
Research on the Mechanisms of Plant Enrichment and Detoxification of Cadmium


Abstract
Simple Summary
Abstract
1. Introduction
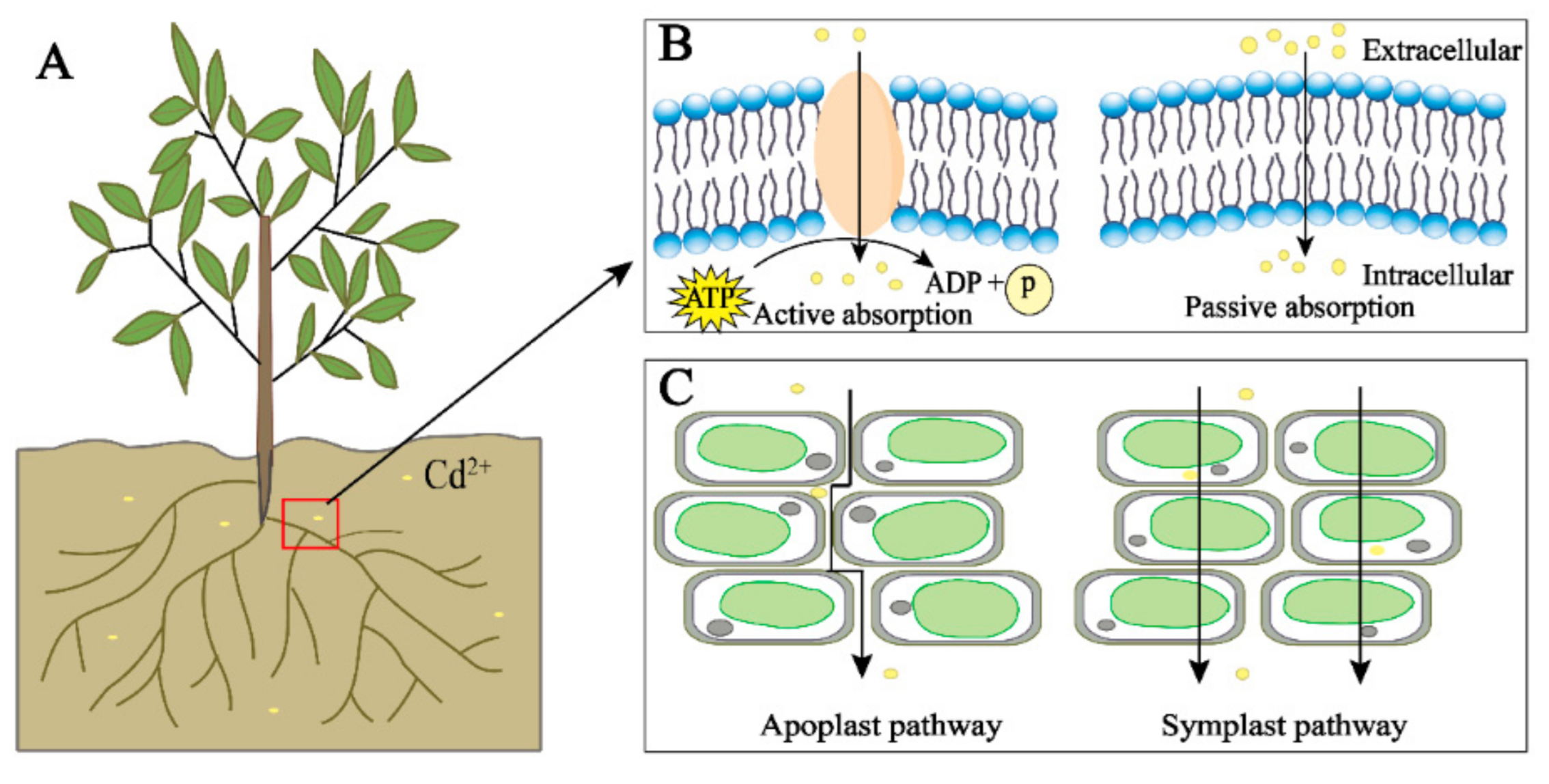
2. Uptake, Transport, and Distribution of Cd in Plants
2.1. Pathways of Cd Uptake by Plants
2.2. Plant Transport of Cd and Involved Transporters
2.3. Distribution and Subcellular Distribution of Cd in Plants
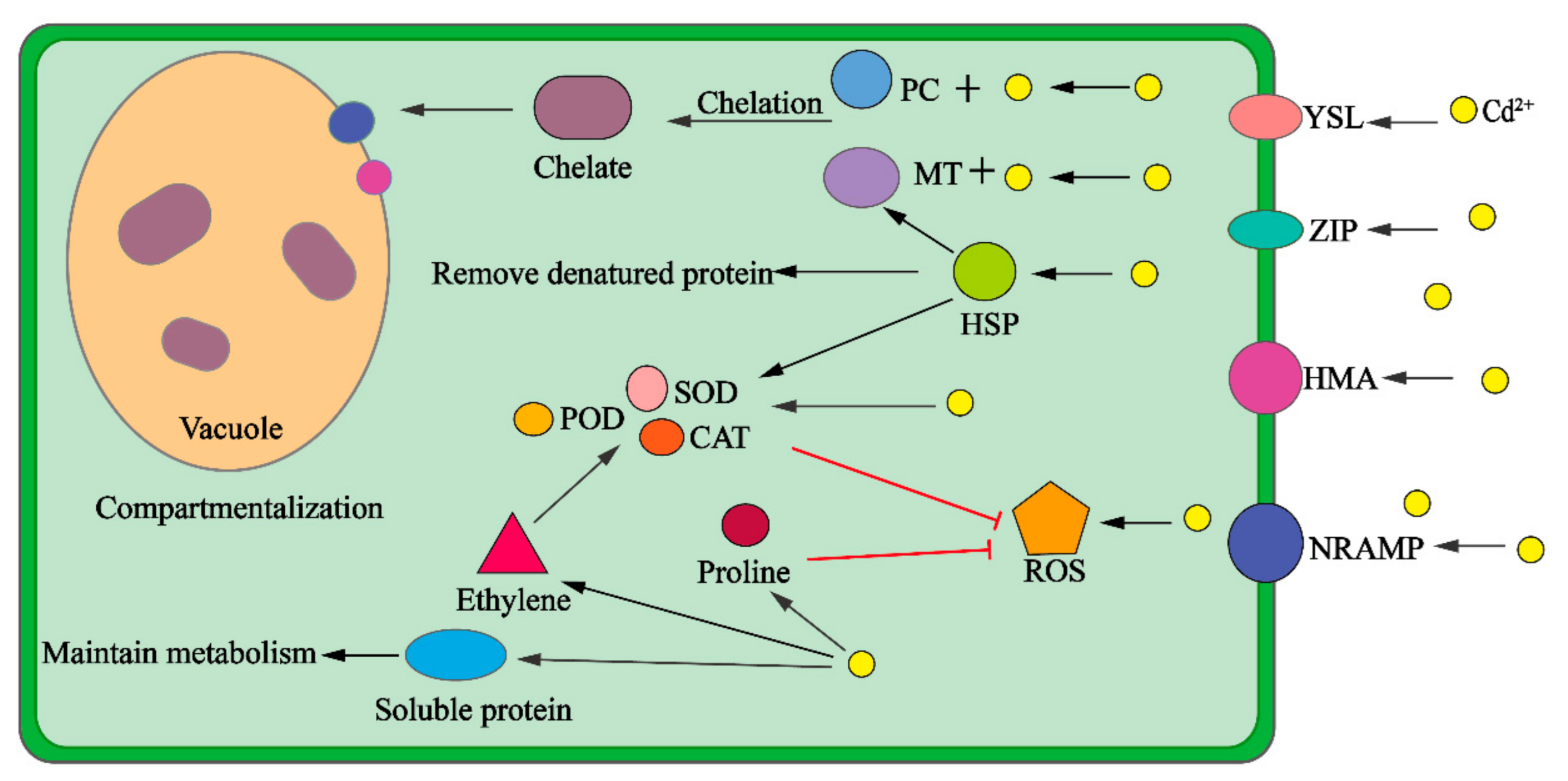
3. The Mechanisms of Plant Enrichment of Cd
3.1. The Role of Plant Roots
3.2. Compartmentalization and Chelation
3.3. Antioxidant System
3.4. Stress
3.5. Osmotic Adjustment
4. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Jeong, H.; Choi, J.Y.; Lee, J.; Lim, J.; Ra, K. Heavy metal pollution by road-deposited sediments and its contribution to total suspended solids in rainfall runoff from intensive industrial areas. Environ. Pollut. 2020, 265, 115028. [Google Scholar] [CrossRef] [PubMed]
- Weissmannová, H.D.; Mihočová, S.; Chovanec, P.; Pavlovský, J. Potential Ecological Risk and Human Health Risk Assessment of Heavy Metal Pollution in Industrial Affected Soils by Coal Mining and Metallurgy in Ostrava, Czech Republic. Int. J. Environ. Res. Public Health 2019, 16, 4495. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.; Yang, K.; Lu, W.; Cui, K.; Li, J.; Liang, Y.; Hou, G.; Zhao, X.; Li, H. An overview of heavy metal pollution in Chaohu Lake, China: Enrichment, distribution, speciation, and associated risk under natural and anthropogenic changes. Environ. Sci. Pollut. Res. 2019, 26, 29585–29596. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.G.; Zhou, Q.X.; Wang, X. Reused Path of Heavy Metal Pollution in Soils and its Research Advance. J. Basic Sci. Engine 2003, 02, 143–151. [Google Scholar]
- Zhao, F.-J.; Ma, Y.; Zhu, Y.-G.; Tang, Z.; McGrath, S.P. Soil contamination in China: Current status and mitigation strategies. Environ. Sci. Technol. 2015, 49, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Xue, N.; Han, Z. A meta-analysis of heavy metals pollution in farmland and urban soils in China over the past 20 years. J. Environ. Sci. 2021, 101, 217–226. [Google Scholar] [CrossRef]
- Yang, Q.; Li, Z.; Lu, X.; Duan, Q.; Huang, L.; Bi, J. A review of soil heavy metal pollution from industrial and agricultural regions in China: Pollution and risk assessment. Sci. Total Environ. 2018, 642, 690–700. [Google Scholar] [CrossRef]
- Shahid, M.; Dumat, C.; Khalid, S.; Niazi, N.K.; Antunes, P.M.C. Cadmium Bioavailability, Uptake, Toxicity and Detoxification in Soil-Plant System. Rev. Environ. Contam. Toxicol. 2016, 241, 73–137. [Google Scholar] [CrossRef]
- Wang, P.; Deng, X.; Huang, Y.; Fang, X.; Zhang, J.; Wan, H.; Yang, C. Comparison of subcellular distribution and chemical forms of cadmium among four soybean cultivars at young seedlings. Environ. Sci. Pollut. Res. 2015, 22, 19584–19595. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.J.; Liu, J.T.; Ma, S.H.; Zhao, S.Z.; Zhong, R.; Liu, Y.B.; Zhao, J.Z. Occurrence characteristics of cadmium in atmospheric dustfall in Baotou City. J. Inner. Mong. Agric. Univ. 2010, 31, 105–109. [Google Scholar]
- Zhang, C.; Zhang, P.; Mo, C.; Yang, W.; Li, Q.; Pan, L.; Lee, D.K. Cadmium uptake, chemical forms, subcellular distribution, and accumulation in Echinodorus osiris Rataj. Environ. Sci. Process. Impacts 2013, 15, 1459–1465. [Google Scholar] [CrossRef]
- Li, G.; Li, Q.; Wang, L.; Zhang, D. Cadmium tolerance and detoxification in Myriophyllum aquaticum: Physiological responses, chemical forms, and subcellular distribution. Environ. Sci. Pollut. Res. Int. 2020, 27, 37733–37744. [Google Scholar] [CrossRef] [PubMed]
- Xin, J.; Zhao, X.; Tan, Q.; Sun, X.; Hu, C. Comparison of cadmium absorption, translocation, subcellular distribution and chemical forms between two radish cultivars (Raphanus sativus L.). Ecotoxicol. Environ. Saf. 2017, 145, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Augustsson, A.; Uddh-Söderberg, T.; Filipsson, M.; Helmfrid, I.; Berglund, M.; Karlsson, H.; Hogmalm, J.; Karlsson, A.; Alriksson, S. Challenges in assessing the health risks of consuming vegetables in metal-contaminated environments. Environ. Int. 2018, 113, 269–280. [Google Scholar] [CrossRef]
- Telford, J.N.; Thonney, M.; Hogue, D.E.; Stouffer, J.R.; Bache, C.A.; Gutenmann, W.H.; Lisk, D.J.; Babish, J.G.; Stoewsand, G.S. Toxicologic studies in growing sheep fed silage corn cultured on municipal sludge-amended acid subsoil. J. Toxicol. Environ. Health Part A 1982, 10, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Bora, M.S.; Gogoi, N.; Sarma, K.P. Tolerance mechanism of cadmium in Ceratopteris pteridoides: Translocation and subcellular distribution. Ecotoxicol. Environ. Safe 2020, 197, 110599. [Google Scholar] [CrossRef]
- Farooq, A.; Nadeem, M.; Abbas, G.; Shabbir, A.; Khalid, M.S.; Javeed, H.M.R.; Saeed, M.F.; Akram, A.; Younis, A.; Akhtar, G. Cadmium Partitioning, Physiological and Oxidative Stress Responses in Marigold (Calendula calypso) Grown on Contaminated Soil: Implications for Phytoremediation. Bull. Environ. Contam. Toxicol. 2020, 105, 270–276. [Google Scholar] [CrossRef]
- Jolly, Y.N.; Islam, A.; Akbar, S. Transfer of metals from soil to vegetables and possible health risk assessment. SpringerPlus 2013, 2, 1–8. [Google Scholar] [CrossRef]
- Brams, E.; Anthony, W. Cadmium and lead through an agricultural food chain. Sci. Total Environ. 1983, 28, 295–306. [Google Scholar] [CrossRef]
- Paithankar, J.G.; Saini, S.; Dwivedi, S.; Sharma, A.; Chowdhuri, D.K. Heavy metal associated health hazards: An interplay of oxidative stress and signal transduction. Chemosphere 2021, 262, 128350. [Google Scholar] [CrossRef]
- Cheng, C.-H.; Ma, H.-L.; Deng, Y.-Q.; Feng, J.; Jie, Y.-K.; Guo, Z.-X. Oxidative stress, cell cycle arrest, DNA damage and apoptosis in the mud crab (Scylla paramamosain) induced by cadmium exposure. Chemosphere 2021, 263, 128277. [Google Scholar] [CrossRef] [PubMed]
