
Discovery of Novel GMPS Inhibitors of Candidatus Liberibacter Asiaticus by Structure Based Design and Enzyme Kinetic



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Homology Modeling and Structure Validation
2.2. Virtual Screening
2.3. Molecule Docking Study
2.4. Absorption, Distribution, Metabolism, and Excretion and Toxicity Prediction
2.5. Gene Cloning
2.6. Protein Expression and Purification of CLas GMPS
2.7. Enzyme Assays and Kinetics
2.8. Inhibition Assay against CLas GMPS
3. Results
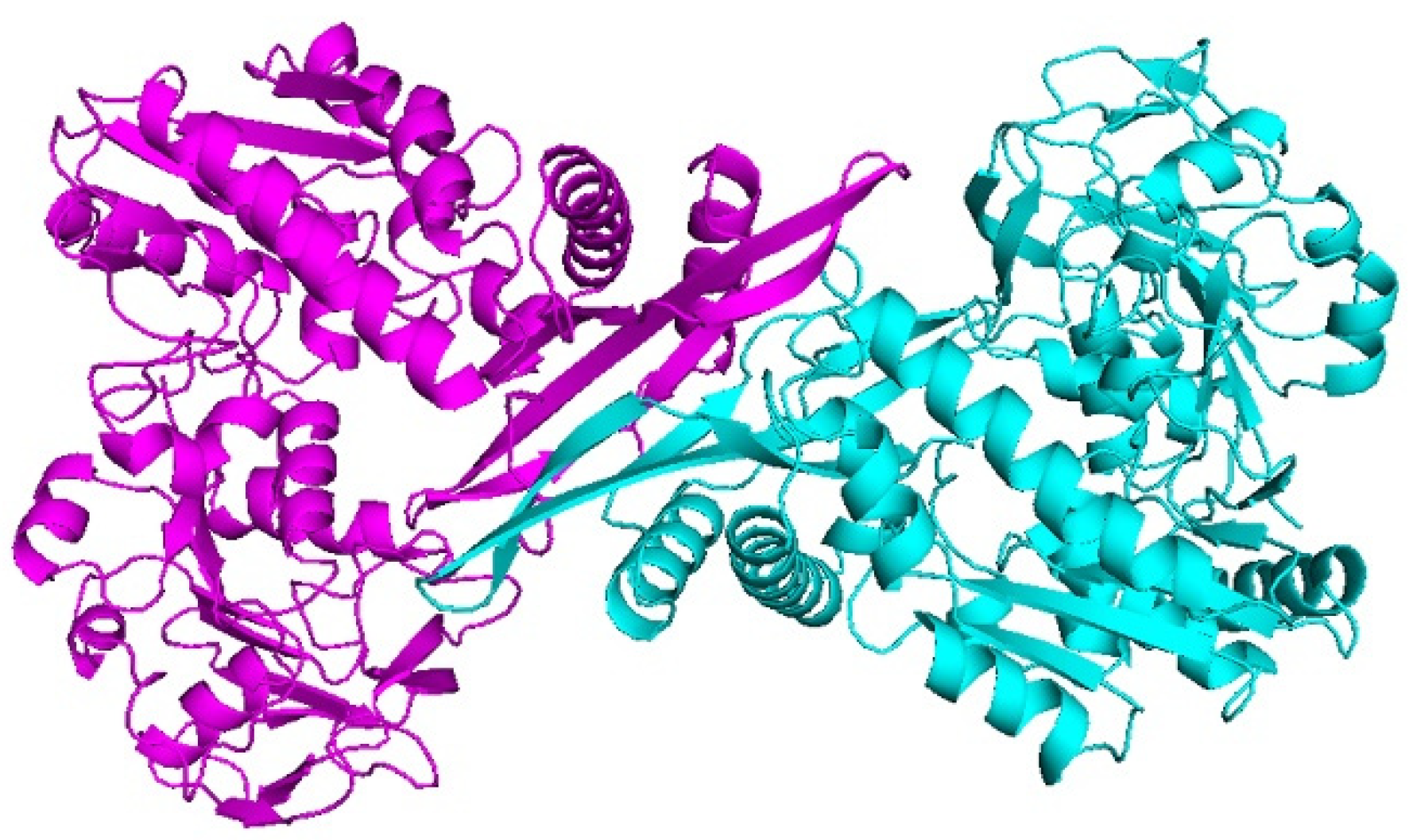
3.1. Model Building and Structure Validation
3.2. Virtual Screening
3.3. Molecular Docking
3.4. ADME and Toxicity Prediction
3.5. Purification of CLas GMPS Protein
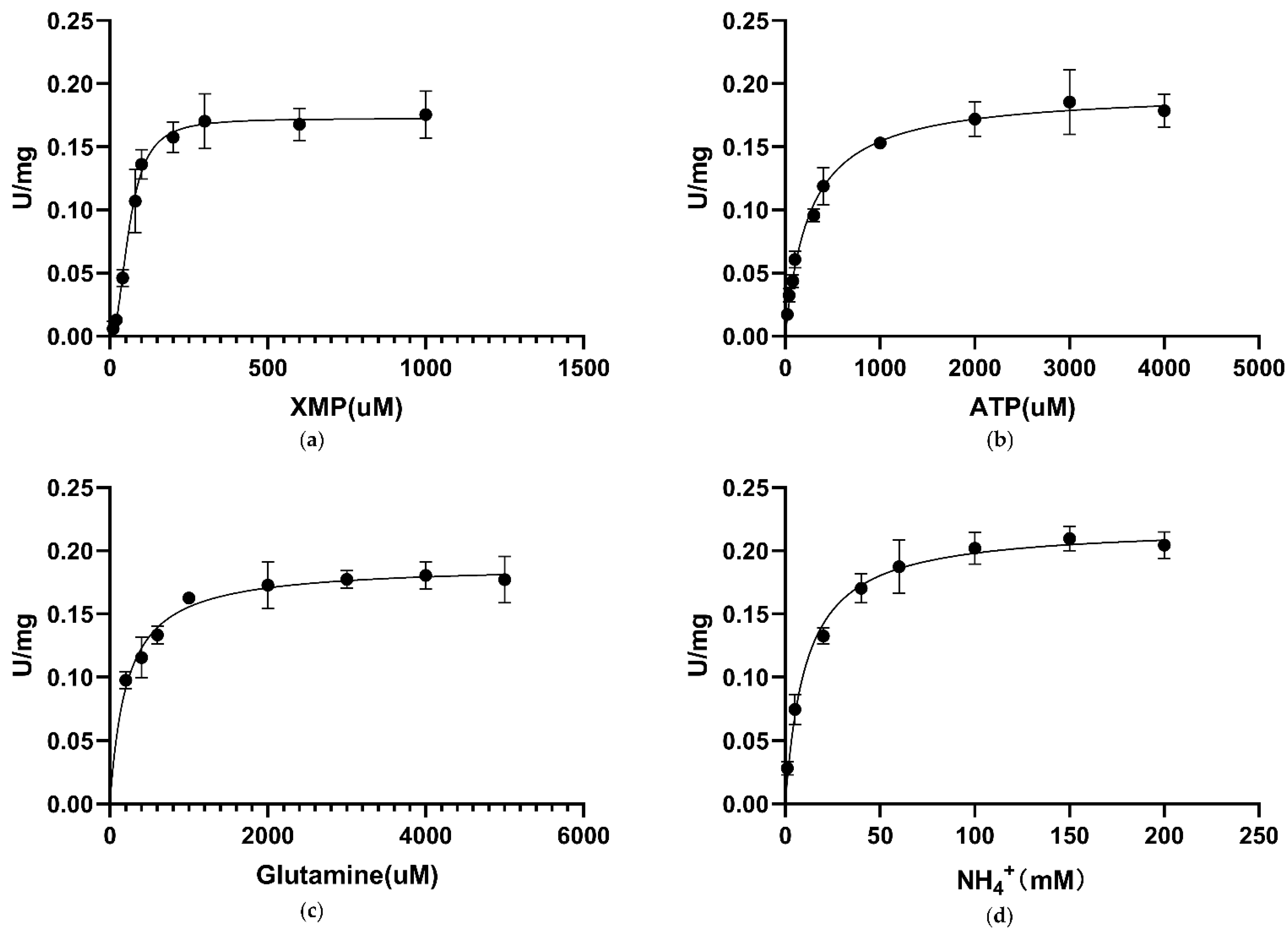
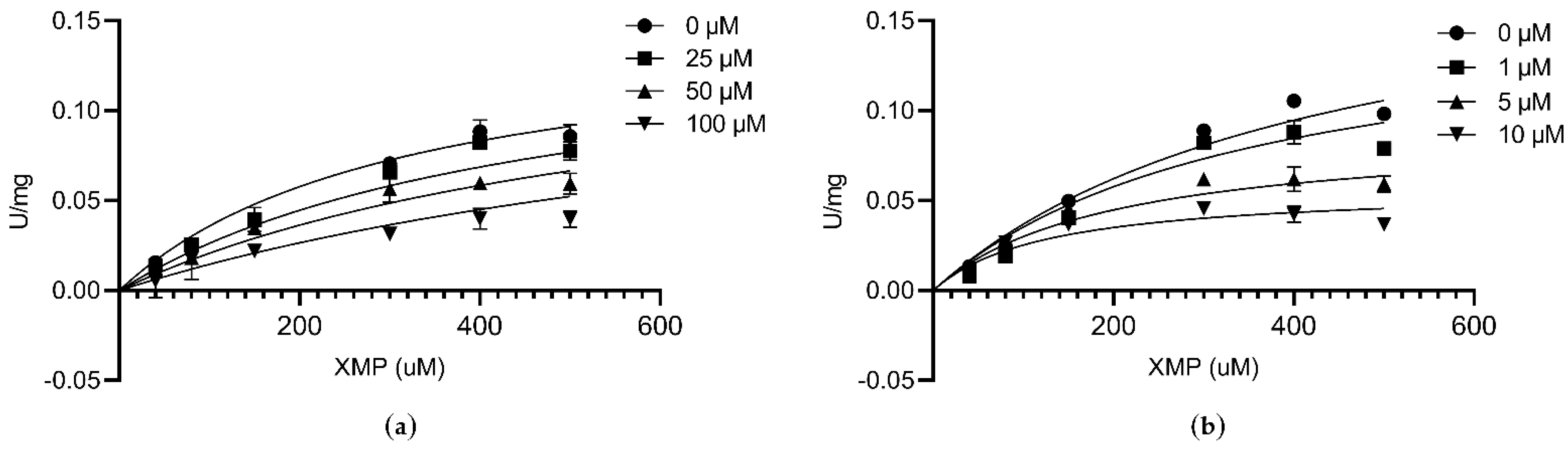
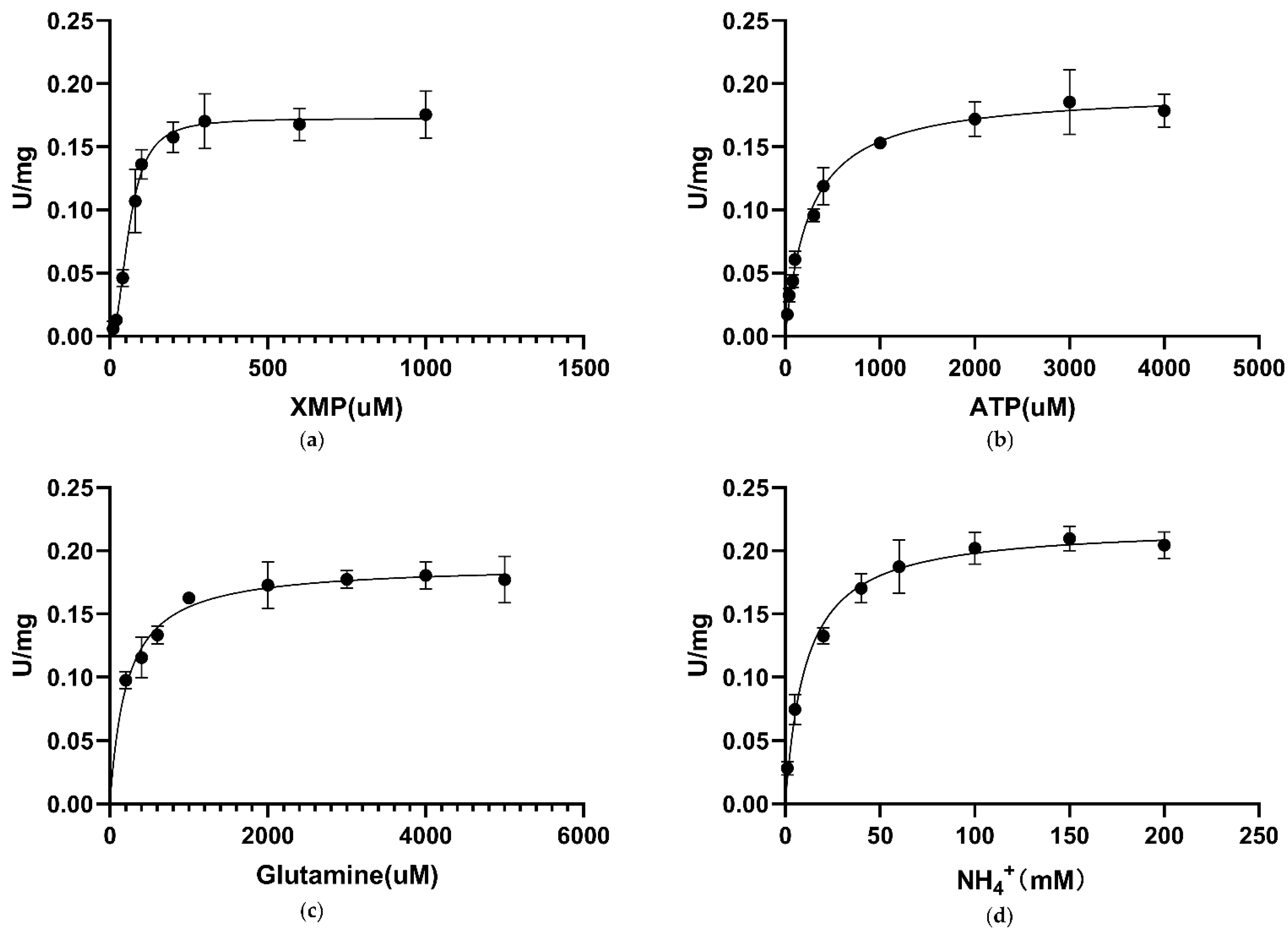
3.6. Kinetic Characterization of CLas GMPS
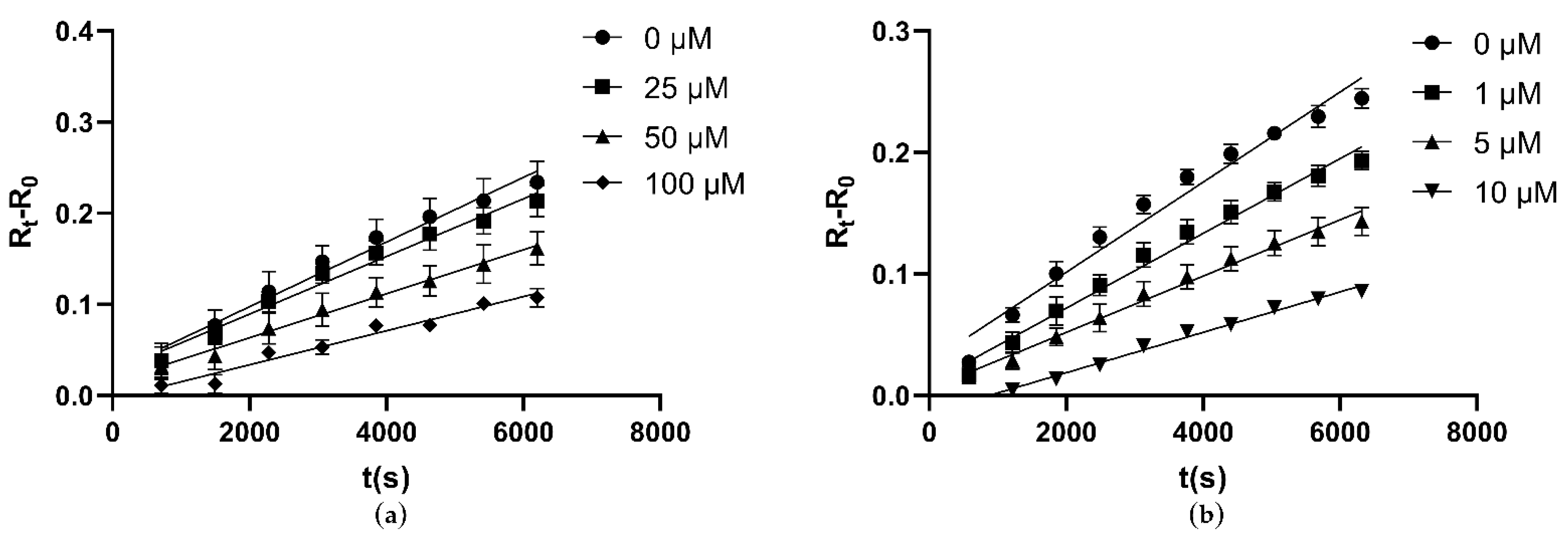
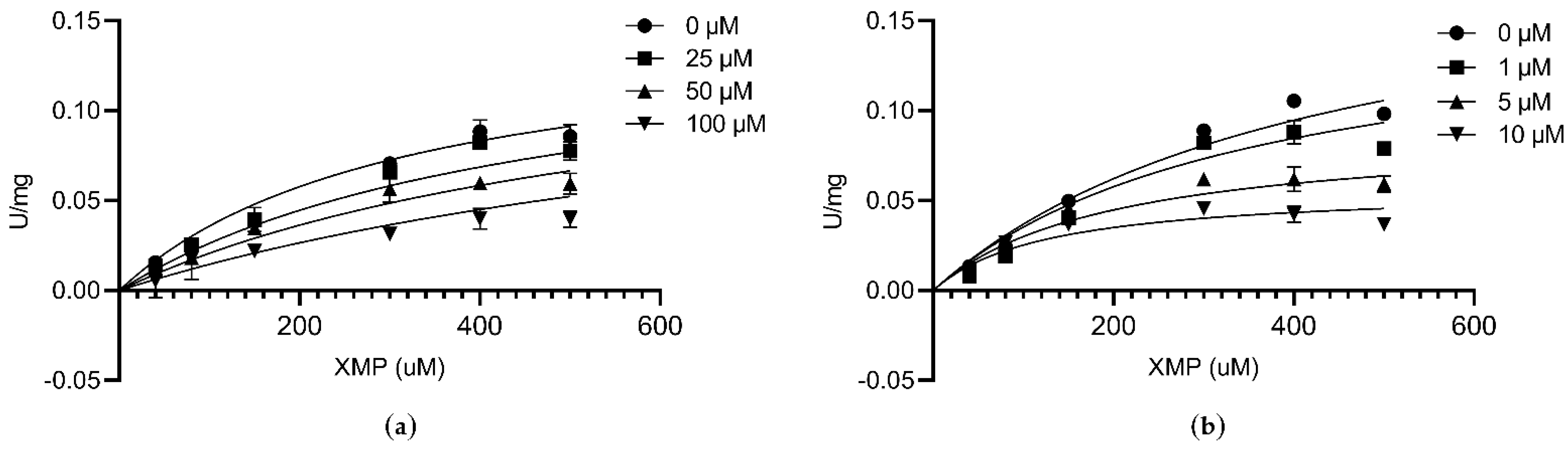
3.7. Inhibitory Assay against CLas GMPS Anzyme Activity
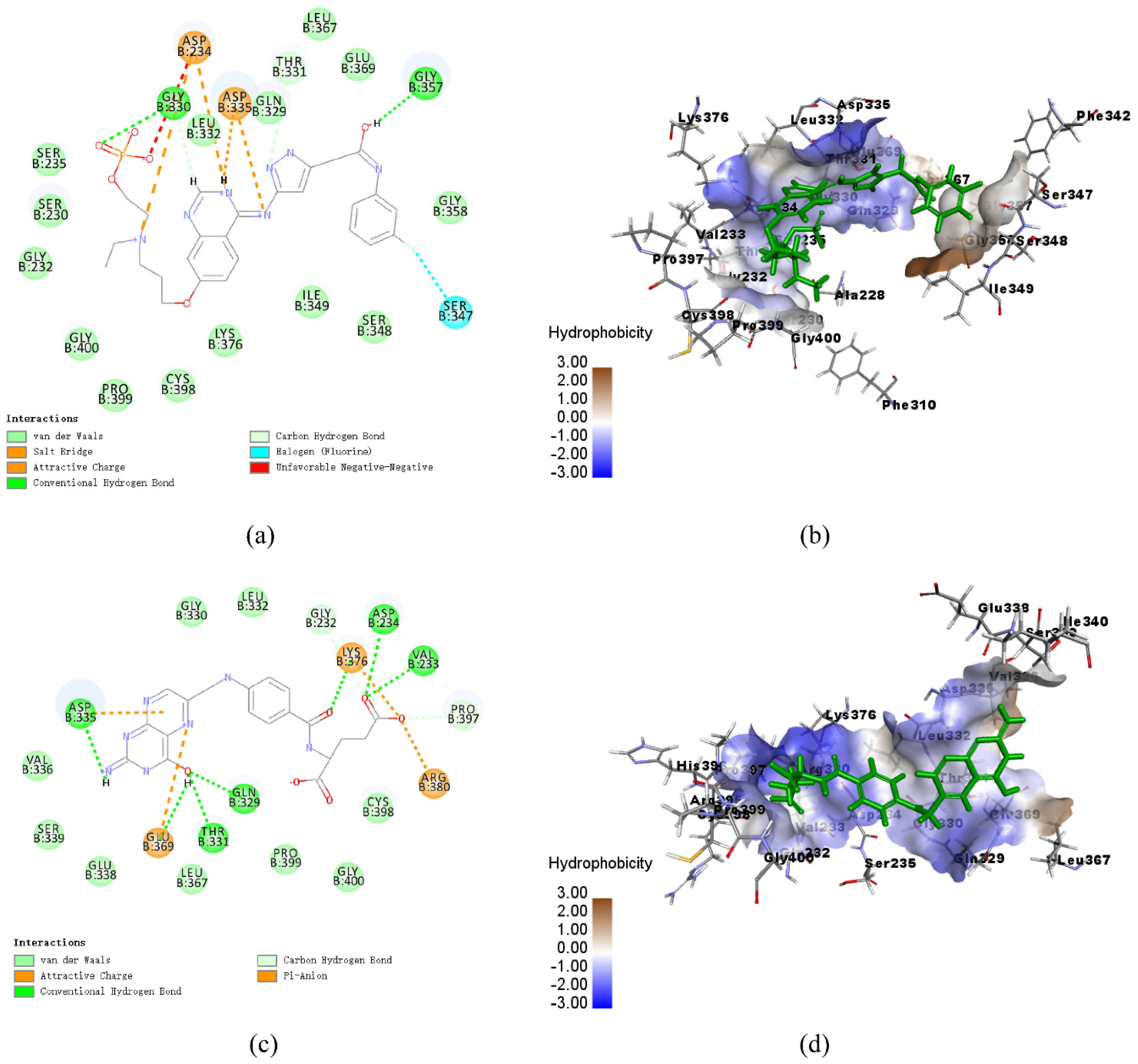
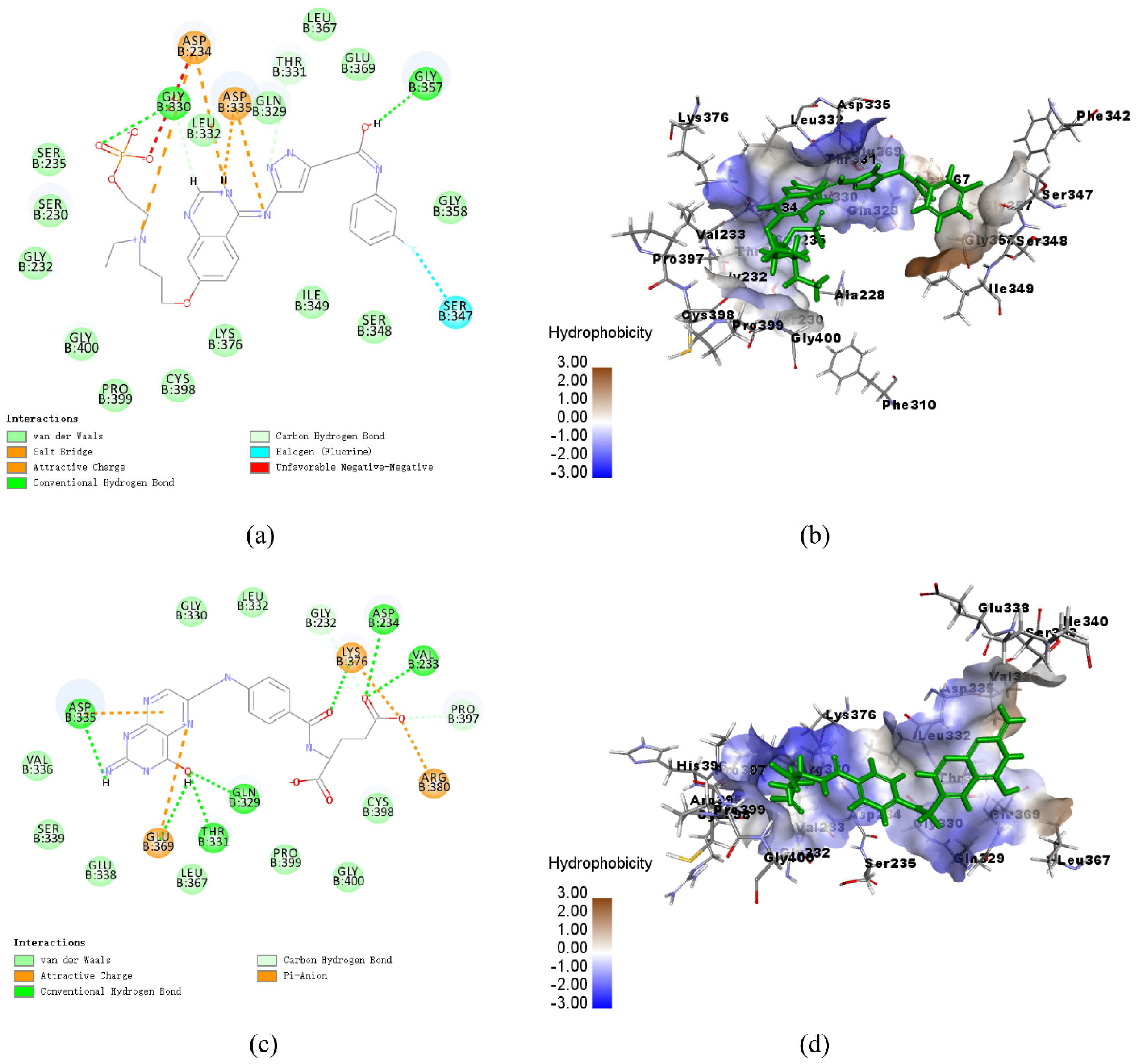
3.8. Analysis of Ligand Binding
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, N.; Trivedi, P. Citrus huanglongbing: A newly relevant disease presents unprecedented challenges. Phytopathology 2013, 103, 652–665. [Google Scholar] [CrossRef] [Green Version]
- da Graca, J.V.; Douhan, G.W.; Halbert, S.E.; Keremane, M.L.; Lee, R.F.; Vidalakis, G.; Zhao, H. Huanglongbing: An overview of a complex pathosystem ravaging the world’s citrus. J. Integr. Plant Biol. 2016, 58, 373–387. [Google Scholar] [CrossRef]
- Wang, N.; Pierson, E.A.; Setubal, J.C.; Xu, J.; Levy, J.G.; Zhang, Y.; Li, J.; Rangel, L.T.; Martins, J., Jr. The Candidatus Liberibacter-host interface: Insights into pathogenesis mechanisms and disease control. Annu. Rev. Phytopathol. 2017, 55, 451–482. [Google Scholar] [CrossRef]
- Bove, J.M. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar]
