
Mimicking of Blood Flow Results in a Distinct Functional Phenotype in Human Non-Adherent Classical Monocytes

 , , and
, , and
Abstract
:Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
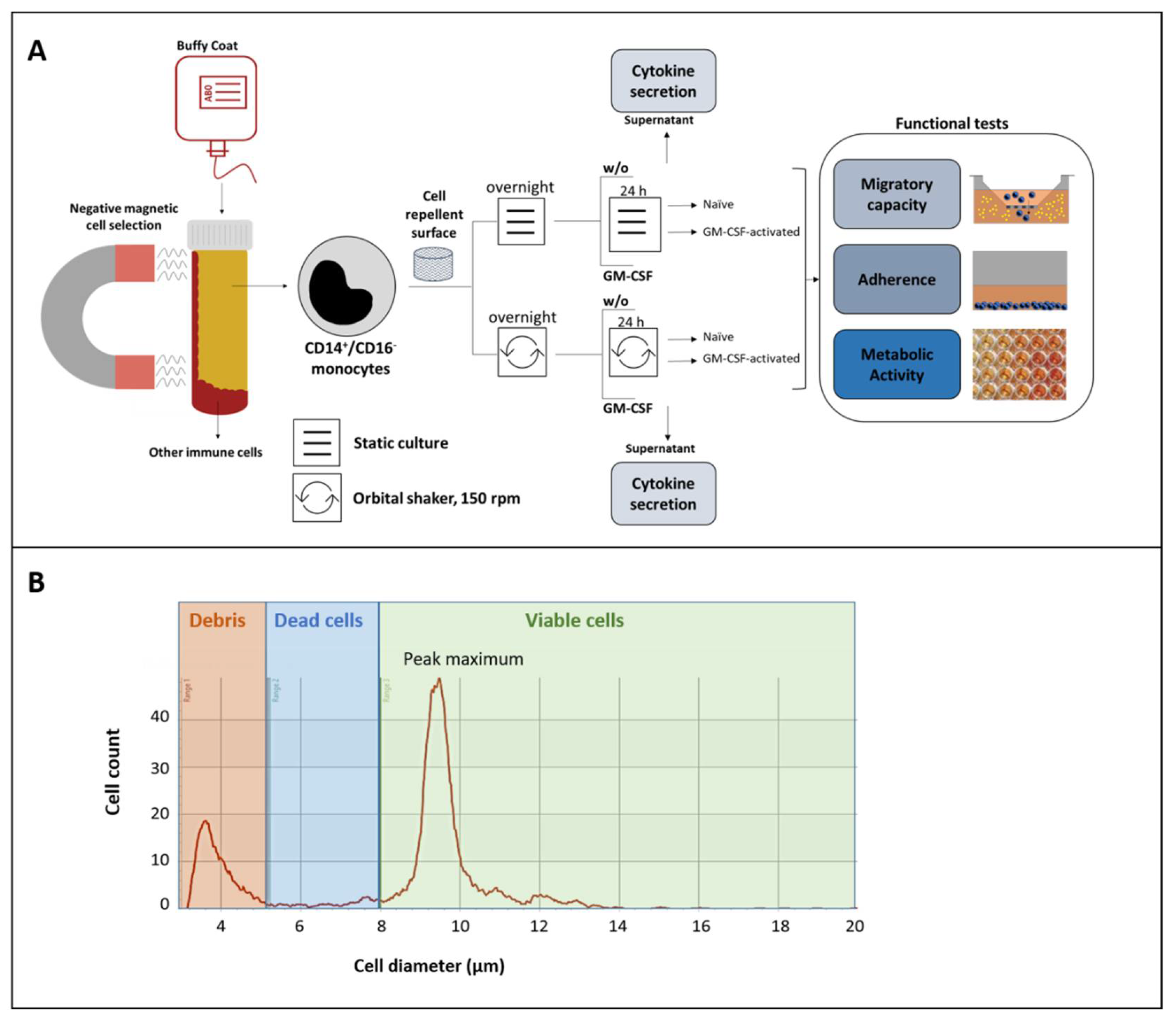
2.1. Human Monocytes
2.2. Experimental Design
2.3. Monitoring of Cell Number, Cell Size, and Viability
2.4. Quantification of Cytokine Release
2.5. Migratory Capacity
2.6. Adherence
2.7. Quantification of Metabolic Activity
2.8. Statistical Analyses
3. Results
3.1. Shear Flow Affects Cytokine Release and Cell Size of Non-Adherent Monocytes
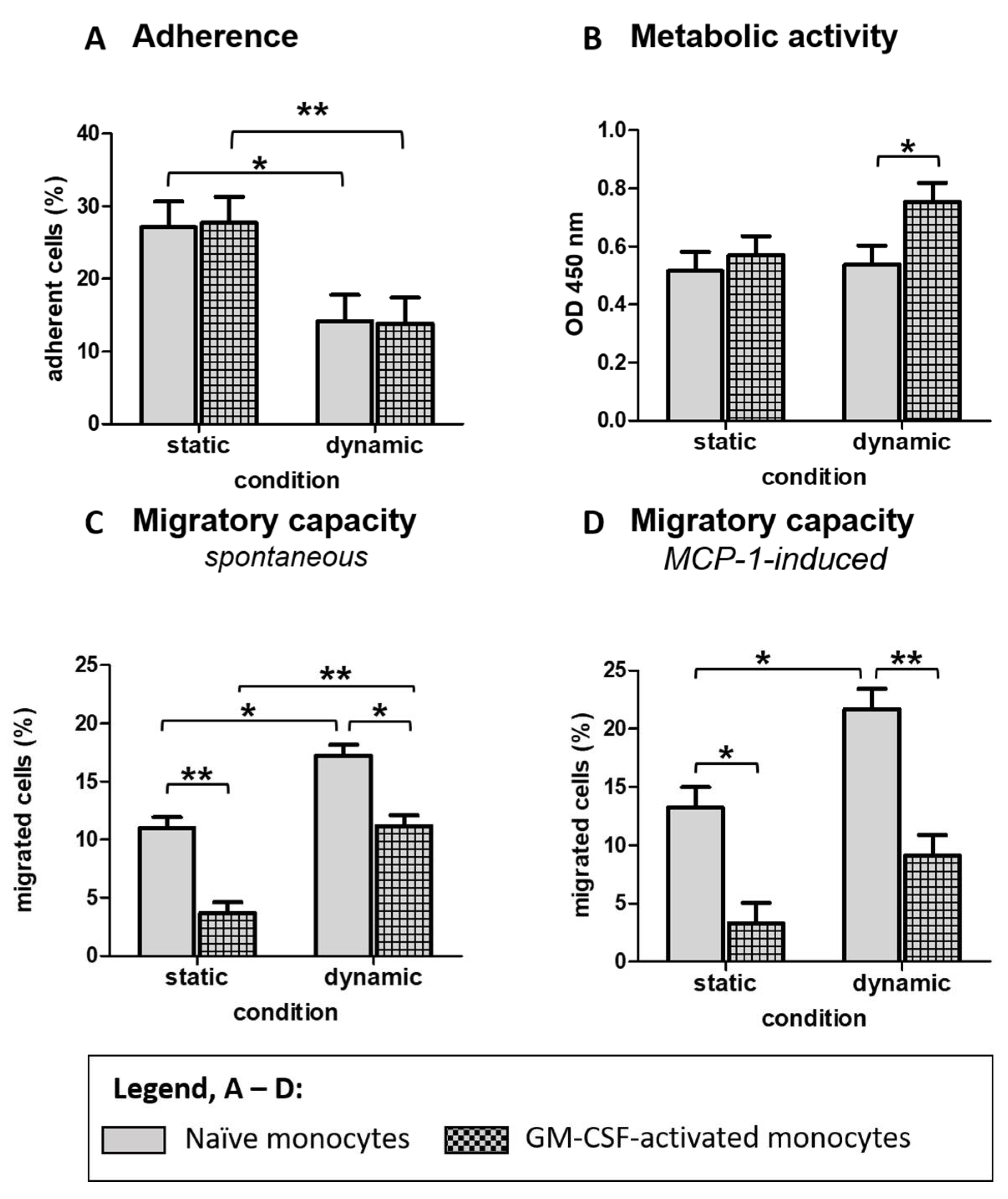
3.2. Shear Flow Affects Functional Properties of Non-Adherent Monocytes Related to Adhesion Cascade and Energy Metabolism
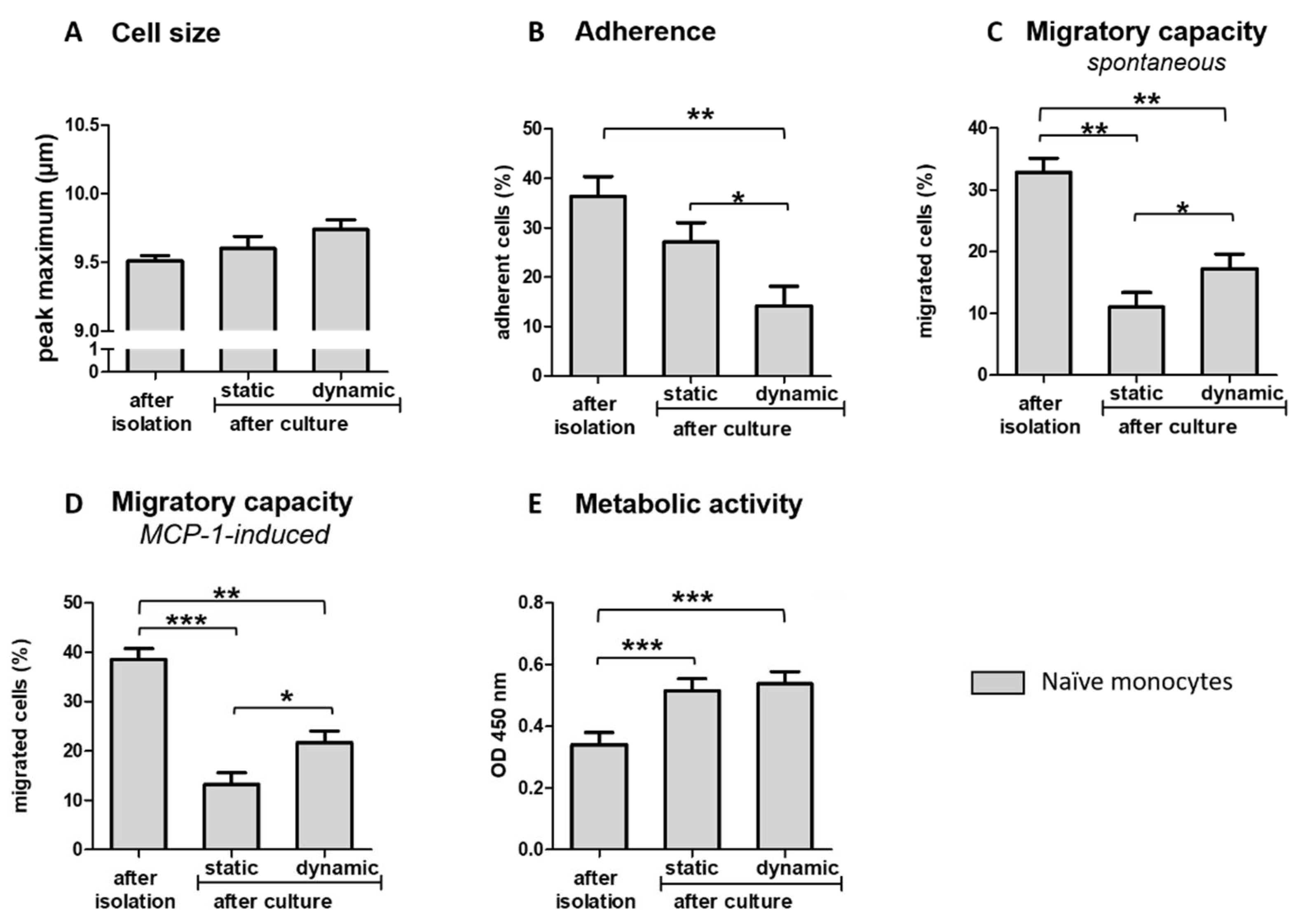
3.3. Ex Vivo Incubation Influences Functional Properties of Blood-Derived Monocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coillard, A.; Segura, E. In vivo Differentiation of Human Monocytes. Front. Immunol. 2019, 10, 1907. [Google Scholar] [CrossRef]
- Jakubzick, C.V.; Randolph, G.J.; Henson, P.M. Monocyte differentiation and antigen-presenting functions. Nat. Rev. Immunol. 2017, 17, 349–362. [Google Scholar] [CrossRef]
- Auffray, C.; Fogg, D.; Garfa, M.; Elain, G.; Join-Lambert, O.; Kayal, S.; Sarnacki, S.; Cumano, A.; Lauvau, G.; Geissmann, F. Monitoring of blood vessels and tissues by a population of monocytes with patrolling behavior. Science 2007, 317, 666–670. [Google Scholar] [CrossRef] [Green Version]
- Kapellos, T.S.; Bonaguro, L.; Gemünd, I.; Reusch, N.; Saglam, A.; Hinkley, E.R.; Schultze, J.L. Human Monocyte Subsets and Phenotypes in Major Chronic Inflammatory Diseases. Front. Immunol. 2019, 10, 2035. [Google Scholar] [CrossRef] [Green Version]
