
Unilateral Ureteral Obstruction for 28 Days in Rats Is Not Associated with Changes in Cardiac Function or Alterations in Mitochondrial Function

 , ,
, ,  , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animals and Ethical Guidelines
2.3. Antibodies
2.4. Experimental Design
2.5. Assessment of Tubular Damage and Fibrosis by Histological Analysis
2.6. Blood Serum Biochemistry
2.7. Evaluation of Cardiac Function by Echocardiography
2.8. Determination of Pro-Inflammatory Cytokines
2.9. Tissue Preparation
2.10. Cardiac Mitochondria Isolation
2.11. Mitochondrial Oxygen Consumption, Transmembranal Potential (∆Ψm), and ATP Synthase Activity
2.12. Activity of Antioxidant Enzymes in Mitochondria
2.13. Immunoblot Analysis
2.14. Statistical Analysis
3. Results
3.1. Development of the UUO Model
3.2. Cardiac Function
3.3. Proteins Related to Cardiac Damage and Inflammation
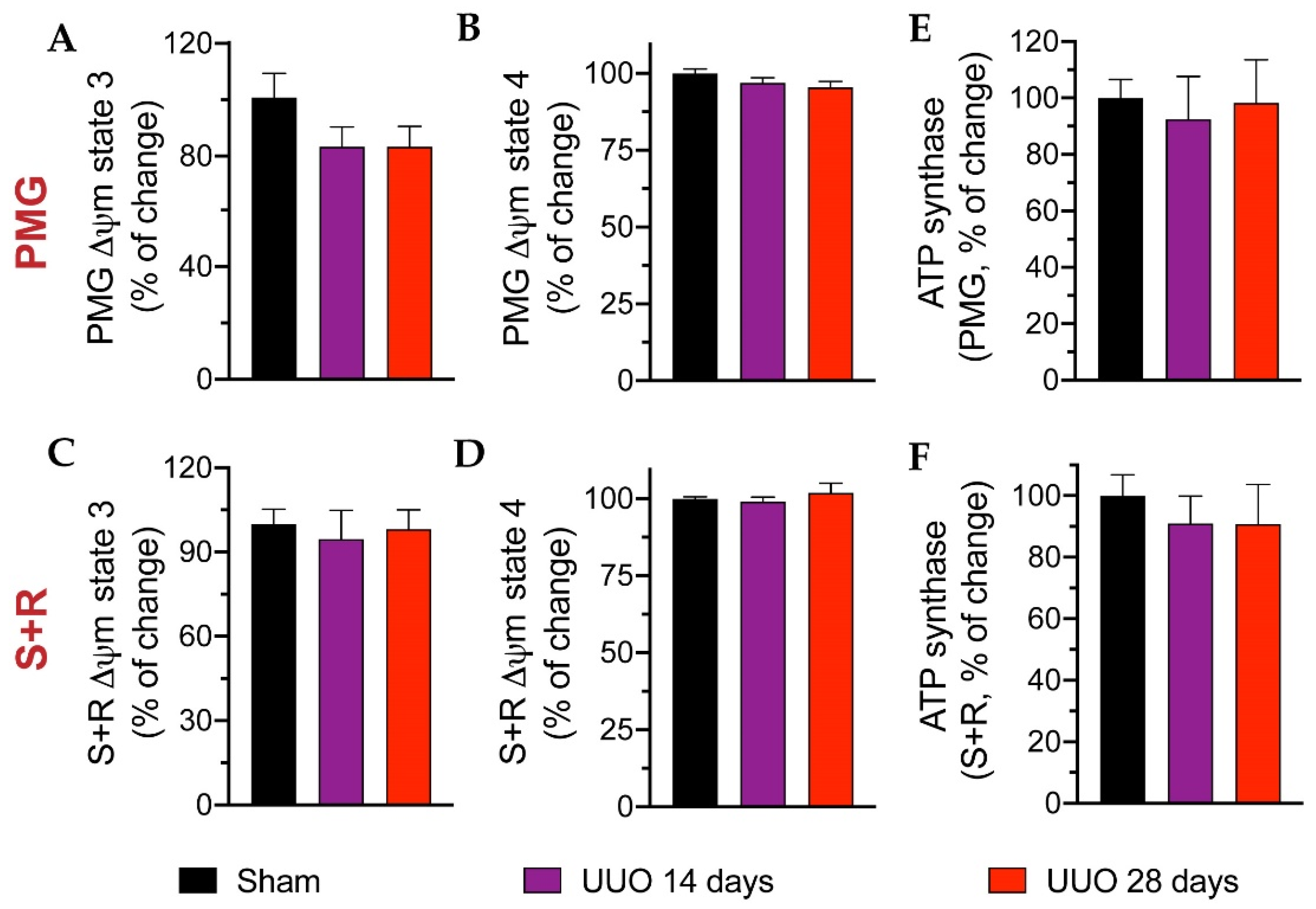
3.4. Mitochondrial Respiration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaeidi, A.; Sahamsizadeh, A.; Allahtavakoli, M.; Fatemi, I.; Rahmani, M.; Hakimizadeh, E.; Hassanshahi, J. The effect of oleuropein on unilateral ureteral obstruction induced-kidney injury in rats: The role of oxidative stress, inflammation and apoptosis. Mol. Biol. Rep. 2020, 47, 1371–1379. [Google Scholar] [CrossRef]
- Chevalier, R.L.; Forbes, M.S.; Thornhill, B.A. Ureteral obstruction as a model of renal interstitial fibrosis and obstructive nephropathy. Kidney Int. 2009, 75, 1145–1152. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Klimova, E.; Aparicio-Trejo, O.E.; Tapia, E.; Pedraza-Chaverri, J. Unilateral Ureteral Obstruction as a Model to Investigate Fibrosis-Attenuating Treatments. Biomolecules 2019, 9, 141. [Google Scholar] [CrossRef] [Green Version]
- Klahr, S.; Morrissey, J. Obstructive nephropathy and renal fibrosis. Am. J. Physiol. Renal Physiol. 2002, 283, F861–F875. [Google Scholar] [CrossRef] [Green Version]
- Manucha, W. Biochemical-molecular markers in unilateral ureteral obstruction. Biocell 2007, 31, 1–12. [Google Scholar] [CrossRef]
- Prud’homme, M.; Coutrot, M.; Michel, T.; Boutin, L.; Genest, M.; Poirier, F.; Launay, J.M.; Kane, B.; Kinugasa, S.; Prakoura, N.; et al. Acute Kidney Injury Induces Remote Cardiac Damage and Dysfunction Through the Galectin-3 Pathway. JACC Basic Transl. Sci. 2019, 4, 717–732. [Google Scholar] [CrossRef]