- Gottwald, T.R.; Graça, J.V.; Bassanezi, R.B. Citrus huanglongbing: The pathogen and its impact. Plant Health Prog. 2007, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Wang, N. The citrus huanglongbing crisis and potential solutions. Mol. Plant 2019, 12, 607–609. [Google Scholar] [CrossRef]
- Lin, K.H. Observations on yellow shoot on citrus. Etiological studies of yellow shoot of citrus. Acta Phytopathol. Sin. 1956, 2, 13–42. [Google Scholar]
- Bassanezi, R.B.; Montesino, L.H.; Gimenes-Fernandes, N.; Yamamoto, P.T.; Gottwald, T.R.; Amorim, L.; Bergamin Filho, A. Efficacy of area-wide inoculum reduction and vector control on temporal progress of huanglongbing in young sweet orange plantings. Plant Dis. 2013, 97, 789–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Powell, C.A.; Zhou, L.; He, Z.; Stover, E.; Duan, Y. Chemical compounds effective against the citrus huanglongbing bacterium ‘Candidatus Liberibacter asiaticus’ in planta. Phytopathology 2011, 101, 1097–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Rimawi, F.; Hijaz, F.; Nehela, Y.; Batuman, O.; Killiny, N. Uptake, translocation, and stability of oxytetracycline and streptomycin in citrus plants. Antibiotics 2019, 8, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Killiny, N.; Hijaz, F.; Al-Rimawi, F.; Batuman, O. Translocation of oxytetracycline in citrus plants after root drench and stem delivery. Proc. Fla. State Hort. Soc. 2019, 132, 68–71. [Google Scholar]
- Shin, K.; Ascunce, M.S.; Narouei-Khandan, H.A.; Sun, X.; Jones, D.; Kolawole, O.O.; Goss, E.M.; van Bruggen, A.H.C. Effects and side effects of penicillin injection in huanglongbing affected grapefruit trees. Crop Protect. 2016, 90, 106–116. [Google Scholar] [CrossRef]
- Zhang, M.; Karuppaiya, P.; Zheng, D.; Sun, X.; Bai, J.; Ferrarezi, R.S.; Powell, C.A.; Duan, Y. Field evaluation of chemotherapy on HLB-affected citrus trees with emphasis on fruit yield and quality. Front. Plant Sci. 2021, 12, 611287. [Google Scholar] [CrossRef]