- Júnior, J.E.G.P.; Moraes, P.Z.; Rodriguez, M.D.; Simões, M.R.; Cibin, F.; Pinton, S.; Junior, F.B.; Peçanha, F.M.; Vassallo, D.V.; Miguel, M.; et al. Cadmium exposure activates NADPH oxidase, renin-angiotensin system and cyclooxygenase 2 pathways in arteries, inducing hypertension and vascular damage. Toxicol. Lett. 2020, 333, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Satarug, S.; Moore, M.R. Adverse health effects of chronic exposure to low-level cadmium in foodstuffs and cigarette smoke. Environ. Health Perspect. 2004, 112, 1099–1103. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Wang, K.; Liu, Z.; Yu, Y.; Wang, Q.; Li, H. Effect of selenium on the subcellular distribution of cadmium and oxidative stress induced by cadmium in rice (Oryza sativa L.). Environ. Sci. Pollut. Res. 2019, 26, 16220–16228. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.R.; Zhang, P.; Du, S.T. Oxidative stress-related signals and their regulation under Cd stress: A review. J. Appl. Ecol. 2016, 27, 981–992. [Google Scholar]
- Singh, S.; Eapen, S.; D’Souza, S. Cadmium accumulation and its influence on lipid peroxidation and antioxidative system in an aquatic plant, Bacopa monnieri L. Chemosphere 2006, 62, 233–246. [Google Scholar] [CrossRef]
- Huang, S.; Song, Q.; Li, Q.; Zhang, H.; Luo, X.; Zheng, Z. Damage of heavy metals to Vallisneria natans (V. natans) and characterization of microbial community in biofilm. Aquat. Toxicol. 2020, 225, 105515. [Google Scholar] [CrossRef]
- Andresen, E.; Kappel, S.; Stärk, H.; Riegger, U.; Borovec, J.; Mattusch, J.; Heinz, A.; Schmelzer, C.; Matoušková, Š.; Dickinson, B.; et al. Cadmium toxicity investigated at the physiological and biophysical levels under environmentally relevant conditions using the aquatic model plant Ceratophyllum demersum. New Phytol. 2016, 210, 1244–1258. [Google Scholar] [CrossRef]
- Liu, S.; Ali, S.; Yang, R.; Tao, J.; Ren, B. A newly discovered Cd-hyperaccumulator Lantana camara L. J. Hazard. Mater. 2019, 371, 233–242. [Google Scholar] [CrossRef]
- Hu, Y.; Tian, S.; Foyer, C.H.; Hou, D.; Wang, H.; Zhou, W.; Liu, T.; Ge, J.; Lu, L.; Lin, X. Efficient phloem transport significantly remobilizes cadmium from old to young organs in a hyperaccumulator Sedum alfredii. J. Hazard. Mater. 2019, 365, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Küpper, H.; Leitenmaier, B. Cadmium-accumulating plants. Met. Ions Life Sci. 2012, 11, 373–393. [Google Scholar] [CrossRef]
- Jaffre, T.; Brooks, R.R.; Lee, J.; Reeves, R.D. Sebertia acuminata: A Hyperaccumulator of Nickel from New Caledonia. Science 1976, 193, 579–580. [Google Scholar] [CrossRef]
- Yu, B.; Peng, Y.; Xu, J.; Qin, D.; Gao, T.; Zhu, H.; Zuo, S.; Song, H.; Dong, J. Phytoremediation potential of Youngia japonica (L.) DC: A newly discovered cadmium hyperaccumulator. Environ. Sci. Pollut. Res. 2021, 28, 6044–6057. [Google Scholar] [CrossRef]
- Zhong, L.; Lin, L.; Liao, M.; Wang, J.; Tang, Y.; Sun, G.; Liang, D.; Xia, H.; Wang, X.; Zhang, H.; et al. Phytoremediation potential of Pterocypsela laciniata as a cadmium hyperaccumulator. Environ. Sci. Pollut. Res. 2019, 26, 13311–13319. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, H.; Wu, L.; Liu, A.; Zhao, F.; Xu, W. Heavy metal ATPase 3 (HMA3) confers cadmium hypertolerance on the cadmium/zinc hyperaccumulator Sedum plumbizincicola. New Phytol. 2017, 215, 687–698. [Google Scholar] [CrossRef]
- Lan, X.-Y.; Yan, Y.-Y.; Yang, B.; Li, X.-Y.; Xu, F.-L. Subcellular distribution of cadmium in a novel potential aquatic hyperaccumulator—Microsorum pteropus. Environ. Pollut. 2019, 248, 1020–1027. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.T. Research on Cd Absorption, Accumulation and Tolerance of Taraxacum ohwianum Kitam. Ph.D. Thesis, Shenyang Agricultural University, Shenyang, China, 2019. [Google Scholar]
- Xu, X.; Zhang, S.; Cheng, Z.; Li, T.; Jia, Y.; Wang, G.; Yang, Z.; Xian, J.; Yang, Y.; Zhou, W. Transcriptome analysis revealed cadmium accumulation mechanisms in hyperaccumulator Siegesbeckia orientalis L. Environ. Sci. Pollut. Res. 2020, 27, 18853–18865. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Ueno, D.; Zhao, F.-J.; McGrath, S.P. Subcellular localisation of Cd and Zn in the leaves of a Cd-hyperaccumulating ecotype of Thlaspi caerulescens. Planta 2005, 220, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Küpper, H.; Lombi, E.; Zhao, F.-J.; McGrath, S.P. Cellular compartmentation of cadmium and zinc in relation to other elements in the hyperaccumulator Arabidopsis halleri. Planta 2000, 212, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Kowalkowski, T.; Krakowska, A.; Złoch, M.; Hrynkiewicz, K.; Buszewski, B. Cadmium-affected synthesis of exopolysaccharides by rhizosphere bacteria. J. Appl. Microbiol. 2019, 127, 713–723. [Google Scholar] [CrossRef]
- Liu, S.; Yang, B.; Liang, Y.; Xiao, Y.; Fang, J. Prospect of phytoremediation combined with other approaches for remediation of heavy metal-polluted soils. Environ. Sci. Pollut. Res. 2020, 27, 16069–16085. [Google Scholar] [CrossRef]
- Ismael, M.A.; Elyamine, A.M.; Moussa, M.G.; Cai, M.; Zhao, X.; Hu, C. Cadmium in plants: Uptake, toxicity, and its interactions with selenium fertilizers. Metallomics 2019, 11, 255–277. [Google Scholar] [CrossRef] [PubMed]
- Riaz, M.; Kamran, M.; Rizwan, M.; Ali, S.; Parveen, A.; Malik, Z.; Wang, X. Cadmium uptake and translocation: Selenium and silicon roles in Cd detoxification for the production of low Cd crops: A critical review. Chemosphere 2021, 273, 129690. [Google Scholar] [CrossRef] [PubMed]
- Li, J.T.; Gurajala, H.K.; Wu, L.H.; Ent, A.; Qiu, R.L.; Baker, A.J.M.; Tang, Y.T.; Yang, X.E.; Shu, W.S. Hyperaccumulator Plants from China: A Synthesis of the Current State of Knowledge. Environ. Sci. Technol. 2018, 52, 11980–11994. [Google Scholar] [CrossRef] [PubMed]
- Cutler, J.M.; Rains, D.W. Characterization of cadmium uptake by plant tissue. Plant Physiol. 1974, 54, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.M.; Chirgawi, M. The assessment of air and soil as contributors of some trace metals to vegetable plants. II. Translocation of atmospheric and laboratory-generated cadmium aerosols to and within vegetable plants. Sci. Total Environ. 1989, 83, 35–45. [Google Scholar] [CrossRef]
- Hauser, L.; Tandy, S.; Schulin, R.; Nowack, B. Column extraction of heavy metals from soils using the biodegradable chelating agent EDDS. Environ. Sci. Technol. 2005, 39, 6819–6824. [Google Scholar] [CrossRef] [PubMed]
- Meychik, N.R.; Nikolaeva, Y.I.; Yermakov, I.P. Ion-exchange properties of cell walls of Spinacia oleracea L. roots under different environmental salt conditions. Biochemistry 2006, 71, 781–789. [Google Scholar] [CrossRef]
- Palusińska, M.; Barabasz, A.; Kozak, K.; Papierniak, A.; Maślińska, K.; Antosiewicz, D.M. Zn/Cd status-dependent accumulation of Zn and Cd in root parts in tobacco is accompanied by specific expression of ZIP genes. BMC Plant Boil. 2020, 20, 37. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Garland, T.R.; Wildung, R.E. Cadmium uptake kinetics in intact soybean plants. Plant Physiol. 1983, 73, 844–848. [Google Scholar] [CrossRef]
- Peng, K.J.; Luo, C.L.; Chen, Y.H.; Wang, G.P.; Li, X.D.; Shen, Z.G. Cadmium and other metal uptake by Lobelia chinensis and Solanum nigrum from contaminated soils. Bull. Environ. Contam. Toxicol. 2009, 83, 260–264. [Google Scholar] [CrossRef]
- Hart, J.J.; Welch, R.M.; Norvell, W.A.; Kochian, L.V. Transport interactions between cadmium and zinc in roots of bread and durum wheat seedlings. Physiol. Plant 2002, 116, 73–78. [Google Scholar] [CrossRef]
- De Temmerman, L.; Ruttens, A.; Waegeneers, N. Impact of atmospheric deposition of As, Cd and Pb on their concentration in carrot and celeriac. Environ. Pollut. 2012, 166, 187–195. [Google Scholar] [CrossRef]
- Fernández, E.A.; Rossini, O.S. The composition and relationships between trace element levels in inhalable atmospheric particles (PM10) and in leaves of Nerium oleander L. and Lantana camara L/. Chemosphere 2006, 62, 1665–1672. [Google Scholar] [CrossRef] [PubMed]
- Anda, A.; Illés, B.; Soós, G. Effect of cadmium pollution of atmospheric origin on field-grown maize in two consecutive years with diverse weather conditions. Acta Biol. Hung. 2013, 64, 476–489. [Google Scholar] [CrossRef]
- Lu, H.; Zhang, Y.L.; Wu, Y.B. Research progress on foliar absorption mechanism of pollutants. Environ. Chem. 2020, 12, 3371–3383. [Google Scholar]
- Huang, X.; Duan, S.; Wu, Q.; Yu, M.; Shabala, S. Reducing Cadmium Accumulation in Plants: Structure-Function Relations and Tissue-Specific Operation of Transporters in the Spotlight. Plants 2020, 9, 223. [Google Scholar] [CrossRef] [PubMed]
- Da Cunha, K.P.V.; Nascimento, C.W.A.D.; Pimentel, R.M.D.M.; Ferreira, C.P. Cellular localization of cadmium and structural changes in maize plants grown on a cadmium contaminated soil with and without liming. J. Hazard. Mater. 2008, 160, 228–234. [Google Scholar] [CrossRef]
- Curie, C.; Cassin, G.; Couch, D.; Divol, F.; Higuchi, K.; Le Jean, M.; Misson, J.; Schikora, A.; Czernic, P.; Mari, S. Metal movement within the plant: Contribution of nicotianamine and yellow stripe 1-like transporters. Ann. Bot. 2008, 103, 1–11. [Google Scholar] [CrossRef]
- Curie, C.; Panaviene, Z.; Loulergue, C.; Dellaporta, S.L.; Briat, J.-F.; Walker, E.L. Maize yellow stripe1 encodes a membrane protein directly involved in Fe (III) uptake. Nat. Cell Biol. 2001, 409, 346–349. [Google Scholar] [CrossRef]
- Guerinot, M.L. The ZIP family of metal transporters. Biochim. Biophys. Acta Biomembr. 2000, 1465, 190–198. [Google Scholar] [CrossRef]
- Grotz, N.; Fox, T.; Connolly, E.; Park, W.; Guerinot, M.L.; Eide, D. Identification of a family of zinc transporter genes from Arabidopsis that respond to zinc deficiency. Proc. Natl. Acad. Sci. USA 1998, 95, 7220–7224. [Google Scholar] [CrossRef]
- Takahashi, R.; Bashir, K.; Ishimaru, Y.; Nishizawa, N.K.; Nakanishi, H. The role of heavy-metal ATPases, HMAs, in zinc and cadmium transport in rice. Plant Signal. Behav. 2012, 7, 1605–1607. [Google Scholar] [CrossRef]
- Williams, L.E.; Mills, R.F. P (1B)-ATPases—An ancient family of transition metal pumps with diverse functions in plants. Trends Plant Sci. 2005, 10, 491–502. [Google Scholar] [CrossRef]
- Milner, M.J.; Mitani, U.N.; Yamaji, N.; Yokosho, K.; Craft, E.; Fei, Z.; Ebbs, S.; Clemencia, Z.M.; Ma, J.F.; Kochian, L.V. Root and shoot transcriptome analysis of two ecotypes of Noccaea caerulescens uncovers the role of NcNramp1 in Cd hyperaccumulation. Plant J. 2014, 78, 398–410. [Google Scholar] [CrossRef]
- Sasaki, A.; Yamaji, N.; Yokosho, K.; Ma, J.F. Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice. Plant Cell 2012, 24, 2155–2167. [Google Scholar] [CrossRef]
- Zhao, F.-J.; Huang, X. Cadmium Phytoremediation: Call Rice CAL1. Mol. Plant 2018, 11, 640–642. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.-S.; Huang, J.; Zeng, D.-L.; Peng, J.-S.; Zhang, G.-B.; Ma, H.-L.; Guan, Y.; Yi, H.-Y.; Fu, Y.-L.; Han, B.; et al. A defensin-like protein drives cadmium efflux and allocation in rice. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.D.; Zhao, K.X.; Yang, Z.M. Identification of genomic ATP binding cassette (ABC) transporter genes and Cd-responsive ABCs in Brassica napus. Gene 2018, 664, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Schaaf, G.; Ludewig, U.; Erenoglu, B.E.; Mori, S.; Kitahara, T.; von Wirén, N. ZmYS1 functions as a proton-coupled symporter for phytosiderophore- and nicotianamine-chelated metals. J. Biol. Chem. 2004, 279, 9091–9096. [Google Scholar] [CrossRef]
- Murata, Y.; Ma, J.F.; Yamaji, N.; Ueno, D.; Nomoto, K.; Iwashita, T. A specific transporter for iron (III)-phytosiderophore in barley roots. Plant J. 2006, 46, 563–572. [Google Scholar] [CrossRef]
- von Wirén, N.; Mori, S.; Marschner, H.; Romheld, V. Iron Inefficiency in Maize Mutant ys1 (Zea mays L. cv Yellow-Stripe) Is Caused by a Defect in Uptake of Iron Phytosiderophores. Plant Physiol. 1994, 106, 71–77. [Google Scholar] [CrossRef]
- Das, S.; Sen, M.; Saha, C.; Chakraborty, D.; Das, A.; Banerjee, M.; Seal, A. Isolation and expression analysis of partial sequences of heavy metal transporters from Brassica juncea by coupling high throughput cloning with a molecular fingerprinting technique. Planta 2011, 234, 139–156. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.; Zhang, Y.-X.; Chai, T.-Y. Molecular cloning and characterization of a Brassica juncea yellow stripe-like gene, BjYSL7, whose overexpression increases heavy metal tolerance of tobacco. Plant Cell Rep. 2013, 32, 651–662. [Google Scholar] [CrossRef]
- Feng, S.; Tan, J.; Zhang, Y.; Liang, S.; Xiang, S.; Wang, H.; Chai, T. Isolation and characterization of a novel cadmium-regulated Yellow Stripe-Like transporter (SnYSL3) in Solanum nigrum. Plant Cell Rep. 2016, 36, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.M.; Guerinot, M.L. Facing the challenges of Cu, Fe and Zn homeostasis in plants. Nat. Chem. Biol. 2009, 5, 333–340. [Google Scholar] [CrossRef]
- Eng, B.; Guerinot, M.; Eide, D.; Saier, M. Sequence analyses and phylogenetic characterization of the ZIP family of metal ion transport proteins. J. Membr. Biol. 1998, 166, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Nishida, S.; Mizuno, T.; Obata, H. Involvement of histidine-rich domain of ZIP family transporter TjZNT1 in metal ion specificity. Plant Physiol. Biochem. 2008, 46, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, X.; Tang, L.; Peng, Y.; Qian, M.; Guo, Y.; Rui, H.; Zhang, F.; Hu, Z.; Chen, Y.; et al. The root iron transporter 1 governs cadmium uptake in Vicia sativa roots. J. Hazard. Mater. 2020, 398, 122873. [Google Scholar] [CrossRef]
- Wu, X.; Chen, J.; Yue, X.; Wei, X.; Zou, J.; Chen, Y.; Su, N.; Cui, J. The zinc-regulated protein (ZIP) family genes and glutathione s-transferase (GST) family genes play roles in Cd resistance and accumulation of pak choi (Brassica campestris ssp. chinensis). Ecotoxicol. Environ. Saf. 2019, 183, 109571. [Google Scholar] [CrossRef]
- Williams, L.E.; Pittman, J.K.; Hall, J. Emerging mechanisms for heavy metal transport in plants. Biochim. Biophys. Acta Biomembr. 2000, 1465, 104–126. [Google Scholar] [CrossRef]
- Argüello, J.M. Identification of ion-selectivity determinants in heavy-metal transport P1B-type ATPases. J. Membr. Biol. 2003, 195, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhi, J.; Liu, X.; Zhang, H.; Liu, H.; Xu, J. Transgenic tobacco plants expressing a P1B-ATPase gene from Populus tomentosa Carr. (PtoHMA5) demonstrate improved cadmium transport. Int. J. Biol. Macromol. 2018, 113, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liang, S.; Xiang, W.; Dai, H.; Duan, Y.; Kang, F.; Chai, T. A repeat region from the Brassica juncea HMA4 gene BjHMA4R is specifically involved in Cd (2+) binding in the cytosol under low heavy metal concentrations. BMC Plant Biol. 2019, 19, 89. [Google Scholar] [CrossRef]
- Kumagai, S.; Suzuki, T.; Tezuka, K.; Satoh-Nagasawa, N.; Takahashi, H.; Sakurai, K.; Watanabe, A.; Fujimura, T.; Akagi, H. Functional analysis of the C-terminal region of the vacuolar cadmium-transporting rice OsHMA3. FEBS Lett. 2014, 588, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.E.; Cobbett, C.S. HMA P-type ATPases are the major mechanism for root-to-shoot Cd translocation in Arabidopsis thaliana. New Phytol. 2009, 181, 71–78. [Google Scholar] [CrossRef]
- Cun, P.; Sarrobert, C.; Richaud, P.; Chevalier, A.; Soreau, P.; Auroy, P.; Gravot, A.; Baltz, A.; Leonhardt, N.; Vavasseur, A. Modulation of Zn/Cd P (1B2)-ATPase activities in Arabidopsis impacts differently on Zn and Cd contents in shoots and seed. Metallomics 2014, 6, 2109–2116. [Google Scholar] [CrossRef] [PubMed]
- Migocka, M.; Papierniak, A.; Maciaszczyk-Dziubinska, E.; Posyniak, E.; Kosieradzka, A. Molecular and biochemical properties of two P1B2-ATPases, CsHMA3 and CsHMA4, from cucumbe. Plant Cell Environ. 2014, 38, 1127–1141. [Google Scholar] [CrossRef]
- Fleming, M.; Trenor, C.C.; Su, M.A.; Foernzler, D.; Beier, D.R.; Dietrich, W.F.; Andrews, N. Microcytic anaemia mice have a mutation in Nramp2, a candidate iron transporter gene. Nat. Genet. 1997, 16, 383–386. [Google Scholar] [CrossRef]
- Chen, S.; Han, X.; Fang, J.; Lu, Z.; Qiu, W.; Liu, M.; Sang, J.; Jiang, J.; Zhuo, R. Sedum alfredii SaNramp6 Metal Transporter Contributes to Cadmium Accumulation in Transgenic Arabidopsis thaliana. Sci. Rep. 2017, 7, 13318. [Google Scholar] [CrossRef]
- Wang, C.; Chen, X.; Yao, Q.; Long, D.; Fan, X.; Kang, H.; Zeng, J.; Sha, L.; Zhang, H.; Zhou, Y.; et al. Overexpression of TtNRAMP6 enhances the accumulation of Cd in Arabidopsis. Gene 2019, 696, 225–232. [Google Scholar] [CrossRef]
- Wei, W.; Chai, T.; Zhang, Y.; Han, L.; Xu, J.; Guan, Z. The Thlaspi caerulescens NRAMP homologue TcNRAMP3 is capable of divalent cation transport. Mol. Biotechnol. 2009, 41, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Yamaji, N.; Yamane, M.; Kashino-Fujii, M.; Sato, K.; Ma, J.F. The HvNramp5 Transporter Mediates Uptake of Cadmium and Manganese, But Not Iron. Plant Physiol. 2016, 172, 1899–1910. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, M.; Sharma, D.; Dwivedi, S.; Singh, M.; Tripathi, R.D.; Trivedi, P.K. Expression in Arabidopsis and cellular localization reveal involvement of rice NRAMP, OsNRAMP1, in arsenic transport and tolerance. Plant Cell Environ. 2014, 37, 140–152. [Google Scholar] [CrossRef] [PubMed]
- Spielmann, J.; Ahmadi, H.; Scheepers, M.; Weber, M.; Nitsche, S.; Carnol, M.; Bosman, B.; Kroymann, J.; Motte, P.; Clemens, S.; et al. The two copies of the zinc and cadmium ZIP6 transporter of Arabidopsis halleri have distinct effects on cadmium tolerance. Plant Cell Environ. 2020, 43, 2143–2157. [Google Scholar] [CrossRef] [PubMed]
- Safe, S.; Naira, V.; Karki, K. Metformin-induced anticancer activities: Recent insights. Biol. Chem. 2018, 399, 321–335. [Google Scholar] [CrossRef]
- Sasaki, A.; Yamaji, N.; Ma, J.F. Overexpression of OsHMA3 enhances Cd tolerance and expression of Zn transporter genes in rice. J. Exp. Bot. 2014, 65, 6013–6021. [Google Scholar] [CrossRef]
- Liu, X.S.; Feng, S.J.; Zhang, B.Q.; Wang, M.Q.; Cao, H.W.; Rono, J.K.; Chen, X.; Yang, Z.M. OsZIP1 functions as a metal efflux transporter limiting excess zinc, copper and cadmium accumulation in rice. BMC Plant Biol. 2019, 19, 1–16. [Google Scholar] [CrossRef]
- Tian, S.; Liang, S.; Qiao, K.; Wang, F.; Zhang, Y.; Chai, T. Co-expression of multiple heavy metal transporters changes the translocation, accumulation, and potential oxidative stress of Cd and Zn in rice (Oryza sativa). J. Hazard. Mater. 2019, 380, 120853. [Google Scholar] [CrossRef]
- Küpper, H.; Kochian, L.V. Transcriptional regulation of metal transport genes and mineral nutrition during acclimatization to cadmium and zinc in the Cd/Zn hyperaccumulator, Thlaspi caerulescens (Ganges population). New Phytol. 2010, 185, 114–129. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, L.; Zhao, F.-J.; Wu, L.; Liu, A.; Xu, W. SpHMA1 is a chloroplast cadmium exporter protecting photochemical reactions in the Cd hyperaccumulator Sedum plumbizincicola. Plant Cell Environ. 2018, 42, 1112–1124. [Google Scholar] [CrossRef]
- Yu, R.; Li, D.; Du, X.; Xia, S.; Liu, C.; Shi, G. Comparative transcriptome analysis reveals key cadmium transport-related genes in roots of two pak choi (Brassica rapa L. ssp. chinensis) cultivars. BMC Genom. 2017, 18, 587. [Google Scholar] [CrossRef]
- Wu, S.; Shi, K.; Hu, C.; Guo, J.; Tan, Q.; Sun, X. Non-invasive microelectrode cadmium flux measurements reveal the decrease of cadmium uptake by zinc supply in pakchoi root (Brassica chinensis L.). Ecotoxicol. Environ. Saf. 2019, 168, 363–368. [Google Scholar] [CrossRef]
- Wu, X.; Zhu, Z.B.; Chen, J.H.; Huang, Y.F.; Liu, Z.L.; Zou, J.W.; Chen, Y.H.; Na Su, N.; Cui, J. Transcriptome analysis revealed pivotal transporters involved in the reduction of cadmium accumulation in pak choi (Brassica chinensis L.) by exogenous hydrogen-rich water. Chemosphere 2019, 216, 684–697. [Google Scholar] [CrossRef] [PubMed]
- Banakar, R.; Alvarez, F.Á.; Abadía, J.; Capell, T.; Christou, P. The expression of heterologous Fe (III) phytosiderophore transporter HvYS1 in rice increases Fe uptake, translocation and seed loading and excludes heavy metals by selective Fe transport. Plant Biotechnol. J. 2017, 15, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Xiao, X.; Wu, X. Physiological and molecular mechanism of cadmium (Cd) tolerance at initial growth stage in rapeseed (Brassica napus L.). Ecotoxicol. Environ. Saf. 2020, 197, 110613. [Google Scholar] [CrossRef]
- Peng, F.; Wang, C.; Zhu, J.; Zeng, J.; Kang, H.; Fan, X.; Sha, L.; Zhang, H.; Zhou, Y.; Wang, Y. Expression of TpNRAMP5, a metal transporter from Polish wheat (Triticum polonicum L.), enhances the accumulation of Cd, Co and Mn in transgenic Arabidopsis plants. Planta 2018, 247, 1395–1406. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Cai, H.; Li, J.; Lv, Y.; Zhang, W.; Zhao, F.-J. Allelic Variation of NtNramp5 Associated with Cultivar Variation in Cadmium Accumulation in Tobacco. Plant Cell Physiol. 2017, 58, 1583–1593. [Google Scholar] [CrossRef]
- Chen, Y.C.; Zhao, Z.J.; Sun, P.S. Distribution and control of heavy metals by addtives in soil-vegetable system in Chongqing. J. Agro Environ. Sci. 2003, 01, 44–47. [Google Scholar]
- Lan, X.-Y.; He, Q.-S.; Yang, B.; Yan, Y.-Y.; Li, X.-Y.; Xu, F.-L. Influence of Cd exposure on H (+) and Cd (2+) fluxes in the leaf, stem and root of a novel aquatic hyperaccumulator—Microsorum pteropus. Chemosphere 2020, 249, 126552. [Google Scholar] [CrossRef]
- Lan, H.X. Effect of Pb, Cd and Their Compound Pollution on the Physilogical and Ecological Characteristics of the Tea. Master’s Thesis, Sichuan Agricultural University, Ya’an, Sichuan Province, China, 2008. [Google Scholar]
- Yu, Z.Y. Study on the Difference of Different Types of Soybean from Cadmium Stress. Master’s Thesis, Shenyang Agricultural University, Shenyang, China, 2018. [Google Scholar]
- He, Y. Studies on the Accumulation Characters and Influence Mechanism of Cadmium in Tobacco. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2014. [Google Scholar]
- Guo, Z. Study on Mechanisms of Physiological Response of Hyperaccumulator Plant Solanum nigrum L. to Cadmium Stress. Ph.D. Thesis, Shanghai Jiao Tong University, Shanghai, China, 2009. [Google Scholar]
- Yang, L.P.; Zhu, J.; Wang, P.; Zeng, J.; Tan, R.; Yang, Y.Z.; Liu, Z.M. Effect of Cd on growth, physiological response, Cd subcellular distribution and chemical forms of Koelreuteria paniculata. Ecotoxicol. Environ. Safe 2018, 160, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.-Z.; Jiang, Y.-B.; Jia, C.-H.; Jiang, S.-M.; Yan, X.-P. Subcellular distribution and chemical forms of cadmium in Morus alba L. Int. J. Phytoremediation 2018, 20, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Li, L.; Duan, Q.; Liu, X.; Chen, M. Progress in our understanding of plant responses to the stress of heavy metal cadmium. Plant Signal. Behav. 2021, 16, 1836884. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Li, Z.; Li, S.; Deng, N.; Mei, P. Effects of root exudates on the activation and remediation of cadmium ion in contaminated soils. Environ. SCI Pollut. Res. Int. 2020, 27, 2926–2934. [Google Scholar] [CrossRef] [PubMed]
- Caffall, K.H.; Mohnen, D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharide. Carbohydr. Res. 2009, 344, 1879–1900. [Google Scholar] [CrossRef] [PubMed]
- Brewin, L.E.; Mehra, A.; Lynch, P.T.; Farago, M.E. Mechanisms of copper tolerance by Armeria maritima in Dolfrwyong Bog, north Wales—initial studies. Environ. Geochem. Health 2003, 25, 147–156. [Google Scholar] [CrossRef]
- Guo, J.; Xu, W.; Ma, M. The assembly of metals chelation by thiols and vacuolar compartmentalization conferred increased tolerance to and accumulation of cadmium and arsenic in transgenic Arabidopsis thaliana. J. Hazard. Mater. 2012, 199–200, 309–313. [Google Scholar] [CrossRef]
- Anjum, N.A.; Hasanuzzaman, M.; Hossain, M.A.; Thangavel, P.; Roychoudhury, A.; Gill, S.S.; Rodrigo, M.A.M.; Adam, V.; Fujita, M.; Kizek, R.; et al. Jacks of metal/metalloid chelation trade in plants—An overview. Front. Plant Sci. 2015, 6, 192. [Google Scholar] [CrossRef]
- Morris, C.A.; Nicolaus, B.; Sampson, V.; Harwood, J.L.; Kille, P. Identification and characterization of a recombinant metallothionein protein from a marine alga, Fucus vesiculosus. Biochem. J. 1999, 338, 553–560. [Google Scholar] [CrossRef]
- Zhang, H.; Lv, S.; Xu, H.; Hou, D.; Li, Y.; Wang, F. H2O2 Is Involved in the Metallothionein-Mediated Rice Tolerance to Copper and Cadmium Toxicity. Int. J. Mol. Sci. 2017, 18, 10. [Google Scholar] [CrossRef]
- Rono, J.K.; Wang, L.; Wu, X.C.; Cao, H.W.; Zhao, Y.N.; Khan, I.U.; Yang, Z.M. Identification of a new function of metallothionein-like gene OsMT1e for cadmium detoxification and potential phytoremediation. Chemosphere 2021, 265, 129136. [Google Scholar] [CrossRef]
- Gu, C.-S.; Liu, L.-Q.; Zhao, Y.-H.; Deng, Y.-M.; Zhu, X.-D.; Huang, S.-Z. Overexpression of Iris lactea var. chinensis metallothionein llMT2a enhances cadmium tolerance in Arabidopsis thaliana. Ecotoxicol. Environ. Saf. 2014, 105, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhi, J.; Li, X.; Zhang, H.; Liu, H.; Xu, J. Diversity in cadmium accumulation and resistance associated with various metallothionein genes (type III) in Phytolacca americana L. Int. J. Biol. Macromol. 2018, 108, 704–709. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Tripathi, S.; Shanker, K.; Sharma, A. Cadmium-induced conformational changes in type 2 metallothionein of medicinal plant Coptis japonica: Insights from molecular dynamics studies of apo, partially and fully metalated forms. J. Biomol. Struct. Dyn. 2018, 37, 1520–1533. [Google Scholar] [CrossRef] [PubMed]
- Vatamaniuk, O.K.; Mari, S.; Lu, Y.-P.; Rea, P.A. AtPCS1, a phytochelatin synthase from Arabidopsis: Isolation and in vitro reconstitution. Proc. Natl. Acad. Sci. USA 1999, 96, 7110–7115. [Google Scholar] [CrossRef] [PubMed]
- Grill, E.; Winnacker, E.-L.; Zenk, M.H. Phytochelatins: The principal heavy-metal complexing peptides of higher plants. Science 1985, 230, 674–676. [Google Scholar] [CrossRef]
- Grill, E.; Löffler, S.; Winnacker, E.L.; Zenk, M.H. Phytochelatins, the heavy-metal-binding peptides of plants, are synthesized from glutathione by a specific gamma-glutamylcysteine dipeptidyl transpeptidase (phytochelatin synthase). Proc. Natl. Acad. Sci. USA 1989, 86, 6838–6842. [Google Scholar] [CrossRef]
- Hirschi, K.D.; Korenkov, V.D.; Wilganowski, N.L.; Wagner, G.J. Expression of arabidopsis CAX2 in tobacco. Altered metal accumulation and increased manganese tolerance. Plant Physiol. 2000, 124, 125–134. [Google Scholar] [CrossRef]
- Tong, Y.-P.; Kneer, R.; Zhu, Y.-G. Vacuolar compartmentalization: A second-generation approach to engineering plants for phytoremediation. Trend. Plant Sci. 2004, 9, 7–9. [Google Scholar] [CrossRef]
- Küpper, H.; Mijovilovich, A.; Meyer-Klaucke, W.; Kroneck, P.M. Tissue- and age-dependent differences in the complexation of cadmium and zinc in the cadmium/zinc hyperaccumulator Thlaspi caerulescens (Ganges ecotype) revealed by x-ray absorption spectroscopy. Plant Physiol. 2004, 134, 748–757. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K.J.; Mimura, T. Vacuolar compartmentalization as indispensable component of heavy metal detoxification in plants. Plant Cell Environ. 2016, 39, 1112–1126. [Google Scholar] [CrossRef]
- Allan, D.L.; Jarrell, W.M. Proton and copper adsorption to maize and soybean root cell walls. Plant Physiol. 1989, 89, 823–832. [Google Scholar] [CrossRef]
- Ueno, D.; Yamaji, N.; Kono, I.; Huang, C.F.; Ando, T.; Yano, M.; Ma, J.F. Gene limiting cadmium accumulation in rice. Proc. Natl. Acad. Sci. USA 2010, 107, 16500–16505. [Google Scholar] [CrossRef]
- Weng, B.; Xie, X.; Weiss, D.J.; Liu, J.; Lu, H.; Yan, C. Kandelia obovata (S., L.) Yong tolerance mechanisms to Cadmium: Subcellular distribution, chemical forms and thiol pools. Mar. Pollut. Bull. 2012, 64, 2453–2460. [Google Scholar] [CrossRef]
- Wei, Y.Y.; Meng, M.; Huang, X.F. Tolerance Mechanisms of Cu and Cd by Eucalyptus in Mine Contaminated Soil. Genom. Appl. Biol. 2016, 35, 227–234. [Google Scholar]
- Guan, M.Y.; Zhang, H.H.; Pan, W.; Jin, C.W.; Lin, X.Y. Sulfide alleviates cadmium toxicity in Arabidopsis plants by altering the chemical form and the subcellular distribution of cadmium. Sci. Total Environ. 2018, 627, 663–670. [Google Scholar] [CrossRef]
- Zhou, L.; Zhao, Y.; Wang, S. Cadmium transfer and detoxification mechanisms in a soil-mulberry-silkworm system: Phytoremediation potential. Environ. Sci. Pollut. Res. Int. 2015, 22, 18031–18039. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Lu, L.; Labavitch, J.; Yang, X.; He, Z.; Hu, H.; Sarangi, R.; Newville, M.; Commisso, J.; Brown, P. Cellular sequestration of cadmium in the hyperaccumulator plant species Sedum alfredii. Plant Physiol. 2011, 157, 1914–1925. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Qin, J.; Long, L.; Ma, Y.; Li, H.; Li, K.; Jiang, X.; Liu, T.; Polle, A.; Liang, Z.; et al. Net cadmium flux and accumulation reveal tissue-specific oxidative stress and detoxification in Populus × canescens. Physiol. Plant. 2011, 143, 50–63. [Google Scholar] [CrossRef]
- Watanabe, M.; Suzuki, T. Involvement of reactive oxygen stress in cadmium-induced cellular damage in Euglena gracilis. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2002, 131, 491–500. [Google Scholar] [CrossRef]
- Cuypers, A.; Plusquin, M.; Remans, T.; Jozefczak, M.; Keunen, E.; Gielen, H.; Opdenakker, K.; Nair, A.R.; Munters, E.; Artois, T.J.; et al. Cadmium stress: An oxidative challenge. Biometals 2010, 23, 927–940. [Google Scholar] [CrossRef]
- Jedelská, T.; Kraiczová, V. Šmotková; Berčíková, L.; Činčalová, L.; Luhová, L.; Petřivalský, M. Tomato Root Growth Inhibition by Salinity and Cadmium Is Mediated By S-Nitrosative Modifications of ROS Metabolic Enzymes Controlled by S-Nitrosoglutathione Reductase. Biomolecues 2019, 9, 393. [Google Scholar] [CrossRef]
- Qiu, R.L.; Zhao, X.; Tang, Y.T.; Yu, F.M.; Hu, P.J. Antioxidative response to Cd in a newly discovered cadmium hyperaccumulator, Arabis paniculata F. Chemosphere 2008, 74, 6–12. [Google Scholar] [CrossRef]
- Zhang, C. The Physiological and Biochemical Response of Perennial Ryegrass to Cadmium Stress and the Regulation Effect of Exogenous Calcium Treatment. Master’s Thesis, Sichuan Agricultural University, Wenchuan, China, 2018. [Google Scholar]
- Deng, G.; Li, M.; Li, H.; Yin, L.; Li, W. Exposure to cadmium causes declines in growth and photosynthesis in the endangered aquatic fern (Ceratopteris pteridoides). Aquat. Bot. 2014, 112, 23–32. [Google Scholar] [CrossRef]
- Lian, J.; Wu, J.; Zeb, A.; Zheng, S.; Ma, T.; Peng, F.; Tang, J.; Liu, W. Do polystyrene nanoplastics affect the toxicity of cadmium to wheat (Triticum aestivum L.)? Environ. Pollut. 2020, 263, 114498. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Lu, L.M.; Dai, Q.X.; Zhu, L.X.; Zheng, H.Y. Cadmium-induced Peroxidation and Production of Sulfur-related Complex in Roots of Narcissuss tazeta var. chinesis. Fujian J. Agric. Sci. 2016, 31, 585–591. [Google Scholar]
- Yang, J.R.; He, J.Q.; Zhang, G.X.; Mao, X.Q. The response of several enzyme activities in different tolerant crops to Cd stress. China Environ. Sci. 1996, 113, 117. [Google Scholar]
- Sabehat, A.; Lurie, S.; Weiss, D. Expression of small heat-shock proteins at low temperatures. A possible role in protecting against chilling injurie. Plant Physiol. 1998, 117, 651–658. [Google Scholar] [CrossRef]
- Rodríguez-Serrano, M.; Romero-Puertas, M.C.; Pazmiño, D.M.; Testillano, P.S.; Risueño, M.C.; del RiíoL, A.; Sandalio, L.M. Cellular response of pea plants to cadmium toxicity: Cross talk between reactive oxygen species, nitric oxide, and calcium. Plant Physiol. 2009, 150, 229–243. [Google Scholar] [CrossRef]
- Cai, S.-Y.; Zhang, Y.; Xu, Y.-P.; Qi, Z.-Y.; Li, M.-Q.; Ahammed, G.J.; Xia, X.-J.; Shi, K.; Zhou, Y.-H.; Reiter, R.J.; et al. HsfA1a upregulates melatonin biosynthesis to confer cadmium tolerance in tomato plant. J. Pineal Res. 2017, 62, e12387. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yu, M.; Li, H.; Wang, Y.; Lu, Z.; Zhang, Y.; Liu, M.; Qiao, G.; Wu, L.; Han, X.; et al. SaHsfA4c From Sedum alfredii Hance Enhances Cadmium Tolerance by Regulating ROS-Scavenger Activities and Heat Shock Proteins Expression. Front. Plant Sci. 2020, 11, 142. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Peng, C.; Wang, P.; Fang, S.G.; Yan, L.L.; Qiu, L.H. Identification of co-chaperone Cdc37 in Penaeus monodon: Coordination with Hsp90 can reduce cadmium stress-induced lipid peroxidation. Ecotoxicol Environ. Safe 2021, 209, 111800. [Google Scholar] [CrossRef]
- Shim, D.; Hwang, J.U.; Lee, J.; Lee, S.; Choi, Y.; An, G.; Martinoia, E.; Lee, Y. Orthologs of the class A4 heat shock transcription factor HsfA4a confer cadmium tolerance in wheat and rice. Plant Cell 2009, 21, 4031–4043. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tao, Q.; Li, J.; Guo, X.; Luo, J.; Jupa, R.; Liang, Y.; Li, T. Ethylene-mediated apoplastic barriers development involved in cadmium accumulation in root of hyperaccumulator Sedum alfredii. J. Hazard. Mater. 2021, 403, 123729. [Google Scholar] [CrossRef] [PubMed]
- Schellingen, K.; Van Der Straeten, D.; Vandenbussche, F.; Prinsen, E.; Remans, T.; Vangronsveld, J.; Cuypers, A. Cadmium-induced ethylene production and responses in Arabidopsis thaliana rely on ACS2 and ACS6 gene expression. BMC Plant Biol. 2014, 14, 214. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Li, C.; Zhang, F.; Yu, Q.; Gao, S.; Zhang, M.; Tian, H.; Zhang, J.; Yuan, X.; Ding, Z. Ethylene promotes cadmium-induced root growth inhibition through EIN3 controlled XTH33 and LSU1 expression in Arabidopsis. Plant Cell Environ. 2018, 41, 2449–2462. [Google Scholar] [CrossRef]
- Fuhrer, J. Ethylene Biosynthesis and Cadmium Toxicity in Leaf Tissue of Beans (Phaseolus vulgaris L.). Plant Physiol. 1982, 70, 162–167. [Google Scholar] [CrossRef]
- Wang, Y.; Yuan, M.; Li, Z.; Niu, Y.; Jin, Q.; Zhu, B.; Xu, Y. Effects of ethylene biosynthesis and signaling on oxidative stress and antioxidant defense system in Nelumbo nucifera G. under cadmium exposure. Environ. Sci. Pollut. Res. 2020, 27, 40156–40170. [Google Scholar] [CrossRef]
- Abozeid, A.; Ying, Z.; Lin, Y.; Liu, J.; Zhang, Z.; Tang, Z. Ethylene Improves Root System Development under Cadmium Stress by Modulating Superoxide Anion Concentration in Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 253. [Google Scholar] [CrossRef]
- Jia, X.; Zhao, Y.H.; Liu, T.; He, Y.H. Leaf defense system of Robinia pseudoacacia L. seedlings exposed to 3years of elevated atmospheric CO (2) and Cd-contaminated soils. SCI Total Environ. 2017, 605–606, 48–57. [Google Scholar] [CrossRef]
- Li, X.; Zhang, X.; Wu, Y.; Li, B.; Yang, Y. Physiological and biochemical analysis of mechanisms underlying cadmium tolerance and accumulation in turnip. Plant Divers. 2018, 40, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Ahmad, A. Proline enhances antioxidative enzyme activity, photosynthesis and yield of Cicer arietinum L. exposed to cadmium stress. Acta Bot. Croat. 2013, 72, 323–335. [Google Scholar] [CrossRef]
- Zouari, M.; Ben Ahmed, C.; Zorrig, W.; Elloumi, N.; Rabhi, M.; Delmail, D.; Ben Rouina, B.; Labrousse, P.; Ben Abdallah, F. Exogenous proline mediates alleviation of cadmium stress by promoting photosynthetic activity, water status and antioxidative enzymes activities of young date palm (Phoenix dactylifera L.). Ecotoxicol. Environ. Saf. 2016, 128, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Zouari, M.; Ben Ahmed, C.; Elloumi, N.; Bellassoued, K.; Delmail, D.; Labrousse, P.; Ben Abdallah, F.; Ben Rouina, B. Impact of proline application on cadmium accumulation, mineral nutrition and enzymatic antioxidant defense system of Olea europaea L. cv Chemlali exposed to cadmium stress. Ecotoxicol. Environ. Saf. 2016, 128, 195–205. [Google Scholar] [CrossRef]
- Ali, N.; Hadi, F. Phytoremediation of cadmium improved with the high production of endogenous phenolics and free proline contents in Parthenium hysterophorus plant treated exogenously with plant growth regulator and chelating agent. Environ. Sci. Pollut. Res. 2015, 22, 13305–13318. [Google Scholar] [CrossRef]
- Zhou, J.; Cheng, K.; Huang, G.; Chen, G.; Zhou, S.; Huang, Y.; Zhang, J.; Duan, H.; Fan, H. Effects of exogenous 3-indoleacetic acid and cadmium stress on the physiological and biochemical characteristics of Cinnamomum camphora. Ecotoxicol. Environ. Saf. 2020, 191, 109998. [Google Scholar] [CrossRef]
- Rady, M.M.; Hemida, K.A. Modulation of cadmium toxicity and enhancing cadmium-tolerance in wheat seedlings by exogenous application of polyamines. Ecotoxicol Environ. Safe 2015, 119, 178–185. [Google Scholar] [CrossRef]
- Alyemeni, M.N.; Ahanger, M.A.; Wijaya, L.; Alam, P.; Bhardwaj, R.; Ahmad, P. Selenium mitigates cadmium-induced oxidative stress in tomato (Solanum lycopersicum L.) plants by modulating chlorophyll fluorescence, osmolyte accumulation, and antioxidant system. Protoplasma 2018, 255, 459–469. [Google Scholar] [CrossRef]
- Jiang, H.P.; Gao, B.B.; Li, W.H.; Zhu, M.; Zheng, C.F.; Zheng, Q.S.; Wang, C.H. Physiological and biochemical responses of Ulva prolifera and Ulva linza to cadmium stress. Sci. World J. 2013, 2013, 289537. [Google Scholar] [CrossRef]
- Amri, B.; Khamassi, K.; Ali, M.B.; Jaime, A.; Teixeira, S.; Leila, B.B.K. Effects of gibberellic acid on the process of organic reserve mobilization in barley grains germinated in the presence of cadmium and molybdenum. S. Afr. J. Bot. 2016, 106, 35–40. [Google Scholar] [CrossRef]
- Mihoub, A.; Chaoui, A.; El, F.E. Biochemical changes associated with cadmium and copper stress in germinating pea seeds (Pisum sativum L.). C R Biol. 2005, 328, 33–41. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | Transporter | Metal Ions | Refs |
|---|---|---|---|
| Brassica juncea | BjYSLs BjNRAMPs | Cd2+ | [74] |
| BjYSL7 | Fe2+, Cd2+, Ni2+ | [75] | |
| BjHMA4 | Cd2+ | [85] | |
| Arabidopsis | AtHMA4 | Zn2+, Cd2+ | [88] |
| HMA2, HMA4 | Cd2+ | [88] | |
| ZIP1, ZIP2, ZIP3, ZIP4 | Zn2+ | [63] | |
| Arabidopsis halleri | AhZIP6 | Zn2+, Cd2+ | [96] |
| Oryza sativa L. | OsHMA1, OsHMA 2, OsHMA3 | Zn2+, Cd2+ | [64,86,97,98] |
| OsNRAMP1, OsNRAMP5 | Cd2+, As3+, Mn2+ | [63,95,97] | |
| OsZIP1, OsZIP3 | Cu2+, Zn2+, Cd2+ | [99,100] | |
| Thlaspi caerulescens | TcNRAMP3 | Fe3+, Cd2+ | [93] |
| ZNT1, ZNT5 ZIP | Zn2+, Cd2+ | [101] | |
| Sedum alfredii | SaNRAMP6 | Cd2+ | [91] |
| Sedum plumbizincicola | SpHMA1, SpHMA3 | Cd2+ | [35,102] |
| Brassica campestris ssp. chinensis | HMA2, HMA4, BcGSTU | Cd2+ | [81,103] |
| BcNRAMP5 | Cd2+ | [104] | |
| BcIRT1, BcZIP2 | Cd2+, Mn2+, Zn2+, Fe2+ | [105] | |
| Hordeum vulgare | HvNRAMP5 | Mn2+, Cd2+ | [94] |
| HvYS1 | Fe3+ | [106] | |
| cucumber | CsHMA3, CsHMA4 | Pb2+, Zn2+, Cd2+ | [89] |
| Solanum nigrum | SnYSL3 | Fe2+, Cu2+, Zn2+, Cd2+ | [76] |
| Populus tomentosa Carr. | PtoHMA5 | Cd2+ | [84] |
| Brassica napus L. | BnaHMA4c | Cd2+ | [107] |
| Vicia sativa | VsRIT1 | Cd2+, Fe(EDTA-Fe), Zn2+ | [80] |
| Triticum turgidum L. ssp. turgidum | TtNRAMP6 | Cd2+ | [92] |
| Triticum polonicum L. | TpNRAMP5 | Cd2+, Co2+, Mn2+ | [108] |
| Nicotiana tabacum | NtZIP1 | Cd2+, Zn2+ | [50] |
| NtNRAMP5 | Mn2+, Cd2+ | [109] |
| Plant | Distribution (mg kg−1, DW) | SD (%) | BCF | TF | Refs | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Root | Leaf | Stem | Shoot | CW | OE | SF | ||||
| Ceratopteris pteridoides | 1316.34 | 191.38 | 186.19 | 28–69 | 14–44 | 6–46 | 27.99–570.75 | 0.10–0.14 | [16] | |
| Lantana camara L. | 293.4 | 423.06 | 392.37 | 301.78 | 1.32–3.14 | 1.04–1.41 | [29] | |||
| Myriophyllum aquaticum | 122.530 | 111.828 | 24.92–38.57 | 0.97–12.04 | 57.40–66.25 | [12] | ||||
| Calendula calypso | 165 | 78 | 2.7–4.14 | 0.46–0.50 | [17] | |||||
| Koelreuteria paniculata | 8.48 | 4.04 | 2.31 | 45–77 | 2–11 | 20–45 | 0.052–0.318 | 0.464–0.705 | [116] | |
| Echinodorus Osiris Rataj | 2742.95 | 502.97 | 8.44–25.62 | −22.07 | 69.49–88.39 | >1 | 0.18 | [11] | ||
| Raphanus sativus L. | 3.52–4.94 | 4.66–6.86 | 18–49 | 15–20 | 36–51 | 1.64–2.36 | 1.32–1.38 | [13] | ||
| Morus alba L. | 31.6 | 8.57 | 15 | 31–77 | 3–13 | 16–66 | 0.10–0.35 | 0.12–0.27 | [117] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, G.-L.; Zheng, M.-M.; Tan, A.-J.; Liu, Y.-T.; Feng, D.; Lv, S.-M. Research on the Mechanisms of Plant Enrichment and Detoxification of Cadmium. Biology 2021, 10, 544. https://doi.org/10.3390/biology10060544
Yang G-L, Zheng M-M, Tan A-J, Liu Y-T, Feng D, Lv S-M. Research on the Mechanisms of Plant Enrichment and Detoxification of Cadmium. Biology. 2021; 10(6):544. https://doi.org/10.3390/biology10060544
Chicago/Turabian StyleYang, Gui-Li, Meng-Meng Zheng, Ai-Juan Tan, Yu-Ting Liu, Dan Feng, and Shi-Ming Lv. 2021. "Research on the Mechanisms of Plant Enrichment and Detoxification of Cadmium" Biology 10, no. 6: 544. https://doi.org/10.3390/biology10060544
APA StyleYang, G.-L., Zheng, M.-M., Tan, A.-J., Liu, Y.-T., Feng, D., & Lv, S.-M. (2021). Research on the Mechanisms of Plant Enrichment and Detoxification of Cadmium. Biology, 10(6), 544. https://doi.org/10.3390/biology10060544