| Source of Variation | Sum of Squares | DF | Mean Square | F | P |
|---|---|---|---|---|---|
| Sphericity assumed | 941,550 | 9 | 104,671 | 9.69 | <0.001 |
| Greenhouse Geisser | 941,550 | 3.711 | 253,701 | 9.69 | <0.001 |
| Huynh-Feldt | 941,550 | 8.420 | 111,827 | 9.69 | <0.001 |
| Factor | Mean Difference | Std. Error | p | 95% CI | |
|---|---|---|---|---|---|
| Spheroid cell type A vs. | Xenograft cell type A | 5.500 | 0.732 | 0.0061 | 1.613 to 9.387 |
| Spheroid cell type B vs. | Xenograft cell type B | 1.500 | 2.619 | 1.0000 | −12.405 to 15.405 |
| Spheroid cell type C vs. | Xenograft cell type C | 0.250 | 1.461 | 1.0000 | −7.507 to 8.007 |
| Spheroid cell type D vs. | Xenograft cell type D | −1.500 | 1.195 | 1.0000 | −7.847 to 4.847 |
| Spheroid cell type E vs. | Xenograft cell type E | −4.500 | 0.732 | 0.0211 | −8.387 to −0.613 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Relucenti, M.; Francescangeli, F.; De Angelis, M.L.; D’Andrea, V.; Miglietta, S.; Pilozzi, E.; Li, X.; Boe, A.; Chen, R.; Zeuner, A.; et al. The Ultrastructural Analysis of Human Colorectal Cancer Stem Cell-Derived Spheroids and Their Mouse Xenograft Shows That the Same Cells Types Have Different Ratios. Biology 2021, 10, 929. https://doi.org/10.3390/biology10090929
Relucenti M, Francescangeli F, De Angelis ML, D’Andrea V, Miglietta S, Pilozzi E, Li X, Boe A, Chen R, Zeuner A, et al. The Ultrastructural Analysis of Human Colorectal Cancer Stem Cell-Derived Spheroids and Their Mouse Xenograft Shows That the Same Cells Types Have Different Ratios. Biology. 2021; 10(9):929. https://doi.org/10.3390/biology10090929
Chicago/Turabian StyleRelucenti, Michela, Federica Francescangeli, Maria Laura De Angelis, Vito D’Andrea, Selenia Miglietta, Emanuela Pilozzi, Xiaobo Li, Alessandra Boe, Rui Chen, Ann Zeuner, and et al. 2021. "The Ultrastructural Analysis of Human Colorectal Cancer Stem Cell-Derived Spheroids and Their Mouse Xenograft Shows That the Same Cells Types Have Different Ratios" Biology 10, no. 9: 929. https://doi.org/10.3390/biology10090929
APA StyleRelucenti, M., Francescangeli, F., De Angelis, M. L., D’Andrea, V., Miglietta, S., Pilozzi, E., Li, X., Boe, A., Chen, R., Zeuner, A., & Familiari, G. (2021). The Ultrastructural Analysis of Human Colorectal Cancer Stem Cell-Derived Spheroids and Their Mouse Xenograft Shows That the Same Cells Types Have Different Ratios. Biology, 10(9), 929. https://doi.org/10.3390/biology10090929