
Bicarbonate Evokes Reciprocal Changes in Intracellular Cyclic di-GMP and Cyclic AMP Levels in Pseudomonas aeruginosa

 ,
,  , , ,
, , ,  , ,
, ,  , , and
, , and 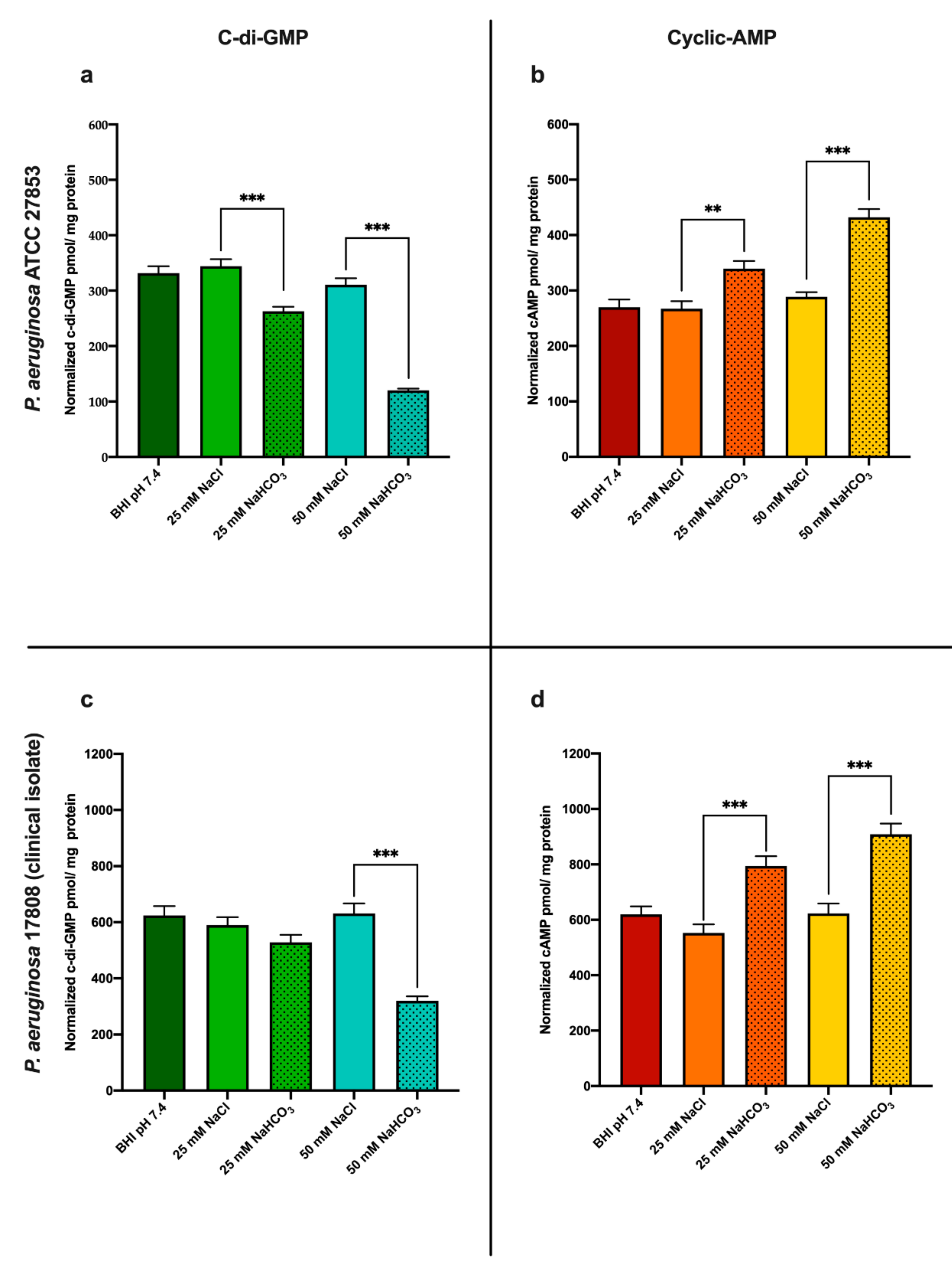{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Growth Conditions and Bacterial Strains
2.2. Extraction of c-di-GMP and cAMP
2.3. Quantification of c-di-GMP and cAMP
2.4. Real-Time Biofilm Monitoring
2.5. Biofilm Assessment Using Crystal Violet Assay
2.6. Statistical Analysis
3. Results
3.1. Sodium Bicarbonate Modulates Both c-di-GMP and cAMP Levels in P. aeruginosa
3.1.1. Sodium Bicarbonate Decreases Intracellular c-di-GMP Levels
3.1.2. Sodium Bicarbonate Increases Intracellular cAMP Levels
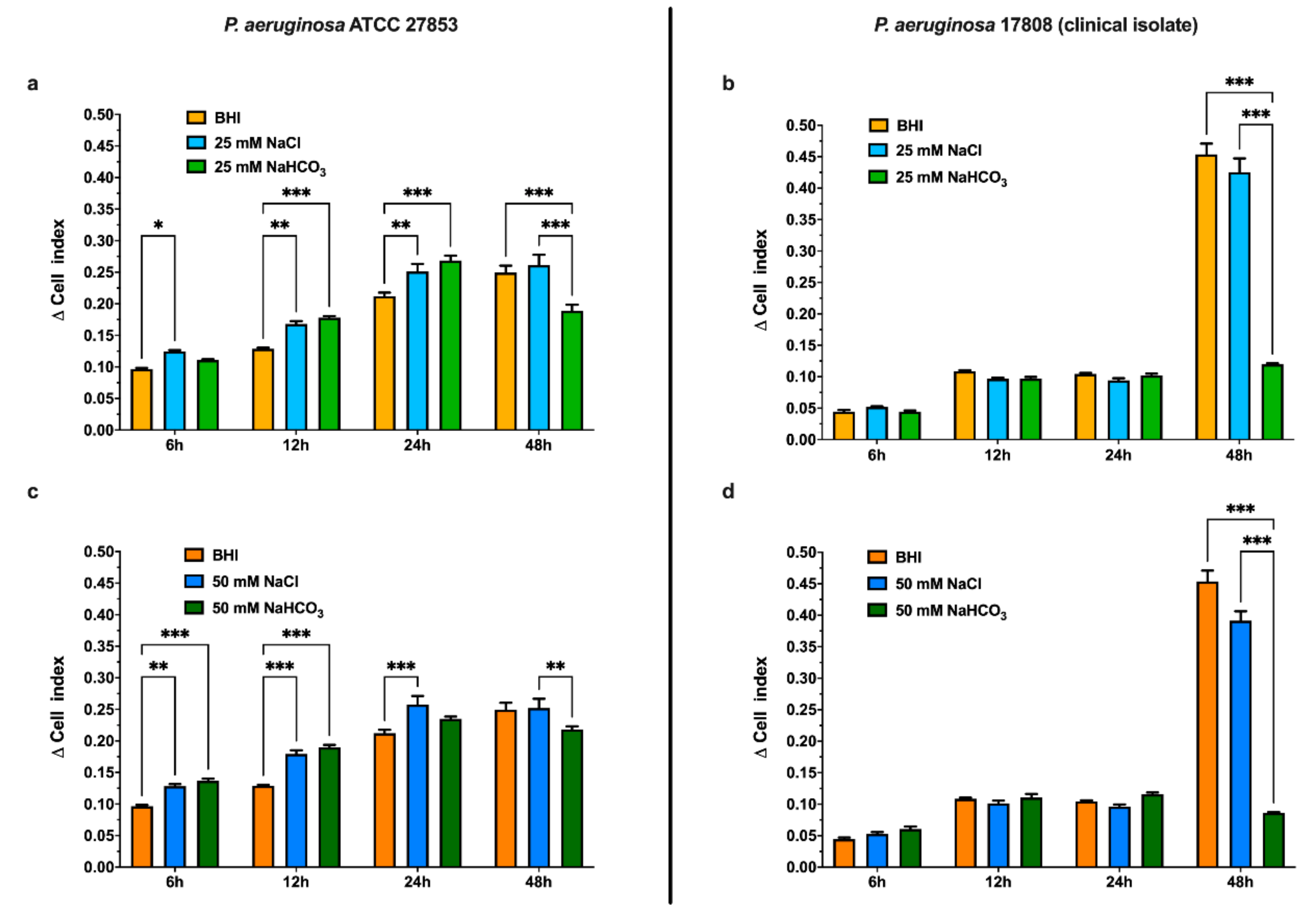
3.2. Sodium Bicarbonate Inhibits P. aeruginosa Biofilm Formation
3.2.1. Effects of Sodium Bicarbonate Assessed by Real-Time Cell Analysis (RTCA)
3.2.2. Effects of Sodium Bicarbonate Assessed by Crystal Violet Assay
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Obritsch, M.D.; Fish, D.N.; MacLaren, R.; Jung, R. Nosocomial infections due to multidrug-resistant Pseudomonas aeruginosa: Epidemiology and treatment options. Pharmacotherapy 2005, 25, 1353–1364. [Google Scholar] [CrossRef]
- Valentini, M.; Filloux, A. Biofilms and Cyclic di-GMP (c-di-GMP) signaling: Lessons from Pseudomonas aeruginosa and other bacteria. J. Biol. Chem. 2016, 291, 12547–12555. [Google Scholar] [CrossRef]