- Barnett, M.J.; Solow-Cordero, D.E.; Long, S.R. A high-throughput system to identify inhibitors of Candidatus Liberibacter asiaticus transcription regulators. Proc. Natl. Acad. Sci. USA 2019, 116, 18009–18014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nan, J.; Zhang, S.; Zhan, P.; Jiang, L. Evaluation of bronopol and disulfiram as potential Candidatus Liberibacter asiaticus inosine 5′-monophosphate dehydrogenase inhibitors by using molecular docking and enzyme kinetic. Molecules 2020, 25, 2313. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-Y.; Araujo, K.; Sanchez, J.N.; Kund, G.; Trumble, J.; Roper, C.; Godfrey, K.E.; Jin, H. A stable antimicrobial peptide with dual functions of treating and preventing citrus Huanglongbing. Proc. Natl. Acad. Sci. USA 2021, 118, e2019628118. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Trivedi, P.; Wang, N. Field evaluation of plant defense inducers for the control of citrus Huanglongbing. Phytopathology 2016, 106, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Jiang, J.; Wang, N. Control of citrus huanglongbing via trunk injection of plant defense activators and antibiotics. Phytopathology 2018, 108, 186–195. [Google Scholar] [CrossRef] [Green Version]
- Nan, J.; Zhang, S.; Jiang, L. Antibacterial potential of Bacillus amyloliquefaciens GJ1 against citrus huanglongbing. Plants 2021, 10, 261. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, C.; Powell, C.A.; Avery, P.B.; Wang, J.; Huang, Y.; Duan, Y. Field evaluation of integrated management for mitigating citrus huanglongbing in Florida. Front. Plant. Sci. 2019, 9, 1890. [Google Scholar] [CrossRef] [Green Version]
- Cheviet, T.; Lefebvre-Tournier, I.; Wein, S.; Peyrottes, S. Plasmodium purine metabolism and its inhibition by nucleoside and nucleotide analogues. J. Med. Chem. 2019, 62, 8365–8391. [Google Scholar] [CrossRef] [Green Version]
- McConkey, G.A. Plasmodium falciparum: Isolation and characterisation of a gene encoding protozoan GMP synthase. Exp. Parasitol. 2000, 94, 23–32. [Google Scholar] [CrossRef]