- Wen, Y.; Lu, X.; Ren, J.; Privratsky, J.R.; Yang, B.; Rudemiller, N.P.; Zhang, J.; Griffiths, R.; Jain, M.K.; Nedospasov, S.A.; et al. KLF4 in Macrophages Attenuates TNFα-Mediated Kidney Injury and Fibrosis. J. Am. Soc. Nephrol. 2019, 30, 1925–1938. [Google Scholar] [CrossRef]
- Bhargava, P.; Schnellmann, R.G. Mitochondrial energetics in the kidney. Nat. Rev. Nephrol. 2017, 13, 629–646. [Google Scholar] [CrossRef]
- Ham, O.; Jin, W.; Lei, L.; Huang, H.H.; Tsuji, K.; Huang, M.; Roh, J.; Rosenzweig, A.; Lu, H.A.J. Pathological cardiac remodeling occurs early in CKD mice from unilateral urinary obstruction, and is attenuated by Enalapril. Sci. Rep. 2018, 8, 16087. [Google Scholar] [CrossRef]
- Correa, F.; Buelna-Chontal, M.; Hernández-Reséndiz, S.; García-Niño, W.R.; Roldán, F.J.; Soto, V.; Silva-Palacios, A.; Amador, A.; Pedraza-Chaverrí, J.; Tapia, E.; et al. Curcumin maintains cardiac and mitochondrial function in chronic kidney disease. Free Radic. Biol. Med. 2013, 6, 119–129. [Google Scholar] [CrossRef]
- Rojas-Morales, P.; Tapia, E.; León-Contreras, J.C.; González-Reyes, S.; Jiménez-Osorio, A.S.; Trujillo, J.; Pavón, N.; Granados-Pineda, J.; Hernández-Pando, R.; Sánchez-Lozada, L.G.; et al. Mechanisms of Fasting-Mediated Protection against Renal Injury and Fibrosis Development after Ischemic Acute Kidney Injury. Biomolecules 2019, 22, 404. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Klimova, E.; Aparicio-Trejo, O.E.; Gómez-Sierra, T.; Jiménez-Uribe, A.P.; Bellido, B.; Pedraza-Chaverri, J. Mitochondrial dysfunction and endoplasmic reticulum stress in the promotion of fibrosis in obstructive nephropathy induced by unilateral ureteral obstruction. Biofactors 2020, 46, 716–733. [Google Scholar] [CrossRef]
- Bianco, M.; Lopes, J.A.; Beiral, H.J.V.; Filho, J.D.D.; Frankenfeld, S.P.; Fortunato, R.S.; Gattass, C.R.; Vieyra, A.; Takiya, C.M. The contralateral kidney presents with impaired mitochondrial functions and disrupted redox homeostasis after 14 days of unilateral ureteral obstruction in mice. PLoS ONE 2019, 14, e0218986. [Google Scholar] [CrossRef]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving bioscience research reporting: The ARRIVE guidelines for reporting animal research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef]
- Silva-Palacios, A.; Ostolga-Chavarría, M.; Buelna-Chontal, M.; Garibay, C.; Hernández-Reséndiz, S.; Roldán, F.J.; Flores, P.L.; Luna-López, A.; Königsberg, M.; Zazueta, C. 3-NP-induced Huntington’s-like disease impairs Nrf2 activation without loss of cardiac function in aged rats. Exp. Gerontol. 2017, 96, 89–98. [Google Scholar] [CrossRef]
- Pavón, N.; Aranda, A.; García, N.; Hernández-Esquivel, L.; Chávez, E. In hyperthyroid rats octylguanidine protects the heart from reperfusion damage. Endocrine 2009, 35, 158–165. [Google Scholar] [CrossRef]
- Aparicio-Trejo, O.E.; Avila-Rojas, S.H.; Tapia, E.; Rojas-Morales, P.; León-Contreras, J.C.; Martínez-Klimova, E.; Hernández-Pando, R.; Sánchez-Lozada, L.G.; Pedraza-Chaverri, J. Chronic impairment of mitochondrial bioenergetics and β-oxidation promotes experimental AKI-to-CKD transition induced by folic acid. Free Radic. Biol. Med. 2020, 154, 18–32. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farra, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Aparicio-Trejo, O.E.; Rojas-Morales, P.; Avila-Rojas, S.H.; León-Contreras, J.C.; Hernández-Pando, R.; Jiménez-Uribe, A.P.; Prieto-Carrasco, R.; Sánchez-Lozada, L.G.; Pedraza-Chaverri, J.; Tapia, E. Temporal Alterations in Mitochondrial β-Oxidation and Oxidative Stress Aggravate Chronic Kidney Disease Development in 5/6 Nephrectomy Induced Renal Damage. Int. J. Mol. Sci. 2020, 21, 6512. [Google Scholar] [CrossRef]
- Ojuka, E.; Andrew, B.; Bezuidenhout, N.; George, S.; Maarman, G.; Madlala, H.P.; Mendham, A.; Osiki, P.O. Measurement of β-oxidation capacity of biological samples by respirometry: A review of principles and substrates. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E715–E723. [Google Scholar] [CrossRef] [Green Version]
- Aparicio-Trejo, O.E.; Reyes-Fermín, L.M.; Briones-Herrera, A.; Tapia, E.; León-Contreras, J.C.; Hernández-Pando, R.; Sánchez-Lozada, L.G.; Pedraza-Chaverri, J. Protective effects of N-acetyl-cysteine in mitochondria bioenergetics, oxidative stress, dynamics and S-glutathionylation alterations in acute kidney damage induced by folic acid. Free Radic. Biol. Med. 2019, 130, 379–396. [Google Scholar] [CrossRef]
- Aparicio-Trejo, O.E.; Tapia, E.; Molina-Jijón, E.; Medina-Campos, O.N.; Macías-Ruvalcaba, N.A.; León-Contreras, J.C.; Hernández-Pando, R.; García-Arroyo, F.E.; Cristóbal, M.; Sánchez-Lozada, L.G.; et al. Curcumin prevents mitochondrial dynamics disturbances in early 5/6 nephrectomy: Relation to oxidative stress and mitochondrial bioenergetics. Biofactors 2017, 43, 293–310. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Gajjala, P.R.; Sanati, M.; Jankowski, J. Cellular and Molecular Mechanisms of Chronic Kidney Disease with Diabetes Mellitus and Cardiovascular Diseases as Its Comorbidities. Front. Immunol. 2015, 6, 340. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.W.; Su, C.T.; Song, E.J.; Tsai, W.C.; Li, Y.H.; Tsai, L.M.; Chen, J.H.; Sung, J.M. The role of echocardiographic study in patients with chronic kidney disease. J. Formos. Med. Assoc. 2015, 114, 797–805. [Google Scholar] [CrossRef] [Green Version]
- Mitsnefes, M.M. Cardiovascular disease in children with chronic kidney disease. J. Am. Soc. Nephrol. 2012, 23, 578–585. [Google Scholar] [CrossRef]
- Cai, Q.; Mukku, V.K.; Ahmad, M. Coronary artery disease in patients with chronic kidney disease: A clinical update. Curr. Cardiol. Rev. 2013, 9, 331–339. [Google Scholar] [CrossRef] [Green Version]
- Winterberg, P.D.; Jiang, R.; Maxwell, J.T.; Wang, B.; Wagner, M.B. Myocardial dysfunction occurs prior to changes in ventricular geometry in mice with chronic kidney disease (CKD). Physiol. Rep. 2016, 4, e12732. [Google Scholar] [CrossRef] [Green Version]
- McCullough, P.A.; Kellum, J.A.; Haase, M.; Müller, C.; Damman, K.; Murray, P.T.; Cruz, D.; House, A.A.; Schmidt-Ott, K.M.; Vescovo, G.; et al. Pathophysiology of the cardiorenal syndromes: Executive summary from the eleventh consensus conference of the Acute Dialysis Quality Initiative (ADQI). Contrib. Nephrol. 2013, 182, 82–98. [Google Scholar]
- Das, S.; Aiba, T.; Rosenberg, M.; Hessler, K.; Xiao, C.; Quintero, P.A.; Ottaviano, F.G.; Knight, A.C.; Graham, E.L.; Boström, P.; et al. Pathological role of serum- and glucocorticoid-regulated kinase 1 in adverse ventricular remodeling. Circulation 2012, 126, 2208–2219. [Google Scholar] [CrossRef]
- Bongartz, L.G.; Braam, B.; Gaillard, C.A.; Cramer, M.J.; Goldschmeding, R.; Verhaar, M.C.; Doevendans, P.A.; Joles, J.A. Target organ cross talk in cardiorenal syndrome: Animal models. Am. J. Physiol. Renal Physiol. 2012, 303, F1253–F1263. [Google Scholar] [CrossRef]
- Hewitson, T.D.; Holt, S.G.; Smith, E.R. Animal Models to Study Links between Cardiovascular Disease and Renal Failure and Their Relevance to Human Pathology. Front. Immunol. 2015, 6, 465. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Lu, Z.; Wang, F.; Jiang, Z.; Lu, L.; Miao, N.; Wang, N. Renalase attenuates hypertension, renal injury and cardiac remodelling in rats with subtotal nephrectomy. J. Cell. Mol. Med. 2016, 20, 1106–1117. [Google Scholar] [CrossRef] [Green Version]
- Husain-Syed, F.; McCullough, P.A.; Birk, H.W.; Renker, M.; Brocca, A.; Seeger, W.; Ronco, C. Cardio-Pulmonary-Renal Interactions: A Multidisciplinary Approach. J. Am. Coll. Cardiol. 2015, 65, 2433–2448. [Google Scholar] [CrossRef] [PubMed]
- Rangaswami, J.; Bhalla, V.; Blair, J.E.A.; Chang, T.I.; Costa, S.; Lentine, K.L.; Lerma, E.V.; Mezue, K.; Molitch, M.; Mullens, W.; et al. American Heart Association Council on the Kidney in Cardiovascular Disease and Council on Clinical Cardiology. Cardiorenal Syndrome: Classification, Pathophysiology, Diagnosis, and Treatment Strategies: A Scientific Statement From the American Heart Association. Circulation 2019, 139, e840–e878. [Google Scholar] [PubMed]
- Saada, A. The use of individual patient’s fibroblasts in the search for personalized treatment of nuclear encoded OXPHOS diseases. Mol. Genet. Metab. 2011, 104, 39–47. [Google Scholar] [CrossRef]
- Saada, A. Mitochondria: Mitochondrial OXPHOS (dys) function ex vivo–the use of primary fibroblasts. Int. J. Biochem. Cell. Biol. 2014, 48, 60–65. [Google Scholar] [CrossRef]
- Silva-Palacios, A.; Zazueta, C.; Pedraza-Chaverri, J. ER membranes associated with mitochondria: Possible therapeutic targets in heart-associated diseases. Pharmacol. Res. 2020. [Google Scholar] [CrossRef]
- Dominic, E.A.; Ramezani, A.; Anker, S.D.; Verma, M.; Mehta, N.; Rao, M. Mitochondrial cytopathies and cardiovascular disease. Heart 2014, 100, 611–618. [Google Scholar] [CrossRef]
- Ide, T.; Tsutsui, H.; Hayashidani, S.; Kang, D.; Suematsu, N.; Nakamura, K.; Utsumi, H.; Hamasaki, N.; Takeshita, A. Mitochondrial DNA damage and dysfunction associated with oxidative stress in failing hearts after myocardial infarction. Circ. Res. 2001, 88, 529–535. [Google Scholar] [CrossRef] [Green Version]
- Disatnik, M.H.; Ferreira, J.C.; Campos, J.C.; Gomes, K.S.; Dourado, P.M.; Qi, X.; Mochly-Rosen, D. Acute inhibition of excessive mitochondrial fission after myocardial infarction prevents long-term cardiac dysfunction. J. Am. Heart Assoc. 2013, 2, e000461. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UUO (Days) | |||
|---|---|---|---|
| Parameter | Sham | 14 | 28 |
| IVS (mm) | 0.20 ± 0.003 | 0.20 ± 0.002 | 0.198 ± 0.002 |
| LVEDd (mm) | 5.37 ± 0.07 | 5.25 ± 0.05 | 5.46 ± 0.05 |
| LVEDs (mm) | 2.46 ± 0.04 | 2.34 ± 0.05 | 2.28 ± 0.06 |
| LVPW (mm) | 0.19 ± 0.003 | 0.18 ± 0.002 | 0.19 ± 0.003 |
| EF (%) | 82.1 ± 0.44 | 83.7 ± 0.5 | 85.1 ± 0.6 |
| FS (%) | 54.22 ± 0.57 | 55.7 ± 0.7 | 58.5 ± 0.9 |
| HR (bpm) | 413.79 ± 6.7 | 433.1 ± 6.7 | 417.0 ± 7.9 |
| Body weight (BW, g) | 373.3 ± 4.2 | 350.7 ± 4.7 | 360.0 ± 3.6 |
| Heart weight (HW, g) | 1.41 ± 0.05 | 1.19 ± 0.02 | 1.31 ± 0.03 |
| Lung weight (LW, g) | 2.21 ± 0.08 | 1.92 ± 0.03 | 1.94 ± 0.05 |
| Tibial length (TL, cm) | 5.3 ± 0.03 | 5.3 ± 0.02 | 5.4 ± 0.02 |
| HW/TL (g/cm) | 0.25 ± 0.01 | 0.22 ± 0.005 | 0.24 ± 0.01 |
| LW/TL (g/cm) | 0.36 ± 0.01 | 0.36 ± 0.006 | 0.36 ± 0.01 |
| HW/BW (g/kg) | 3.57 ± 0.11 | 3.40 ± 0.06 | 3.64 ± 0.08 |
| LW/BW (g/kg) | 5.20 ± 0.20 | 5.54 ± 0.12 | 5.38 ± 0.12 |
| Functional parameters | (n = 9) | (n = 10) | (n = 10) |
| UUO (Days) | |||
|---|---|---|---|
| Sham | 14 | 28 | |
| IL-1 (pg/mg) | 10.56 ± 0.5 | 10.67 ± 0.88 | 21.14 ± 1.9 a,b |
| IL-6 (pg/mg) | 27.73 ± 10.76 | 11.98 ± 1.15 | 36.66 ± 13.27 |
| TNF-α (pg/mg) | 9.41 ± 0.41 | 8.77 ± 1.14 | 14.79 ± 2.16 c |
| BUN (mg/dL) | 21.67 ± 0.33 | 23.25 ± 2.05 | 28.50 ± 5.18 |
| Creatinine (mg/dL) | 0.49 ± 0.009 | 0.61 ± 0.02 | 0.65 ± 0.03 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prieto-Carrasco, R.; Silva-Palacios, A.; Rojas-Morales, P.; Aparicio-Trejo, O.E.; Medina-Reyes, E.I.; Hernández-Cruz, E.Y.; Sánchez-Garibay, C.; Salinas-Lara, C.; Pavón, N.; Roldán, F.J.; et al. Unilateral Ureteral Obstruction for 28 Days in Rats Is Not Associated with Changes in Cardiac Function or Alterations in Mitochondrial Function. Biology 2021, 10, 671. https://doi.org/10.3390/biology10070671
Prieto-Carrasco R, Silva-Palacios A, Rojas-Morales P, Aparicio-Trejo OE, Medina-Reyes EI, Hernández-Cruz EY, Sánchez-Garibay C, Salinas-Lara C, Pavón N, Roldán FJ, et al. Unilateral Ureteral Obstruction for 28 Days in Rats Is Not Associated with Changes in Cardiac Function or Alterations in Mitochondrial Function. Biology. 2021; 10(7):671. https://doi.org/10.3390/biology10070671
Chicago/Turabian StylePrieto-Carrasco, Rodrigo, Alejandro Silva-Palacios, Pedro Rojas-Morales, Omar Emiliano Aparicio-Trejo, Estefany Ingrid Medina-Reyes, Estefani Yaquelin Hernández-Cruz, Carlos Sánchez-Garibay, Citlaltepetl Salinas-Lara, Natalia Pavón, Francisco Javier Roldán, and et al. 2021. "Unilateral Ureteral Obstruction for 28 Days in Rats Is Not Associated with Changes in Cardiac Function or Alterations in Mitochondrial Function" Biology 10, no. 7: 671. https://doi.org/10.3390/biology10070671
APA StylePrieto-Carrasco, R., Silva-Palacios, A., Rojas-Morales, P., Aparicio-Trejo, O. E., Medina-Reyes, E. I., Hernández-Cruz, E. Y., Sánchez-Garibay, C., Salinas-Lara, C., Pavón, N., Roldán, F. J., Zazueta, C., Tapia, E., & Pedraza-Chaverri, J. (2021). Unilateral Ureteral Obstruction for 28 Days in Rats Is Not Associated with Changes in Cardiac Function or Alterations in Mitochondrial Function. Biology, 10(7), 671. https://doi.org/10.3390/biology10070671