- McDonough, K.A.; Rodriguez, A. The myriad roles of cyclic AMP in microbial pathogens: From signal to sword. Nat. Rev. Microbiol. 2012, 10, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.L.; Lee, V.T. Cyclic-di-GMP regulation of virulence in bacterial pathogens. Wiley Interdiscip. Rev. RNA 2018, 9, e1454. [Google Scholar] [CrossRef]
- Almblad, H.; Harrison, J.J.; Rybtke, M.; Groizeleau, J.; Givskov, M.; Parsek, M.R.; Tolker-Nielsen, T. The cyclic AMP-Vfr signaling pathway in Pseudomonas aeruginosa is inhibited by cyclic Di-GMP. J. Bacteriol. 2015, 197, 2190–2200. [Google Scholar] [CrossRef]
- Ha, D.-G.; O’Toole, G.A. c-di-GMP and its Effects on Biofilm Formation and Dispersion: A Pseudomonas Aeruginosa Review. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef]
- Srivastava, D.; Waters, C.M. A tangled web: Regulatory connections between quorum sensing and cyclic Di-GMP. J. Bacteriol. 2012, 194, 4485–4493. [Google Scholar] [CrossRef] [PubMed]
- Barraud, N.; Schleheck, D.; Klebensberger, J.; Webb, J.S.; Hassett, D.J.; Rice, S.A.; Kjelleberg, S. Nitric oxide signaling in Pseudomonas aeruginosa biofilms mediates phosphodiesterase activity, decreased cyclic di-GMP levels, and enhanced dispersal. J. Bacteriol. 2009, 191, 7333–7342. [Google Scholar] [CrossRef] [PubMed]
- Dobay, O.; Laub, K.; Stercz, B.; Kéri, A.; Balázs, B.; Tóthpál, A.; Kardos, S.; Jaikumpun, P.; Ruksakiet, K.; Quinton, P.M.; et al. Bicarbonate Inhibits Bacterial Growth and Biofilm Formation of Prevalent Cystic Fibrosis Pathogens. Front. Microbiol. 2018, 9, 2245. [Google Scholar] [CrossRef] [PubMed]
- Dorschner, R.A.; Lopez-Garcia, B.; Peschel, A.; Kraus, D.; Morikawa, K.; Nizet, V.; Gallo, R.L. The mammalian ionic environment dictates microbial susceptibility to antimicrobial defense peptides. FASEB J. 2006, 20, 35–42. [Google Scholar] [CrossRef]