- Sampath, P.; Moideen, K.; Ranganathan, U.D.; Bethunaickan, R. Monocyte Subsets: Phenotypes and Function in Tuberculosis Infection. Front. Immunol. 2018, 9, 1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Däbritz, J.; Weinhage, T.; Varga, G.; Wirth, T.; Walscheid, K.; Brockhausen, A.; Schwarzmaier, D.; Brückner, M.; Ross, M.; Bettenworth, D.; et al. Reprogramming of monocytes by GM-CSF contributes to regulatory immune functions during intestinal inflammation. J. Immunol. 2015, 194, 2424–2438. [Google Scholar] [CrossRef] [Green Version]
- Weinhage, T.; Däbritz, J.; Brockhausen, A.; Wirth, T.; Brückner, M.; Belz, M.; Foell, D.; Varga, G. Granulocyte macrophage colony-stimulating factor-activated CD39+/CD73+ murine monocytes modulate intestinal inflammation via induction of regulatory T cells. Cell. Mol. Gastroenterol. Hepatol. 2015, 1, 433–449. [Google Scholar] [CrossRef] [Green Version]
- Herold, J.; Tillmanns, H.; Xing, Z.; Strasser, R.H.; Braun-Dullaeus, R.C. Isolation and transduction of monocytes: Promising vehicles for therapeutic arteriogenesis. Langenbeck’s Arch. Surg. 2006, 391, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Lebson, L.; Nash, K.; Kamath, S.; Herber, D.; Carty, N.; Lee, D.C.; Li, Q.; Szekeres, K.; Jinwal, U.; Koren, J. Trafficking CD11b-positive blood cells deliver therapeutic genes to the brain of amyloid-depositing transgenic mice. J. Neurosci. 2010, 30, 9651–9658. [Google Scholar] [CrossRef] [PubMed]
- De Palma, M.; Mazzieri, R.; Politi, L.S.; Pucci, F.; Zonari, E.; Sitia, G.; Mazzoleni, S.; Moi, D.; Venneri, M.A.; Indraccolo, S. Tumor-targeted interferon-α delivery by Tie2-expressing monocytes inhibits tumor growth and metastasis. Cancer Cell 2008, 14, 299–311. [Google Scholar] [CrossRef] [Green Version]
- Däbritz, J. Granulocyte macrophage colony-stimulating factor and the intestinal innate immune cell homeostasis in Crohn’s disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G455–G465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsubota, Y.; Frey, J.M.; Raines, E.W. Technical Advance: Novel ex vivo culture method for human monocytes uses shear flow to prevent total loss of transendothelial diapedesis function. J. Leukoc. Biol. 2014, 95, 191–195. [Google Scholar] [CrossRef]
- Hamilton, J.A. Colony-stimulating factors in inflammation and autoimmunity. Nat Rev Immunol 2008, 8, 533–544. [Google Scholar] [CrossRef]
- Hamilton, J.A. GM-CSF in inflammation. J. Exp. Med. 2020, 217, e20190945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolcetti, L.; Peranzoni, E.; Ugel, S.; Marigo, I.; Fernandez Gomez, A.; Mesa, C.; Geilich, M.; Winkels, G.; Traggiai, E.; Casati, A. Hierarchy of immunosuppressive strength among myeloid-derived suppressor cell subsets is determined by GM-CSF. Eur. J. Immunol. 2010, 40, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Rößner, S.; Voigtländer, C.; Wiethe, C.; Hänig, J.; Seifarth, C.; Lutz, M.B. Myeloid dendritic cell precursors generated from bone marrow suppress T cell responses via cell contact and nitric oxide production in vitro. Eur. J. Immunol. 2005, 35, 3533–3544. [Google Scholar] [CrossRef] [PubMed]
- Kelley, J.L.; Rozek, M.M.; Suenram, C.A.; Schwartz, C.J. Activation of human blood monocytes by adherence to tissue culture plastic surfaces. Exp. Mol. Pathol. 1987, 46, 266–278. [Google Scholar] [CrossRef]
- Yamada, K.J.; Heim, C.E.; Xi, X.; Attri, K.S.; Wang, D.; Zhang, W.; Singh, P.K.; Bronich, T.K.; Kielian, T. Monocyte metabolic reprogramming promotes pro-inflammatory activity and Staphylococcus aureus biofilm clearance. PLoS Pathog. 2020, 16, e1008354. [Google Scholar] [CrossRef]
- Xie, L.; Dai, Z.; Pang, C.; Lin, D.; Zheng, M. Cellular glucose metabolism is essential for the reduction of cell-impermeable water-soluble tetrazolium (WST) dyes. Int. J. Biol. Sci. 2018, 14, 1535–1544. [Google Scholar] [CrossRef] [Green Version]
- Palmer, C.S.; Crowe, S.M. How does monocyte metabolism impact inflammation and aging during chronic HIV infection? AIDS Res. Hum. Retrovir. 2014, 30, 335–336. [Google Scholar] [CrossRef]
- Suzuki, H.; Hisamatsu, T.; Chiba, S.; Mori, K.; Kitazume, M.T.; Shimamura, K.; Nakamoto, N.; Matsuoka, K.; Ebinuma, H.; Naganuma, M.; et al. Glycolytic pathway affects differentiation of human monocytes to regulatory macrophages. Immunol. Lett. 2016, 176, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Groh, L.; Keating, S.T.; Joosten, L.A.B.; Netea, M.G.; Riksen, N.P. Monocyte and macrophage immunometabolism in atherosclerosis. Semin. Immunopathol. 2018, 40, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.; Agarwal, M.; Mukherjee, R.; Sen, P.; Sinha, D.K. 3D micro-environment regulates NF-κβ dependent adhesion to induce monocyte differentiation. Cell Death Dis. 2018, 9, 914. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, T.; Ley, K. Monocyte trafficking across the vessel wall. Cardiovasc Res 2015, 107, 321–330. [Google Scholar] [CrossRef] [Green Version]
- World, C.J.; Garin, G.; Berk, B.C. Vascular shear stress and activation of inflammatory genes. Curr. Atheroscler. Rep. 2006, 8, 240–244. [Google Scholar] [CrossRef]
- Phillipson, M.; Heit, B.; Parsons, S.A.; Petri, B.; Mullaly, S.C.; Colarusso, P.; Gower, R.M.; Neely, G.; Simon, S.I.; Kubes, P. Vav1 is essential for mechanotactic crawling and migration of neutrophils out of the inflamed microvasculature. J. Immunol. 2009, 182, 6870–6878. [Google Scholar] [CrossRef] [PubMed]
- Ong, S.-M.; Hadadi, E.; Dang, T.-M.; Yeap, W.-H.; Tan, C.T.-Y.; Ng, T.-P.; Larbi, A.; Wong, S.-C. The pro-inflammatory phenotype of the human non-classical monocyte subset is attributed to senescence. Cell Death Dis. 2018, 9, 266. [Google Scholar] [CrossRef]
- Ramos, T.N.; Bullard, D.C.; Barnum, S.R. ICAM-1: Isoforms and phenotypes. J. Immunol. 2014, 192, 4469–4474. [Google Scholar] [CrossRef] [Green Version]
- Möst, J.; Schwaeble, W.; Drach, J.; Sommerauer, A.; Dierich, M. Regulation of the expression of ICAM-1 on human monocytes and monocytic tumor cell lines. J. Immunol. 1992, 148, 1635–1642. [Google Scholar]
- Alexander, C.L.; Edward, M.; MacKie, R.M. The role of human melanoma cell ICAM-1 expression on lymphokine activated killer cell-mediated lysis, and the effect of retinoic acid. Br. J. Cancer 1999, 80, 1494–1500. [Google Scholar] [CrossRef] [Green Version]
- Marlin, S.D.; Springer, T.A. Purified intercellular adhesion molecule-1 (ICAM-1) is a ligand for lymphocyte function-associated antigen 1 (LFA-1). Cell 1987, 51, 813–819. [Google Scholar] [CrossRef]
- Klinder, A.; Markhoff, J.; Jonitz-Heincke, A.; Sterna, P.; Salamon, A.; Bader, R. Comparison of different cell culture plates for the enrichment of non-adherent human mononuclear cells. Exp. Ther. Med. 2019, 17, 2004–2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yakubenko, V.P.; Lishko, V.K.; Lam, S.C.-T.; Ugarova, T.P. A molecular basis for integrin αMβ2 ligand binding promiscuity. J. Biol. Chem. 2002, 277, 48635–48642. [Google Scholar] [CrossRef] [Green Version]
- Renkawitz, J.; Schumann, K.; Weber, M.; Lämmermann, T.; Pflicke, H.; Piel, M.; Polleux, J.; Spatz, J.P.; Sixt, M. Adaptive force transmission in amoeboid cell migration. Nat. Cell Biol. 2009, 11, 1438–1443. [Google Scholar] [CrossRef] [PubMed]
- Yakubenko, V.P.; Bhattacharjee, A.; Pluskota, E.; Cathcart, M.K. αMβ₂ integrin activation prevents alternative activation of human and murine macrophages and impedes foam cell formation. Circ. Res. 2011, 108, 544–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrchen, J.; Steinmüller, L.; Barczyk, K.; Tenbrock, K.; Nacken, W.; Eisenacher, M.; Nordhues, U.; Sorg, C.; Sunderkötter, C.; Roth, J. Glucocorticoids induce differentiation of a specifically activated, anti-inflammatory subtype of human monocytes. Blood 2006, 109, 1265–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wirthgen, E.; Hornschuh, M.; Wrobel, I.M.; Manteuffel, C.; Däbritz, J. Mimicking of Blood Flow Results in a Distinct Functional Phenotype in Human Non-Adherent Classical Monocytes. Biology 2021, 10, 748. https://doi.org/10.3390/biology10080748
Wirthgen E, Hornschuh M, Wrobel IM, Manteuffel C, Däbritz J. Mimicking of Blood Flow Results in a Distinct Functional Phenotype in Human Non-Adherent Classical Monocytes. Biology. 2021; 10(8):748. https://doi.org/10.3390/biology10080748
Chicago/Turabian StyleWirthgen, Elisa, Melanie Hornschuh, Ida Maria Wrobel, Christian Manteuffel, and Jan Däbritz. 2021. "Mimicking of Blood Flow Results in a Distinct Functional Phenotype in Human Non-Adherent Classical Monocytes" Biology 10, no. 8: 748. https://doi.org/10.3390/biology10080748
APA StyleWirthgen, E., Hornschuh, M., Wrobel, I. M., Manteuffel, C., & Däbritz, J. (2021). Mimicking of Blood Flow Results in a Distinct Functional Phenotype in Human Non-Adherent Classical Monocytes. Biology, 10(8), 748. https://doi.org/10.3390/biology10080748