- Massiere, F.; Badet-Denisot, M.A. The mechanism of glutamine-dependent amidotransferases. Cell. Mol. Life Sci. 1998, 54, 205–222. [Google Scholar] [CrossRef]
- Long, H.; Cameron, S.; Yu, L.; Rao, Y. De novo GMP synthesis is required for axon guidance in Drosophila. Genetics 2006, 172, 1633–1642. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Leija, C.; Rijo-Ferreira, F.; Chen, J.; Cestari, I.; Stuart, K.; Tu, B.P.; Phillips, M.A. GMP synthase is essential for viability and infectivity of Trypanosoma brucei despite a redundant purine salvage pathway. Mol. Microbiol. 2015, 97, 1006–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalkin, H.; Truitt, C.D. Characteriztion of glutamine site of Escherichia-coli guanosine 5′-monophosphate synthetase. J. Biol. Chem. 1977, 252, 5431–5436. [Google Scholar] [CrossRef]
- Chittur, S.V.; Klem, T.J.; Shafer, C.M.; Davisson, V.J. Mechanism for acivicin inactivation of triad glutamine amidotransferases. Biochemistry 2001, 40, 876–887. [Google Scholar] [CrossRef]
- Rodriguez-Suarez, R.; Xu, D.; Veillette, K.; Davison, J.; Sillaots, S.; Kauffman, S.; Hu, W.; Bowman, J.; Martel, N.; Trosok, S.; et al. Mechanism-of-action determination of GMP synthase inhibitors and target validation in Candida albicans and Aspergillus fumigatus. Chem. Biol. 2007, 14, 1163–1175. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, J.; Lou, L. Biochemical-characterization of human GMP synthetase. J. Biol. Chem. 1995, 270, 7347–7353. [Google Scholar] [CrossRef] [Green Version]
- Bhat, J.Y.; Shastri, B.G.; Balaram, H. Kinetic and biochemical characterization of Plasmodium falciparum GMP synthetase. Biochem. J. 2008, 409, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, D.; Luthy, R.; Bowie, J.U. VERIFY3D: Assessment of protein models with three-dimensional profiles. Methods Enzymol. 1997, 277, 396–404. [Google Scholar]
- Laskowski, R.A.; Macarthur, M.W.; Moss, D.S.; Thornton, J.M. Procheck—A Program to Check the Stereochemical Quality of Protein Structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Diller, D.J.; Merz, K.M., Jr. High throughput docking for library design and library prioritization. Proteins Struct. Funct. Genet. 2001, 43, 113–124. [Google Scholar] [CrossRef]
- Wu, G.; Robertson, D.H.; Brooks, C.L.; Vieth, M. Detailed analysis of grid-based molecular docking: A case study of CDOCKER-A CHARMm-based MD docking algorithm. J. Comput. Chem. 2003, 24, 1549–1562. [Google Scholar] [CrossRef]
- Franco, T.M.A.; Rostirolla, D.C.; Ducati, R.G.; Lorenzini, D.M.; Basso, L.A.; Santos, D.S. Biochemical characterization of recombinant guaA-encoded guanosine monophosphate synthetase (EC 6.3.5.2) from Mycobacterium tuberculosis H37Rv strain. Arch. Biochem. Biophys. 2012, 517, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, N.; Hatfield, G.W.; Moyed, H.S. Physical properties and subunit structure of xanthosine 5′-phosphate aminase. J. Biol. Chem. 1972, 247, 5880–5887. [Google Scholar] [CrossRef]
- Kumar, V.; Kancharla, S.; Jena, M.K. In silico virtual screening-based study of nutraceuticals predicts the therapeutic potentials of folic acid and its derivatives against COVID-19. VirusDisease 2021, 32, 1–9. [Google Scholar] [CrossRef]
- Oser, M.G.; Fonseca, R.; Chakraborty, A.A.; Brough, R.; Spektor, A.; Jennings, R.B.; Flaifel, A.; Novak, J.S.; Gulati, A.; Buss, E.; et al. Cells lacking the RB1 tumor suppressor gene are hyperdependent on aurora B kinase for survival. Cancer Discov. 2019, 9, 230–247. [Google Scholar] [CrossRef] [Green Version]
- Ballut, L.; Violot, S.; Shivakumaraswamy, S.; Thota, L.P.; Sathya, M.; Kunala, J.; Dijkstra, B.W.; Terreux, R.; Haser, R.; Balaram, H.; et al. Active site coupling in Plasmodium falciparum GMP synthetase is triggered by domain rotation. Nat. Commun. 2015, 6, 8930. [Google Scholar] [CrossRef] [Green Version]
- Kirubakaran, S.; Gorla, S.K.; Sharling, L.; Zhang, M.; Liu, X.; Ray, S.S.; MacPherson, I.S.; Striepen, B.; Hedstrom, L.; Cuny, G.D. Structure-activity relationship study of selective benzimidazole-based inhibitors of Cryptosporidium parvum IMPDH. Bioorganic Med. Chem. Lett. 2012, 22, 1985–1988. [Google Scholar] [CrossRef] [Green Version]
- Makowska-Grzyska, M.; Kim, Y.; Maltseva, N.; Osipiuk, J.; Gu, M.; Zhang, M.; Mandapati, K.; Gollapalli, D.R.; Gorla, S.K.; Hedstrom, L.; et al. A novel cofactor-binding mode in bacterial IMP dehydrogenases explains inhibitor selectivity. J. Biol. Chem. 2015, 290, 5893–5911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freedman, R.; Yu, R.; Sarkis, A.W.; Hedstrom, L. A structural determinant of mycophenolic acid resistance in eukaryotic inosine 50-monophosphate dehydrogenases. Protein Sci. Publ. Protein Soc. 2020, 29, 686–694. [Google Scholar] [CrossRef]
- Abbott, J.L.; Newell, J.M.; Lightcap, C.M.; Olanich, M.E.; Loughlin, D.T.; Weller, M.A.; Lam, G.; Pollack, S.; Patton, W.A. The effects of removing the GAT domain from E-coli GMP synthetase. Protein J. 2006, 25, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Maruoka, S.; Horita, S.; Lee, W.C.; Nagata, K.; Tanokura, M. Crystal structure of the ATPPase subunit and its substrate-dependent association with the GATase subunit: A novel regulatory mechanism for a two-subunit-type GMP synthetase from Pyrococcus horikoshii OT3. J. Mol. Biol. 2010, 395, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Tesmer, J.G.; Klem, T.J.; Deras, M.L.; Davisson, V.J.; Smith, J.L. The crystal structure of GMP synthetase reveals a novel catalytic triad and is a structural paradigm for two enzyme families. Nat. Struct. Biol. 1996, 3, 74–86. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Name | CAS Number | Molecular Weight | LibDock Score |
|---|---|---|---|---|
| 1 | Compound1334 | 722543-31-9 | 587.54 | 191.661 |
| 2 | Compound3170 | 1004316-88-4 | 776.02 | 183.169 |
| 3 | Compound8511 | 81624-55-7 | 636.79 | 182.643 |
| 4 | Compound3997 | 155213-67-5 | 720.9 | 176.273 |
| 5 | Compound5520 | 84366-81-4 | 829.51 | 174.471 |
| 6 | Compound1226 | 612847-09-3 | 551.64 | 174.129 |
| 7 | Compound3975 | 852808-04-9 | 813.43 | 171.437 |
| 8 | Compound2704 | 722544-51-6 | 507.56 | 167.324 |
| 9 | Compound8477 | 1235034-55-5 | 669.79 | 166.047 |
| 10 | Compound7726 | 1439399-58-2 | 571.57 | 165.201 |
| 11 | Compound2503 | 755038-02-9 | 521.67 | 165.127 |
| 12 | Compound3752 | 943319-70-8 | 532.56 | 164.791 |
| 13 | Compound4073 | 641571-10-0 | 529.53 | 163.076 |
| 14 | Compound1591 | 923288-90-8 | 583.99 | 163.076 |
| 15 | Compound6234 | 1062159-35-6 | 494.59 | 162.396 |
| 16 | Compound5313 | 887650-05-7 | 576.62 | 162.366 |
| 17 | Compound4979 | 1633044-56-0 | 634.73 | 162.348 |
| 18 | Compound8081 | 1108743-60-7 | 560.64 | 162.072 |
| 19 | Compound5162 | 7085-55-4 | 742.68 | 161.335 |
| 20 | Compound1446 | 356057-34-6 | 666.77 | 160.757 |
| Number | Name | Cdocker Energy (kcal/mol) |
|---|---|---|
| 1 | Compound2265 | −90.9198 |
| 2 | Compound531 | −89.2316 |
| 3 | Compound529 | −83.4206 |
| 4 | Compound532 | −81.3088 |
| 5 | Compound1334 | −76.7696 |
| 6 | Compound5292 | −76.5687 |
| 7 | Compound4759 | −75.9609 |
| 8 | Compound2293 | −70.4343 |
| 9 | Compound4419 | −67.4157 |
| 10 | Compound8826 | −66.4995 |
| 11 | Compound1926 | −65.2249 |
| 12 | Compound5481 | −53.8011 |
| 13 | Compound2295 | −51.375 |
| 14 | Compound5520 | −50.2805 |
| 15 | Compound2476 | −49.5968 |
| 16 | Compound8511 | −44.8984 |
| 17 | Compound2240 | −42.2788 |
| 18 | Compound7726 | −42.1659 |
| 19 | Compound2101 | −41.3667 |
| 20 | Compound4965 | −40.6106 |
| 21 | Compound8477 | −38.8839 |
| 22 | DON | −38.6205 |
| Number | Name | Solubility Level | BBB Level | Hepatotoxicity | Absorption Level | PPB Level |
|---|---|---|---|---|---|---|
| 1 | Compound2265 | 3 | 4 | 0 | 3 | 0 |
| 2 | Compound531 | 3 | 4 | 0 | 3 | 0 |
| 3 | Compound529 | 4 | 4 | 0 | 3 | 0 |
| 4 | Compound532 | 3 | 4 | 0 | 3 | 0 |
| 5 | Compound1334 | 4 | 4 | 1 | 3 | 0 |
| 6 | Compound5292 | 3 | 4 | 0 | 3 | 0 |
| 7 | Compound4759 | 2 | 4 | 0 | 3 | 0 |
| 8 | Compound2293 | 2 | 4 | 1 | 2 | 0 |
| 9 | Compound4419 | 5 | 4 | 1 | 3 | 0 |
| 10 | Compound8826 | 1 | 4 | 0 | 3 | 1 |
| 11 | Compound1926 | 3 | 4 | 1 | 2 | 0 |
| 12 | Compound5481 | 5 | 4 | 1 | 3 | 0 |
| 13 | Compound2295 | 3 | 4 | 1 | 2 | 1 |
| 14 | Compound5520 | 2 | 4 | 1 | 3 | 0 |
| 15 | Compound2476 | 3 | 4 | 0 | 3 | 1 |
| 16 | Compound8511 | 1 | 4 | 1 | 2 | 1 |
| 17 | Compound2240 | 3 | 2 | 1 | 0 | 0 |
| 18 | Compound7726 | 2 | 4 | 1 | 2 | 1 |
| 19 | Compound2101 | 2 | 4 | 1 | 3 | 0 |
| 20 | Compound8477 | 2 | 4 | 1 | 2 | 1 |
| 21 | Compound4965 | 2 | 4 | 0 | 3 | 0 |
| Enzyme | XMP, K0.5 µM | XMP, Km µM | ATP, Km µM | Glutamine, Km µM | (NH4)2SO4, Km mM |
|---|---|---|---|---|---|
| Ca. L. asiaticus | 61.6 ± 3.9 | - | 258.2 ± 24.89 | 215 ± 26.76 | 10.92 ± 1.30 |
| M. tuberculosisa | 45 ± 1 | - | 27 ± 2 | 1.24 (±0.06) × 103 | 13 ± 1 |
| P. falciparumb | - | 16.8 ± 2 | 260 ± 38 | 472 ± 69 | 5.4 ± 0.8 |
| E. colic | - | 29 | 530 | 1000 | 1.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nan, J.; Zhang, S.; Zhan, P.; Jiang, L. Discovery of Novel GMPS Inhibitors of Candidatus Liberibacter Asiaticus by Structure Based Design and Enzyme Kinetic. Biology 2021, 10, 594. https://doi.org/10.3390/biology10070594
Nan J, Zhang S, Zhan P, Jiang L. Discovery of Novel GMPS Inhibitors of Candidatus Liberibacter Asiaticus by Structure Based Design and Enzyme Kinetic. Biology. 2021; 10(7):594. https://doi.org/10.3390/biology10070594
Chicago/Turabian StyleNan, Jing, Shaoran Zhang, Ping Zhan, and Ling Jiang. 2021. "Discovery of Novel GMPS Inhibitors of Candidatus Liberibacter Asiaticus by Structure Based Design and Enzyme Kinetic" Biology 10, no. 7: 594. https://doi.org/10.3390/biology10070594
APA StyleNan, J., Zhang, S., Zhan, P., & Jiang, L. (2021). Discovery of Novel GMPS Inhibitors of Candidatus Liberibacter Asiaticus by Structure Based Design and Enzyme Kinetic. Biology, 10(7), 594. https://doi.org/10.3390/biology10070594