- Farha, M.A.; French, S.; Stokes, J.M.; Brown, E.D. Bicarbonate Alters Bacterial Susceptibility to Antibiotics by Targeting the Proton Motive Force. ACS Infect. Dis. 2018, 4, 382–390. [Google Scholar] [CrossRef]
- Pratten, J.; Wiecek, J.; Mordan, N.; Lomax, A.; Patel, N.; Spratt, D.; Middleton, A. Physical disruption of oral biofilms by sodium bicarbonate: An in vitro study. Int. J. Dent. Hyg. 2016, 14, 209–214. [Google Scholar] [CrossRef]
- Jaikumpun, P.; Ruksakiet, K.; Stercz, B.; Pállinger, É.; Steward, M.; Lohinai, Z.; Dobay, O.; Zsembery, Á. Antibacterial Effects of Bicarbonate in Media Modified to Mimic Cystic Fibrosis Sputum. Int. J. Mol. Sci. 2020, 21, 8614. [Google Scholar] [CrossRef] [PubMed]
- Gawande, P.V.; LoVetri, K.; Yakandawala, N.; Romeo, T.; Zhanel, G.G.; Cvitkovitch, D.G.; Madhyastha, S. Antibiofilm activity of sodium bicarbonate, sodium metaperiodate and SDS combination against dental unit waterline-associated bacteria and yeast. J. Appl. Microbiol. 2008, 105, 986–992. [Google Scholar] [CrossRef]
- Malik, Y.S.; Goyal, S.M. Virucidal efficacy of sodium bicarbonate on a food contact surface against feline calicivirus, a norovirus surrogate. Int. J. Food Microbiol. 2006, 109, 160–163. [Google Scholar] [CrossRef]
- Ratjen, F.; Bell, S.C.; Rowe, S.M.; Goss, C.H.; Quittner, A.L.; Bush, A. Cystic fibrosis. Nat. Rev. Dis. Prim. 2015, 1, 15010. [Google Scholar] [CrossRef] [PubMed]
- Quinton, P.M. Role of epithelial HCO3- transport in mucin secretion: Lessons from cystic fibrosis. Am. J. Physiol. Cell Physiol. 2010, 299, C1222. [Google Scholar] [CrossRef]
- Kunzelmann, K.; Schreiber, R.; Hadorn, H.B. Bicarbonate in cystic fibrosis. J. Cyst. Fibros. 2017, 16, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Quinton, P.M. Cystic fibrosis: Impaired bicarbonate secretion and mucoviscidosis. Lancet 2008, 372, 415–417. [Google Scholar] [CrossRef]
- Haq, I.J.; Gray, M.A.; Garnett, J.P.; Ward, C.; Brodlie, M. Airway surface liquid homeostasis in cystic fibrosis: Pathophysiology and therapeutic targets. Thorax 2016, 71, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Bhagirath, A.Y.; Li, Y.; Somayajula, D.; Dadashi, M.; Badr, S.; Duan, K. Cystic fibrosis lung environment and Pseudomonas aeruginosa infection. BMC Pulm. Med. 2016, 16, 174. [Google Scholar] [CrossRef]
- Gomez, C.C.S.; Parazzi, P.L.F.; Clinckspoor, K.J.; Mauch, R.M.; Pessine, F.B.T.; Levy, C.E.; Peixoto, A.O.; Ribeiro, M.Â.G.O.; Ribeiro, A.F.; Conrad, D.; et al. Safety, Tolerability, and Effects of Sodium Bicarbonate Inhalation in Cystic Fibrosis. Clin. Drug Investig. 2020, 40, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Almblad, H.; Rybtke, M.; Hendiani, S.; Andersen, J.B.; Givskov, M.; Tolker-Nielsen, T. High levels of cAMP inhibit Pseudomonas aeruginosa biofilm formation through reduction of the c-di-GMP content. Microbiology 2019, 165, 324–333. [Google Scholar] [CrossRef]
- Opoku-Temeng, C.; Zhou, J.; Zheng, Y.; Su, J.; Sintim, H.O. Cyclic dinucleotide (c-di-GMP, c-di-AMP, and cGAMP) signalings have come of age to be inhibited by small molecules. Chem. