
Cloning and Functional Verification of CYP408A3 and CYP6CS3 Related to Chlorpyrifos Resistance in the Sogatella furcifera (Horváth) (Hemiptera: Delphacidae)

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Insecticides and Chemicals
2.3. Bioassay
2.4. Quantitative PCR (RT-qPCR) for Insecticide Resistant-Related P450 Genes
2.5. Cloning of CYP Genes Sequences
2.6. Analysis of CYP Genes DNA and Protein Sequences
2.7. Preparation of dsRNA
2.8. dsRNA Microinjection
2.9. Assessment of Nymph Mortality and Cytochrome P450 Activity
2.10. Data Analysis
3. Results
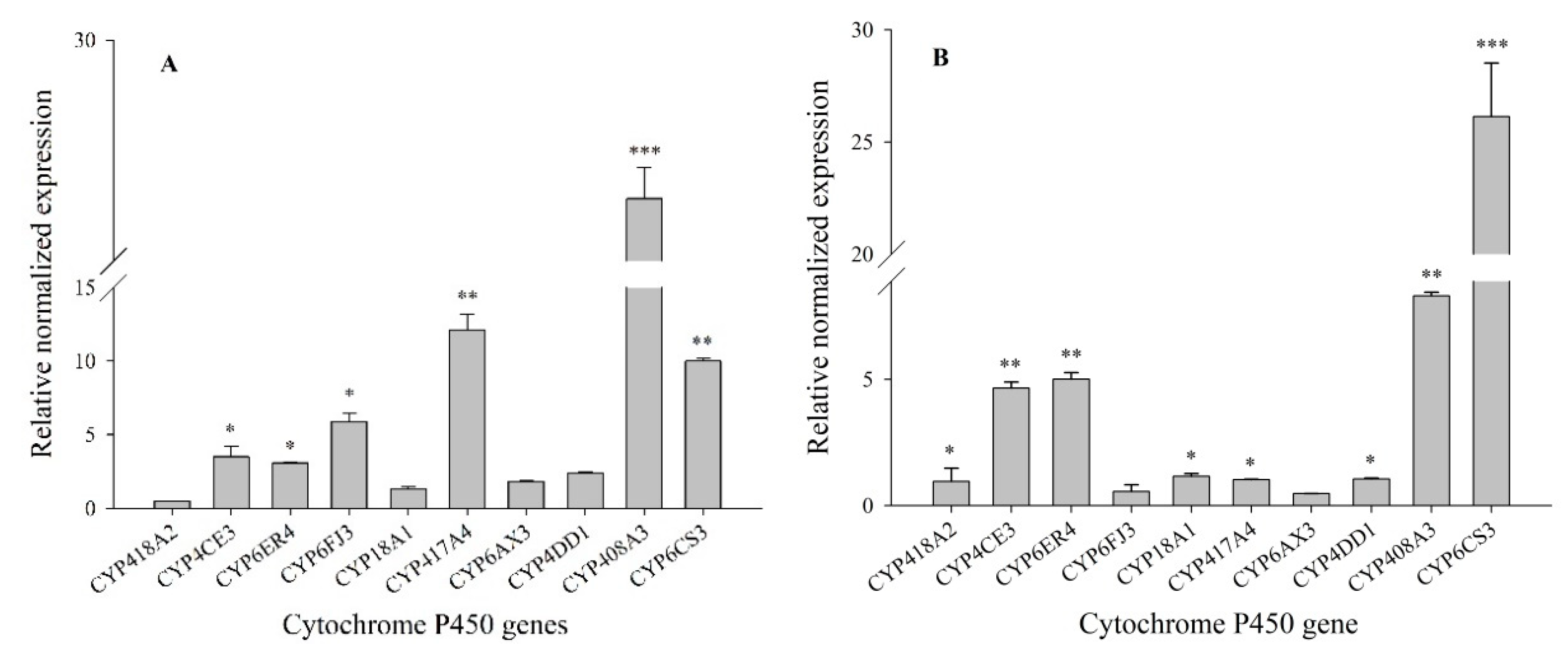
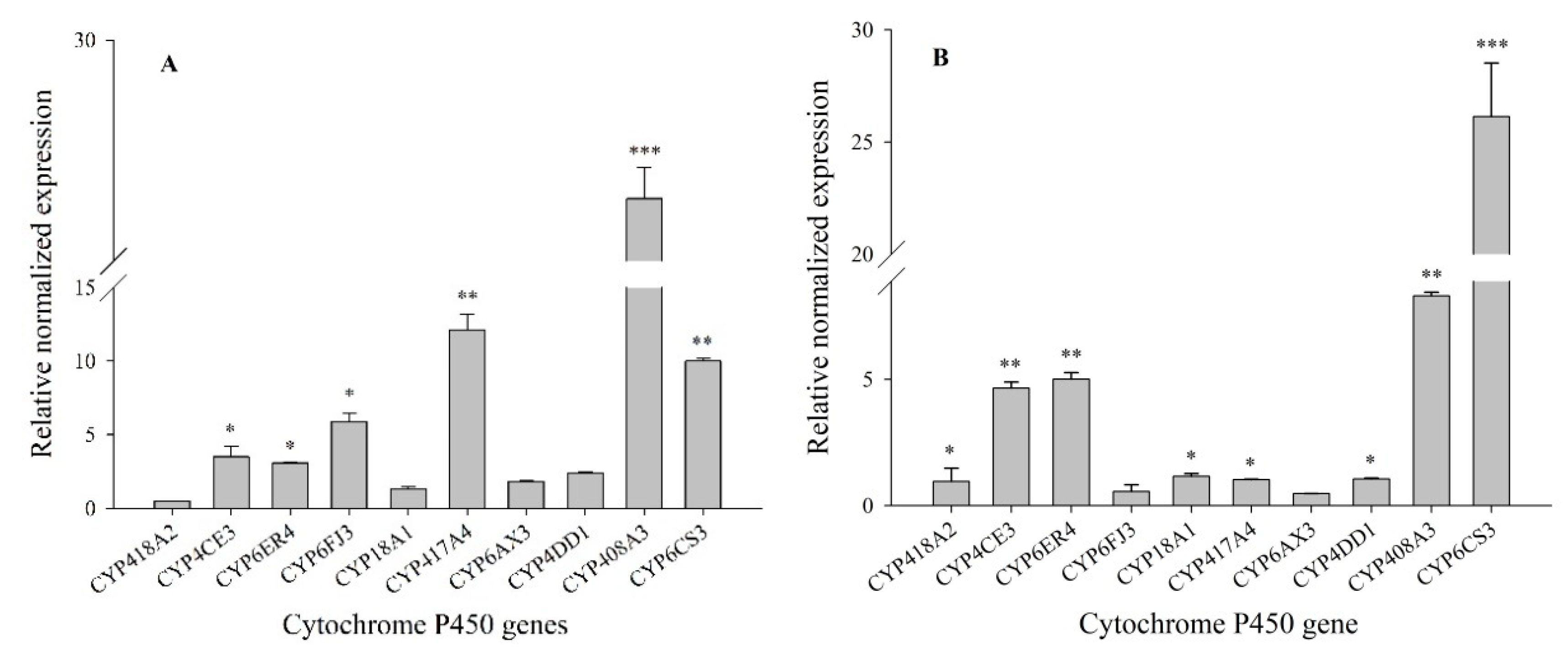
3.1. Expression Profiles of P450 mRNAs in Lab-HN and XY17 Populations of Sogatella furcifera
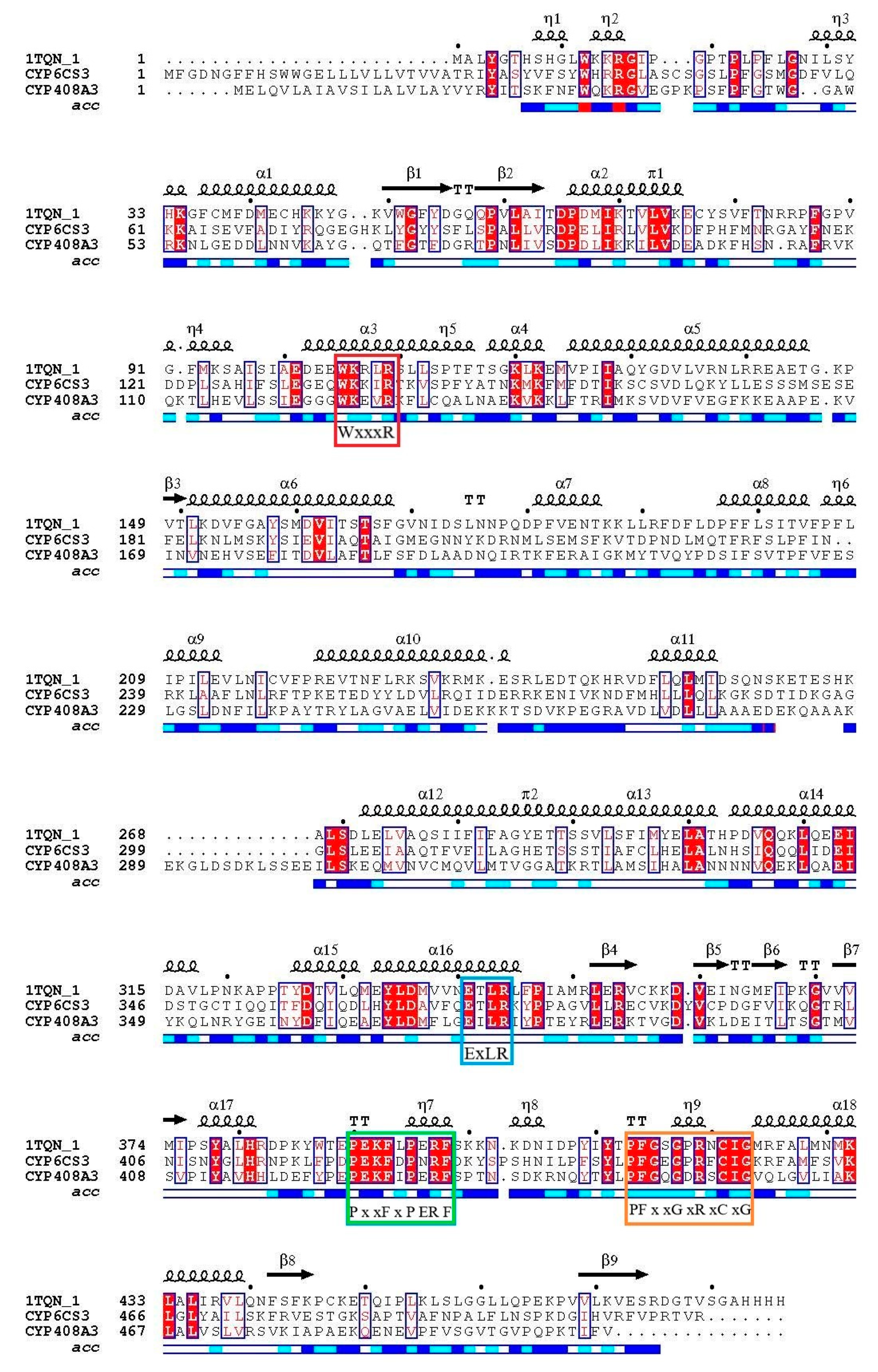
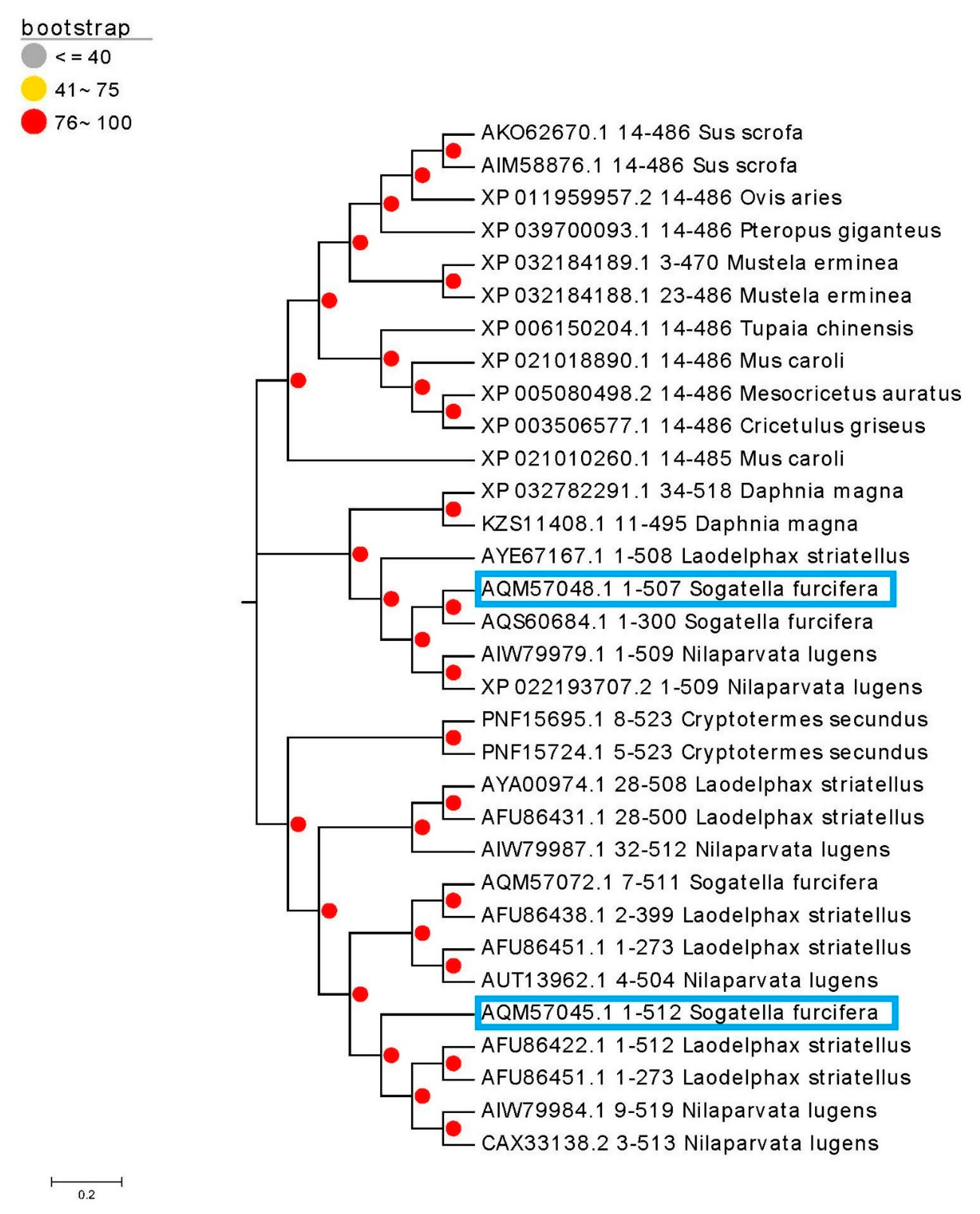
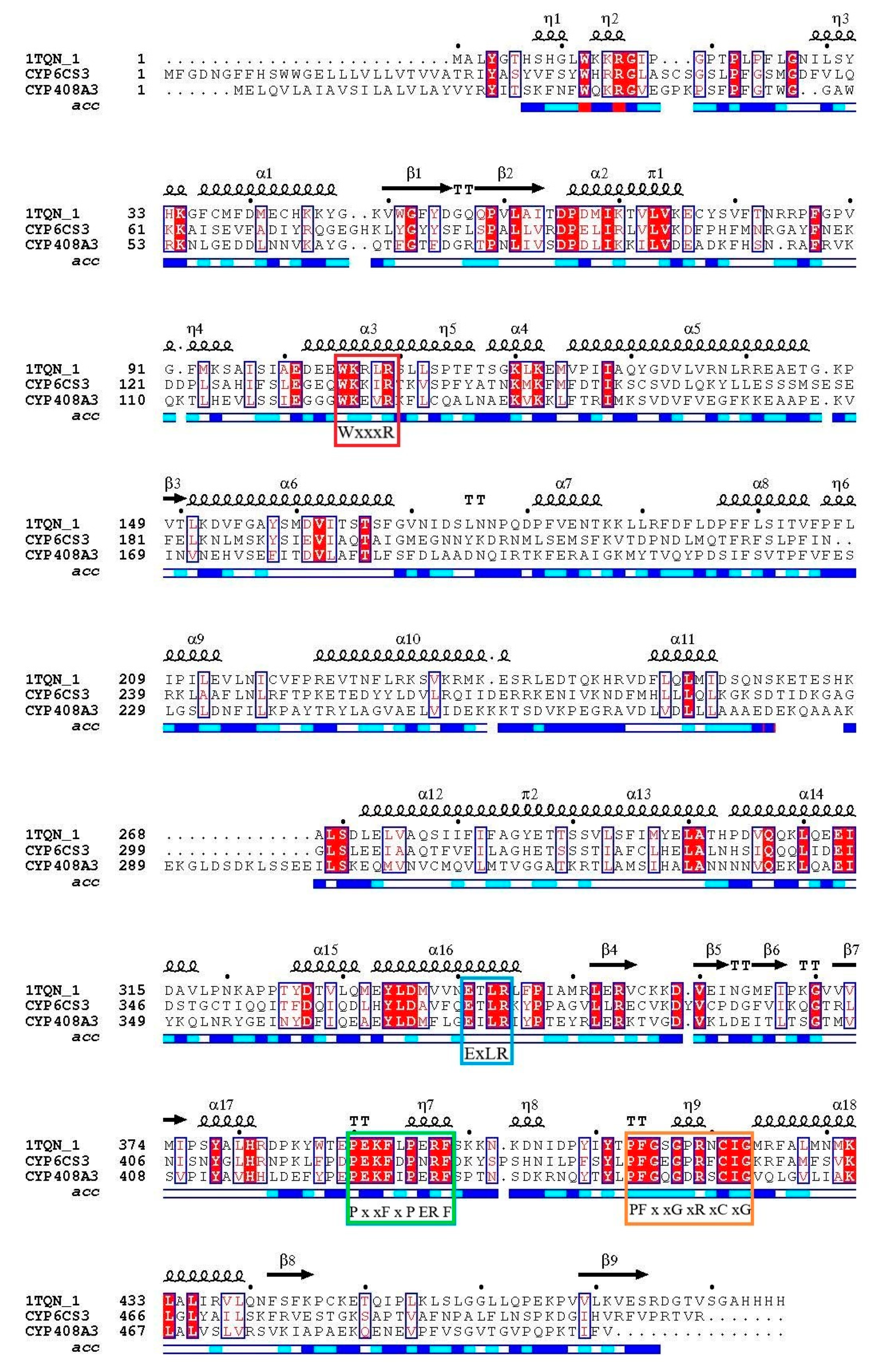
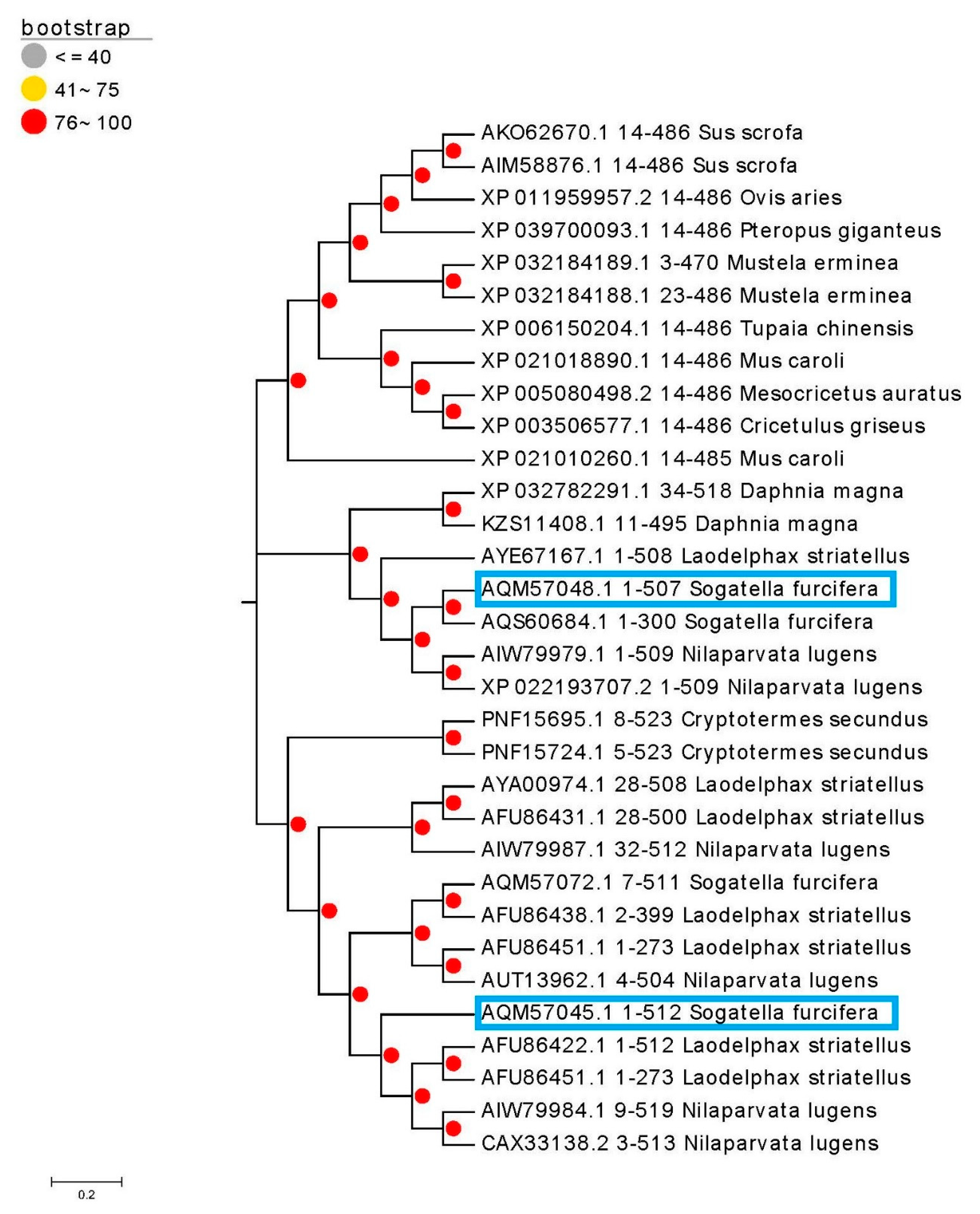
3.2. Characterization of Full-Length Amino Acid Sequences of Two Cloned P450 Genes
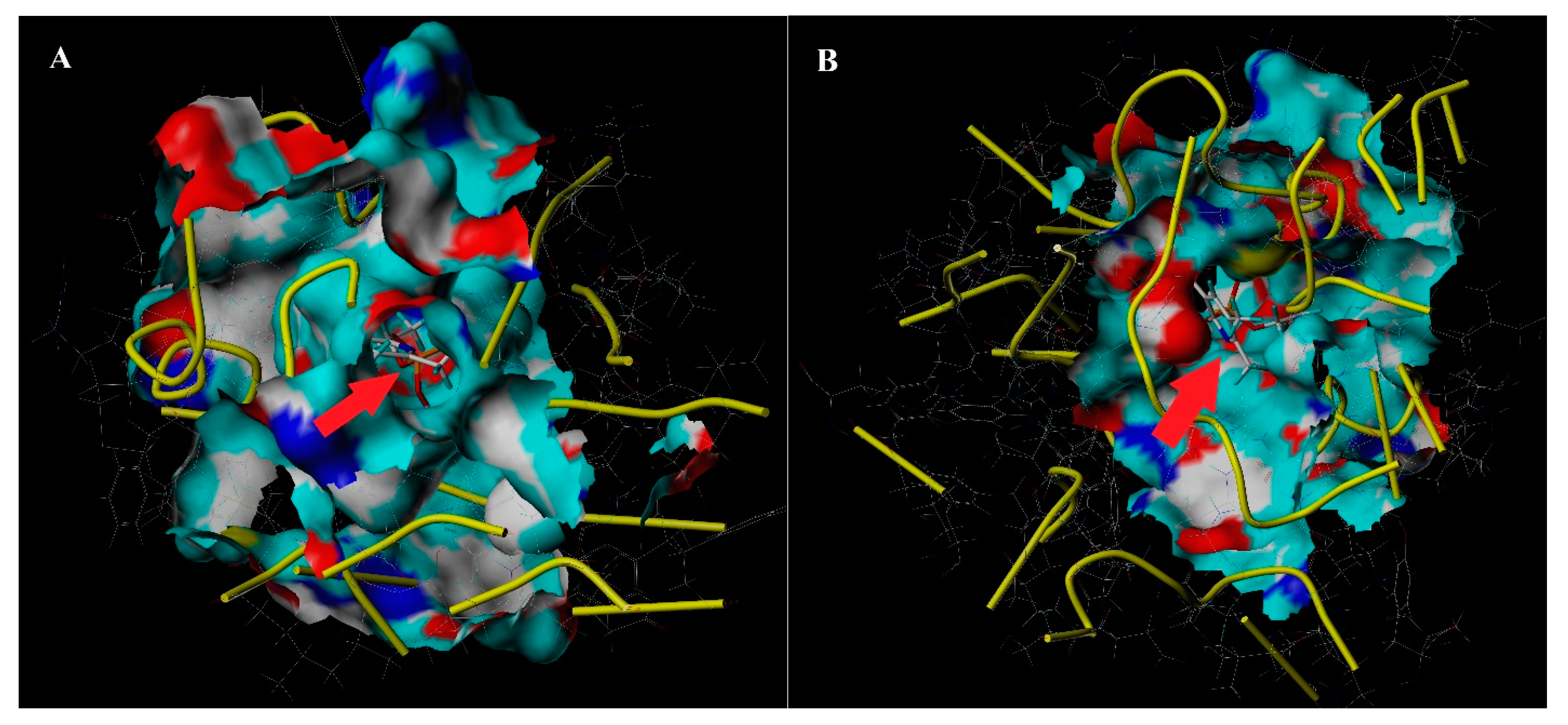
3.3. Interaction of the Tertiary Structure of CYP6CS3 and CYP408A3 with Chlorpyrifos
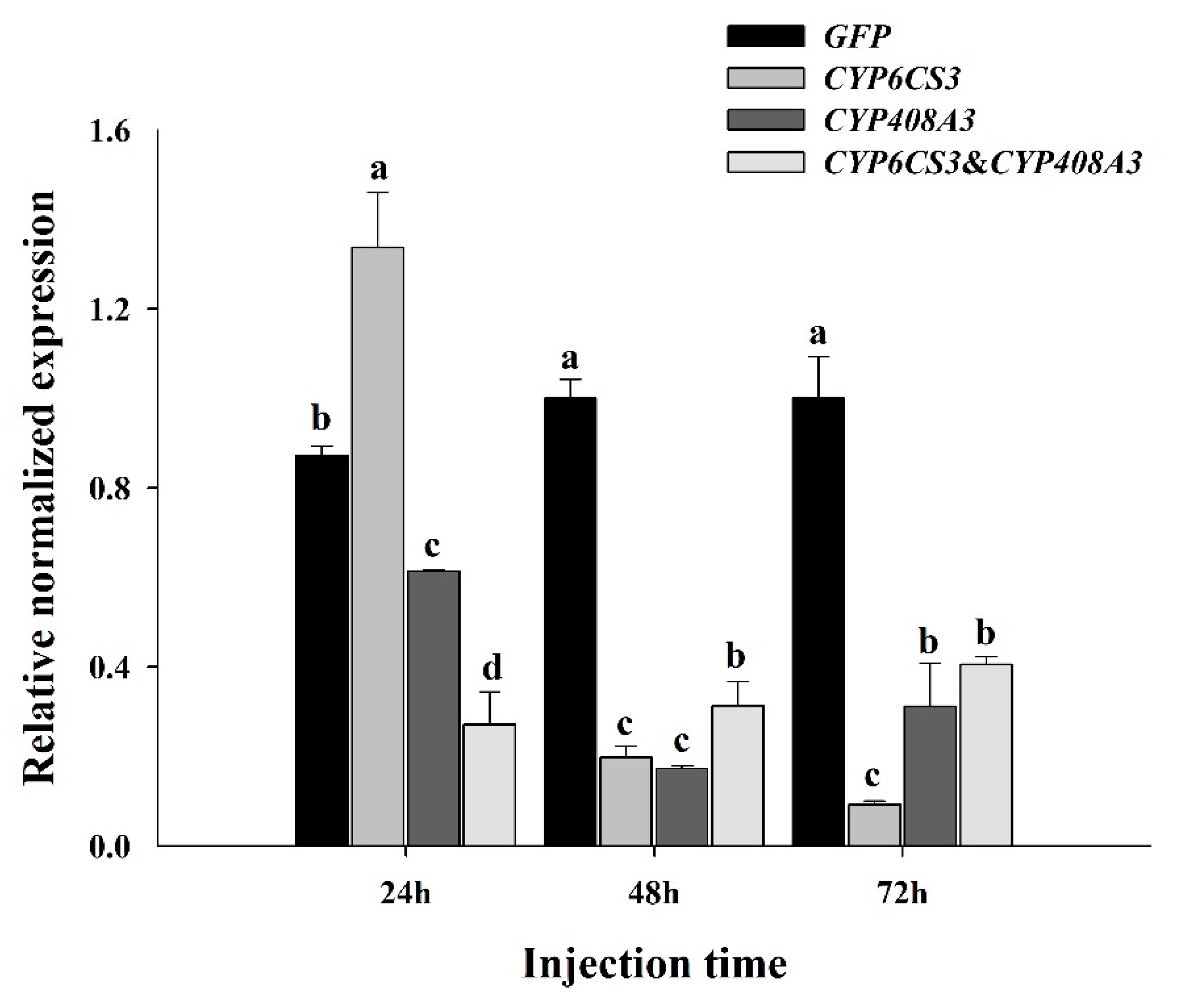
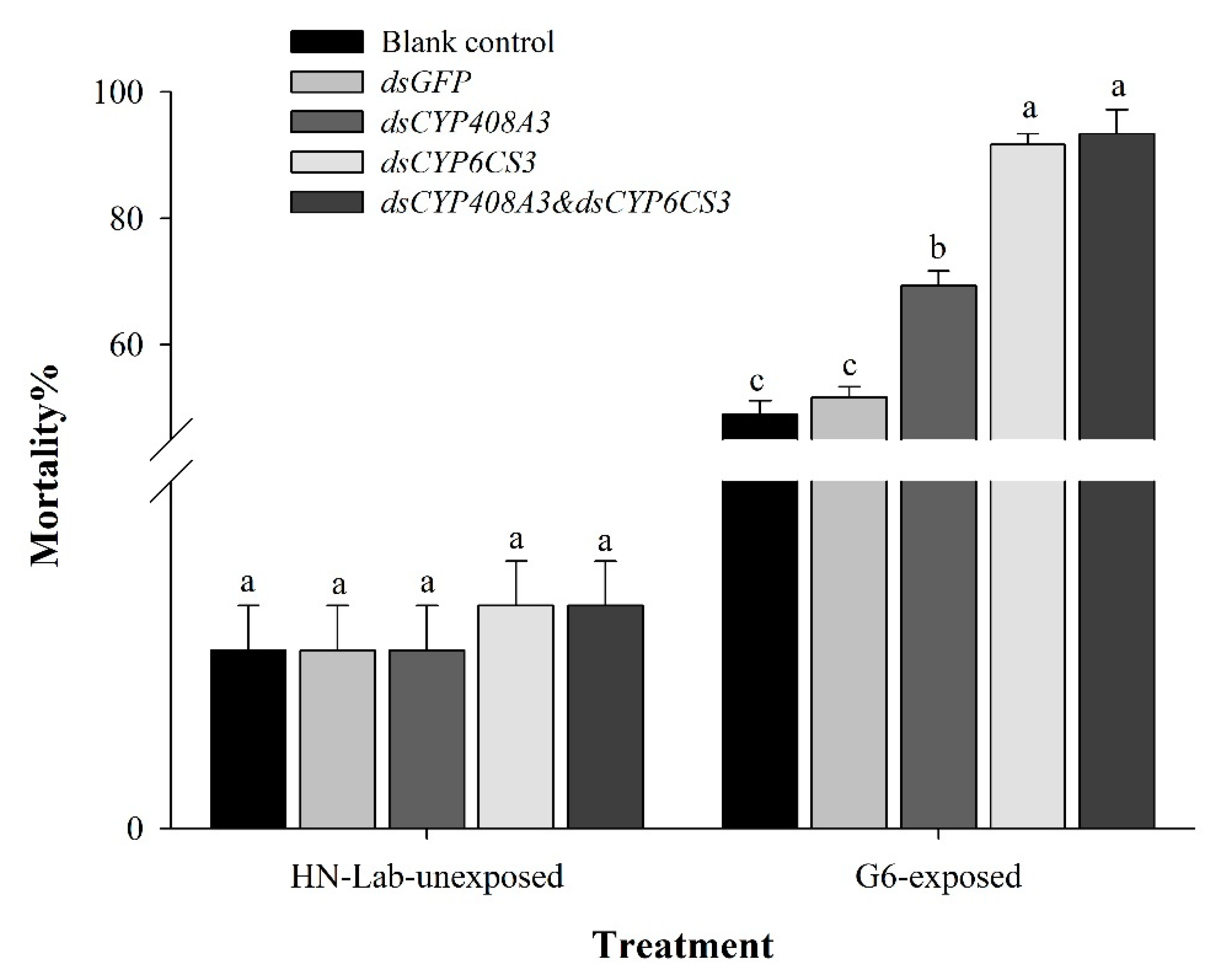
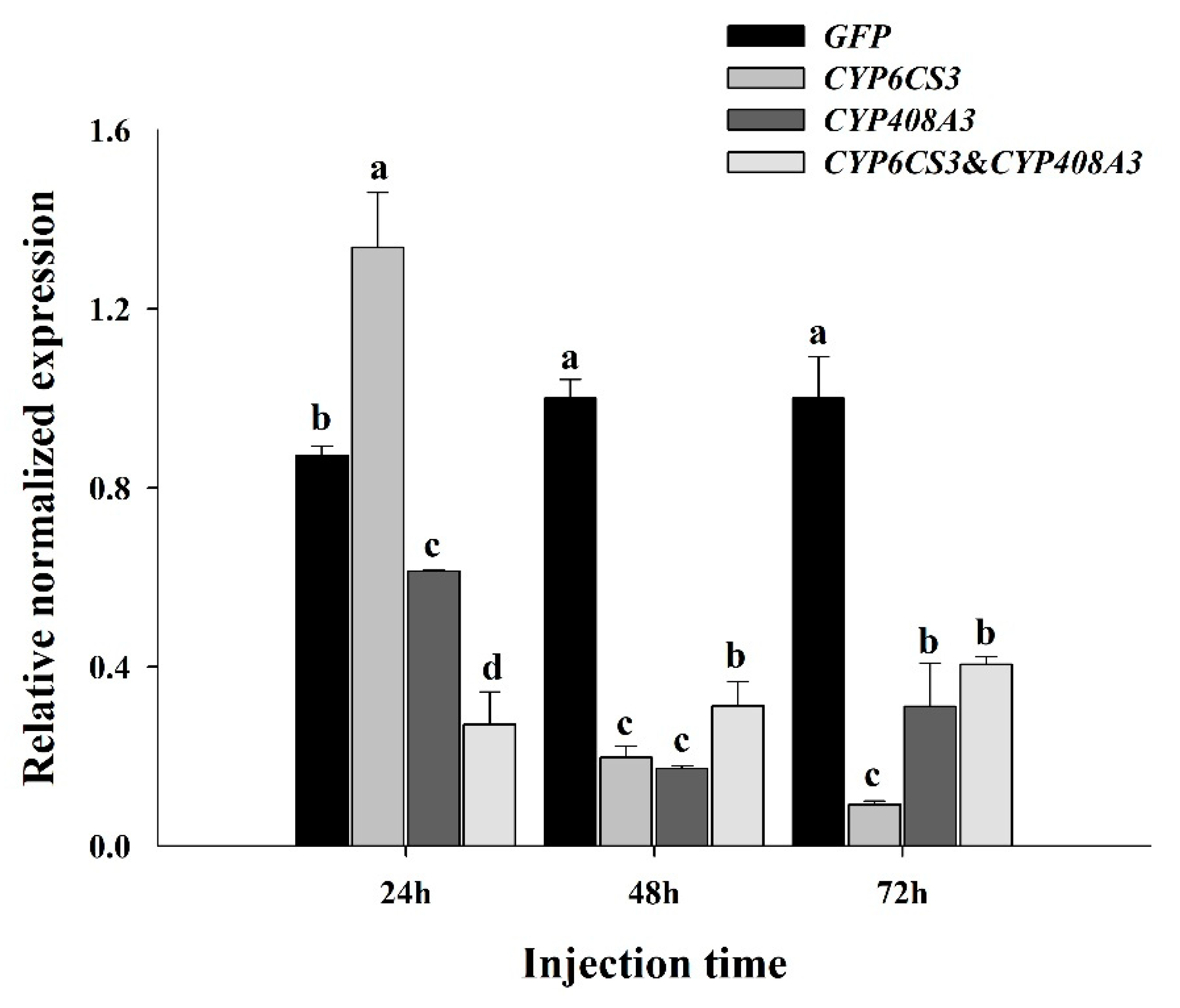
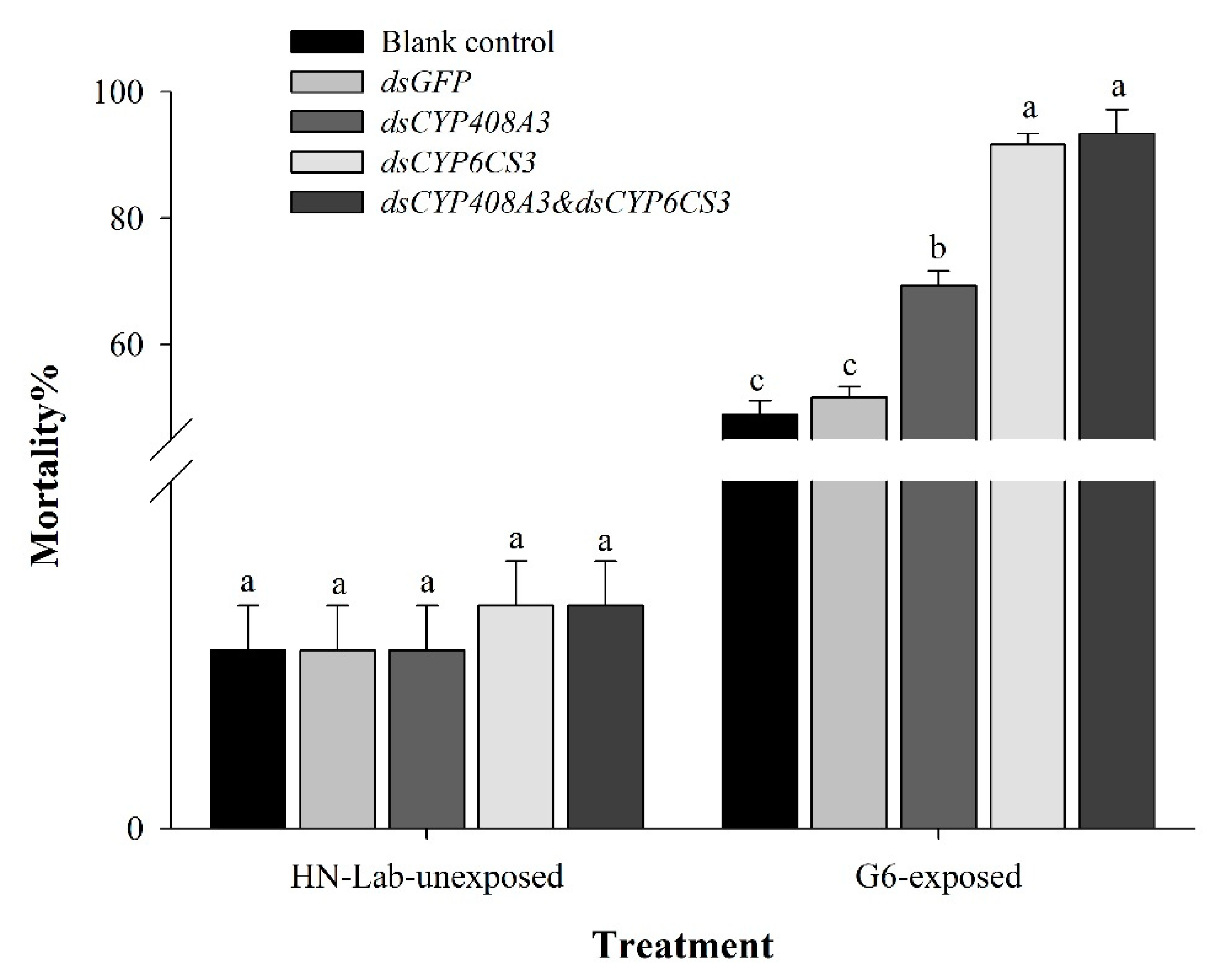
3.4. Functional Analysis of CYP6CS3 and CYP408A3 via RNAi
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sogawa, K. Vulnerability to insect pests in Chinese hybrid rice. Agric. Tech. 2001, 56, 398–402. [Google Scholar]
- Zhou, G.H.; Wen, J.J.; Cai, D.J. Southern rice black-streaked dwarf virus: A new proposed Fijivirus species in the family reoviridae. Sci. Bull. 2008, 53, 3677–3685. [Google Scholar] [CrossRef]
- Cheng, Z.N.; Li, S.; Gao, R.Z.; Sun, F.; Liu, W.C.; Zhou, G.H.; Wu, J.X.; Zhou, X.P.; Zhou, Y.J. Distribution and genetic diversity of southern rice black-streaked dwarf virus in China. Virol. J. 2013, 10, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsukura, K.; Towata, T.; Sakai, J.; Onuki, M.; Okuda, M.; Matsumura, M. Dynamics of Southern rice black-streaked dwarf virus in rice and implication for virus acquisition. Phytopathology 2013, 5, 509–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.G.; Xiang, X.; Yu, H.L.; Liu, S.H.; Yin, Y.; Cui, P.; Wu, Y.Q.; Yang, J.; Jiang, C.X.; Yang, Q.F. Monitoring and biochemical characterization of beta-cypermethrin resistance in Spodoptera exigua (Lepidoptera: Noctuidae) in Sichuan Province, China. Pestic. Biochem. Physiol. 2018, 146, 71–79. [Google Scholar] [CrossRef]
- Liu, M.G.; Jiang, C.X.; Mao, M.; Liu, C.; Li, Q.; Wang, X.G.; Yang, Q.F.; Wang, H.J. Effect of the insecticide dinotefuran on the ultrastructure of the flight muscle of female Sogatella furcifera (Hemiptera: Delphacidae). J. Econ. Entomol. 2017, 110, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; He, J.Y.; Liang, A.; Wang, F.H. Transcriptome profiling and dimorphic expression of sex-related genes in fifth-instar nymphs of Sogatella furcifera, an important rice pest. Genomics 2020, 112, 1105–1111. [Google Scholar] [CrossRef]
- Matsumura, M.; Takeuchi, H.; Satoh, M.; Sanada-Morimura, S.; Otuka, A.; Watanabe, T.; Thanh, D.V. Species-specific insecticide resistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in east and southeast Asia. Pest Manag. Sci. 2008, 64, 1115–1121. [Google Scholar] [CrossRef]
- Tang, J.; Li, J.; Shao, Y.; Yang, B.J.; Liu, Z.W. Fipronil resistance in the white-backed planthopper (Sogatella furcifera): Possible resistance mechanisms and cross-resistance. Pest Manag. Sci. 2010, 66, 121–125. [Google Scholar] [CrossRef]
- Matsumura, M.; Sanada-Morimura, S.; Otuka, A.; Ohtsu, R.; Sakumoto, S.; Takeuchia, H.; Satoha, M. Insecticide susceptibilities in populations of two rice planthoppers, Nilaparvata lugens and Sogatella furcifera, immigrating into Japan in the period 2005–2012. Pest Manag. Sci. 2014, 70, 615–622. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, W.; Zhang, S.; Wu, S.F.; Ban, L.F.; Su, J.Y.; Gao, C.F. Susceptibility of Sogatella furcifera and Laodelphax striatellus (Hemiptera: Delphacidae) to six insecticides in China. J. Econ. Entomol. 2014, 107, 1916–1922. [Google Scholar] [CrossRef]
- Mu, X.C.; Zhang, W.; Wang, L.X.; Zhang, S.; Zhang, K.; Gao, C.F.; Wu, S.F. Resistance monitoring and cross-resistance patterns of three rice planthoppers, Nilaparvata lugens, Sogatella furcifera and Laodelphax striatellus to dinotefuran in China. Pestic. Biochem. Physiol. 2016, 134, 8–13. [Google Scholar] [CrossRef]
- Pope, C.; Karanth, S.; Liu, J. Pharmacology and toxicology of cholinesterase inhibitors: Uses and misuses of a common mechanism of action. Environ. Toxicol. Pharmacol. 2005, 19, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.X.; Jin, D.C.; Li, W.H.; Cheng, Y.; Li, F.L.; Ye, Z.C. Monitoring trends in insecticide resistance of field populations of Sogatella furcifera (Hemiptera: Delphacidae) in Guizhou Province, China, 2012–2015. J. Econ. Entomol. 2017, 110, 641–650. [Google Scholar] [CrossRef]
- Ali, E.; Mao, K.K.; Liao, X.; Jin, R.H.; Li, J.H. Cross-resistance biochemical characterization of buprofezin resistance in the white-backed planthopper, Sogatella furcifera (Horváth). Pestic. Biochem. Physiol. 2019, 158, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Liu, S.H.; Wang, X.G.; Zhang, Y.M.; Gong, C.W.; Chen, L.; Zhang, S.R.; Shen, L.T. Sublethal effects of sulfoxaflor on population projection and development of the white-backed planthopper, Sogatella furcifera (Hemiptera: Delphacidae). Crop Prot. 2019, 120, 97–102. [Google Scholar] [CrossRef]
- Yu, L. New York state will phase out chlorpyrifos in 2021. J. Agrochem. Mark. 2019, 10, 49. [Google Scholar]
- Yang, G. Vietnam will ban Ethyl Chlorpyrifos and Fipronil after 2 years later. Pestic. Mark. News 2019, 6, 48. [Google Scholar]
- Bhattacharyya, A.; Bhaumik, A.; Rani, P.U.; Mandal, S.; Epidi, T.T. Nano-particles-A recent approach to insect pest control. Afr. J. Biotechnol. 2010, 24, 3489–3493. [Google Scholar]
- Su, J.Y.; Wang, Z.W.; Zhang, K.; Tian, X.G.; Yin, Y.Q.; Zhao, X.Q.; Shen, A.; Gao, C.F. Study on insecticide resistance of white-backed planthopper. Fla. Entomol. Soc. 2013, 96, 948–956. [Google Scholar] [CrossRef]
- Mohan, M.; Gujar, G.T. Local variation in susceptibility of the diamondback moth, Plutella xylostella (Linnaeus) to insecticides and role of detoxification enzymes. Crop Prot. 2009, 22, 495–504. [Google Scholar] [CrossRef]
- Lai, T.C.; Li, J.; Su, J.Y. Monitoring of beet armyworm Spodoptera exigua (Lepidoptera: Noctuidae) resistance to chlorantraniliprole in China. Pestic. Biochem. Physiol. 2011, 101, 198–205. [Google Scholar] [CrossRef]
- Wang, X.G.; Huang, Q.; Hao, Q.; Ran, S.; Wu, Y.Q.; Cui, P.; Yang, J.; Jiang, C.X.; Yang, Q.F. Insecticide resistance and enhanced cytochrome P450 monooxygenase activity in field populations of Spodoptera litura from Sichuan, China. Crop Prot. 2018, 106, 110–116. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Yang, Y.X.; Sun, H.H.; Liu, Z.W. Metabolic imidacloprid resistance in the brown planthopper, Nilaparvata lugens, relies on multiple P450 enzymes. Int. J. Mol. Sci. 2016, 79, 50–56. [Google Scholar] [CrossRef]
- Gao, B.L.; Wu, J.; Huang, S.J.; Mu, L.F.; Han, Z.J. Insecticide resistance in field populations of Laodelphax striatellus Fallén (Homoptera: Delphacidae) in China and its possible mechanisms. Int. J. Pest Manag. 2008, 54, 13–19. [Google Scholar] [CrossRef]
- Bao, H.B.; Gao, H.L.; Zhang, Y.X.; Fan, D.; Fang, J.; Liu, Z. The roles of CYP6AY1 and CYP6ER1 in imidacloprid resistance in the brown planthopper: Expression levels and detoxification efficiency. Pestic. Biochem. Physiol. 2016, 129, 70–74. [Google Scholar] [CrossRef]
- Yang, Y.X.; Zhang, Y.X.; Yang, B.J.; Fang, J.C.; Liu, Z.W. Transcriptomic responses to different doses of cycloxaprid involved in detoxification and stress response in the whitebacked planthopper, Sogatella furcifera. Entomol. Exp. Appl. 2016, 158, 248–257. [Google Scholar] [CrossRef]
- Liao, X.; Jin, R.G.; Zhang, X.L.; Ali, E.; Mao, K.K.; Xu, P.F.; Li, J.H.; Wan, H. Characterization of sulfoxaflor resistance in the brown planthopper, Nilaparvata lugens. Pest Manag. Sci. 2019, 75, 1646–1654. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Wu, M.; Han, Z.J. Biochemical and molecular characterisation and cross-resistance in field and laboratory chlorpyrifos-resistant strains of Laodelphax striatellus (Hemiptera: Delphacidae) from eastern China. Pest Manag. Sci. 2014, 70, 1118–1129. [Google Scholar] [CrossRef] [PubMed]
- Mao, K.K.; Zhang, X.L.; Ali, E.; Liao, X.; Jin, R.H.; Ren, Z.J.; Wan, H.; Li, J.H. Characterization of nitenpyram resistance in Nilaparvata lugens (Stål). Pestic. Biochem. Physiol. 2019, 157, 26–32. [Google Scholar] [CrossRef]
- Chen, L.; Wang, X.; Zhang, Y.; Yang, R.; Zhang, S.; Xu, X.; Jiang, C.X. The population growth, development and metabolic enzymes of the white-backed planthopper, Sogatella furcifera (Hemiptera: Delphacidae) under the sublethal dose of triflumezopyrim. Chemosphere 2020, 247, 125865. [Google Scholar] [CrossRef]
- Wang, L.; Tang, N.; Gao, X.L.; Chang, Z.X.; Zhang, L.G.; Zhou, G.H.; Guo, D.Y.; Zeng, Z.; Li, W.J.; Akinyemi, I.A.; et al. Genome sequence of a rice pest, the white-backed planthopper (Sogatella furcifera). GigaScience 2017, 6, 1–9. [Google Scholar]
- An, X.K.; Hou, M.L.; Liu, Y.D. Reference gene selection and evaluation for gene expression studies using RT-qPCR in the white-backed Planthopper, Sogatella furcifera (Hemiptera: Delphacidae). J. Econ. Entomol. 2016, 109, 879. [Google Scholar] [CrossRef]
- Wang, X.G.; Ruan, Y.W.; Gong, C.W.; Xiang, X.; Xu, X.; Zhang, Y.M.; Shen, L.T. Transcriptome analysis of Sogatella furcifera (Homoptera: Delphacidae) in response to sulfoxaflor and functional verification of resistance-related P450 genes. Int. J. Mol. Sci. 2019, 20, 4573. [Google Scholar] [CrossRef] [Green Version]
- Lertkiatmongkol, P.; Jenwitheesuk, E.; Rongnoparut, P. Homology modeling of mosquito cytochrome P450 enzymes involved in pyrethroid metabolism, insights into differences in substrate selectivity. BMC Res. Notes 2011, 4, 321. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhang, Y.; Wang, Y.; Yang, Y.; Cang, X.; Liu, Z. Expression induction of P450 genes by imidacloprid in Nilaparvata lugens: A genome-scale analysis. Pestic. Biochem. Physiol. 2016, 132, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Rose, R.L.; Barbhaiya, L.; Roe, R.M. Cytochrome P450-associated insecticide resistance and the development of biochemical diagnostic assays in Heliothis virescens. Pestic. Biochem. Physiol. 1995, 51, 178–191. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, C.; Long, G.Y.; Yang, H.; Jin, D.C. Sublethal effects of buprofezin on development, reproduction, and chitin synthase 1 gene (SfCHS1) expression in the white-backed planthopper, Sogatella furcifera (Hemiptera: Delphacidae). J. Asia Pac. Entomol. 2018, 21, 585–591. [Google Scholar] [CrossRef]
- Bradford, M.M.A. A rapid and sensitive method for quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 25, 248–256. [Google Scholar] [CrossRef]
- Robertson, J.L.; Preisler, H.K. Pesticide Bioassays with Arthropods. Fla. Entomol. 2008, 91, 510–511. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR, the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Preetha, G.; Stanley, J.; Suresh, S.; Samiyappan, R. Risk assessment of insecticides used in rice on miridbug, Cyrtorhinus lividipennis Reuter, the important predator of brown planthopper, Nilaparvata lugens (Stål.). Chemosphere 2010, 80, 498–503. [Google Scholar] [CrossRef]
- Owolade, O.; Ogunleti, D. Bffects of Titanium Dioxide on The Diseases, Development and yield ofedible cowpea. J. Plant. Prot. Res. 2008, 48, 3. [Google Scholar] [CrossRef]
- Li, W.H.; Mao, K.K.; Liu, C.Y.; Gong, P.P.; Xu, P.F.; Wu, G.; Le, W.; Wan, H.; You, H.; Li, J.H. Resistance monitoring and assessment of the control failure likelihood of insecticides in field populations of the whitebacked planthopper Sogatella furcifera (Horváth). Crop Prot. 2020, 127, 104973. [Google Scholar] [CrossRef]
- Liu, N.N.; Li, M.; Gong, Y.H.; Liu, F.; Li, T. Cytochrome P450s-Their expression, regulation, and role in insecticide resistance. Pestic. Biochem. Physiol. 2015, 120, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Jin, R.; Mao, K.; Liao, X.; Xu, P.; Li, Z.; Ali, E.; Li, J. Overexpression of CYP6ER1 associated with clothianidin resistance in Nilaparvata lugens (Stål). Pestic. Biochem. Physiol. 2019, 154, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.W.; Wang, X.G.; Xiang, X. Status of insecticide resistance and biochemical characterization of chlorpyrifos resistance in Sogatella furcifera (Horváth) in Sichuan Province, China. Pestic. Biochem. Physiol. 2020, 171, 104723. [Google Scholar] [CrossRef]
- Scott, J.G. Cytochromes P450 and insecticide resistance. Insect Biochem. Mol. Biol. 1999, 29, 757–777. [Google Scholar] [CrossRef]
- Karunker, I.; Benting, J.; Lueke, B.; Ponge, T.; Nauen, R.; Roditakis, E.; Vontas, J.; Gorman, K.; Denholm, I.; Morin, S. Over-expression of cytochrome P450 CYP6CM1 is associated with high resistance to imidacloprid in the B and Q biotypes of Bemisia tabaci (Hemiptera: Aleyrodidae). Insect Biochem. Mol. 2008, 38, 634–644. [Google Scholar] [CrossRef]
- Zhou, C.; Yang, H.; Wang, Z.; Long, G.Y.; Jin, D.C. Comparative transcriptome analysis of Sogatella furcifera (Horváth) exposed to different insecticides. Sci. Rep. 2018, 8, 8773. [Google Scholar] [CrossRef]
- Okey, A.B. Enzyme induction in the cytochrome P-450 system. Pharmacol. Ther. 1990, 45, 241–298. [Google Scholar] [CrossRef]
- Scott, J.G.; Michel, K.; Bartholomay, L.C.; Siegfried, B.D.; Hunter, W.B.; Smagghe, G.; Zhu, K.Y.; Douglas, A.E. Towards the elements of successful insect RNAi. J. Insect Physiol. 2013, 59, 1212–1221. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.X.; Liu, Y.B.; Rong, W.H. RNA-seq and its application in transcriptome research. Genetics 2011, 33, 1191–1202. [Google Scholar]
- Zhu, K.Y. RNA interference: A powerful tool in entomological research and a novel approach for insect pest management. Insect Sci. 2013, 20, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.H.; Wu, M.; Pu, J.; Feng, A.; Zhang, Q.; Han, Z.J. A functional study of two dsRNA binding protein genes in Laodelphax striatellus. Pest Manag. Sci. 2013, 69, 1034–1039. [Google Scholar] [CrossRef] [PubMed]
- Denlinger, D.L.; Yocum, G.D.; Rinehart, J.P. Hormonal control of diapause. Compr. Insect Mol. Sci. 2005, 3, 615–650. [Google Scholar]
- Stiborová, M.; Indra, R.; Frei, E.; Schmeiser, H.H.; Eckschlager, T.; Adam, V.; Heger, Z.; Arlt, V.M.; Martínek, V. Cytochrome b5 plays a dual role in the reaction cycle of cytochrome P450 3A4 during oxidation of the anticancer drug ellipticine. Monatsh. Chem. 2017, 148, 1983–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupasinghe, S.G.; Wen, Z.M.; Chiu, T.L.; Schuler, M.A. Helicoverpa zea CYP6B8 and CYP321A1: Different molecular solutions to the problem of metabolizing plant toxins and insecticides. Protein Eng. Des. Sel. 2007, 20, 615–624. [Google Scholar] [CrossRef]
- Cui, S.F.; Wang, L.; Ma, L.; Geng, X.Q. P450-mediated detoxification of botanicals in insects. Phytoparasitica 2016, 44, 585–599. [Google Scholar] [CrossRef]
- Thomas, J.H. Rapid birth-death evolution specific to xenobiotic cytochrome P450 genes in vertebrates. PLoS Genet. 2007, 3, e6. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Wu, M.; Han, Z.J. Overexpression of Multiple Detoxification Genes in Deltamethrin Resistant Laodelphax striatellus (Hemiptera: Delphacidae) in China. PLoS ONE 2013, 8, e79443. [Google Scholar] [CrossRef] [PubMed]
- Lao, S.H.; Huang, X.H.; Huang, H.J.; Liu, C.W.; Zhang, C.X.; Bao, Y.Y. Genomic and transcriptomic insights into the cytochrome P450 monooxygenase gene repertoire in the rice pest brown planthopper, Nilaparvata lugens. Genomics 2015, 106, 301–309. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | P450 Activity | SR a | |
|---|---|---|---|
| nmol·(min·mg·pro)−1 | |||
| Lab-HN | 2.42 ± 0.007 | c | 1.0 |
| XY17-G6 | 3.42 ± 0.02 | a | 1.41 |
| XY17-G6-dsCYP6CS3 | 2.08 ± 0.04 | e | 0.86 |
| XY17-G6-dsCYP408A3 | 2.24 ± 0.19 | d | 0.93 |
| XY17-G6- dsCYP6CS3 & dsCYP408A3 | 2.29 ± 0.07 | cd | 0.95 |
| XY17-G6-dsGFP | 2.96 ± 0.02 | b | 1.22 |
| F4, 10 = 184.468, p < 0.001 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruan, Y.; Liu, X.; Gong, C.; Zhang, Y.; Shen, L.; Ali, H.; Huang, Y.; Wang, X. Cloning and Functional Verification of CYP408A3 and CYP6CS3 Related to Chlorpyrifos Resistance in the Sogatella furcifera (Horváth) (Hemiptera: Delphacidae). Biology 2021, 10, 795. https://doi.org/10.3390/biology10080795
Ruan Y, Liu X, Gong C, Zhang Y, Shen L, Ali H, Huang Y, Wang X. Cloning and Functional Verification of CYP408A3 and CYP6CS3 Related to Chlorpyrifos Resistance in the Sogatella furcifera (Horváth) (Hemiptera: Delphacidae). Biology. 2021; 10(8):795. https://doi.org/10.3390/biology10080795
Chicago/Turabian StyleRuan, Yanwei, Xinxian Liu, Changwei Gong, Yuming Zhang, Litao Shen, Hasnain Ali, Yanyan Huang, and Xuegui Wang. 2021. "Cloning and Functional Verification of CYP408A3 and CYP6CS3 Related to Chlorpyrifos Resistance in the Sogatella furcifera (Horváth) (Hemiptera: Delphacidae)" Biology 10, no. 8: 795. https://doi.org/10.3390/biology10080795
APA StyleRuan, Y., Liu, X., Gong, C., Zhang, Y., Shen, L., Ali, H., Huang, Y., & Wang, X. (2021). Cloning and Functional Verification of CYP408A3 and CYP6CS3 Related to Chlorpyrifos Resistance in the Sogatella furcifera (Horváth) (Hemiptera: Delphacidae). Biology, 10(8), 795. https://doi.org/10.3390/biology10080795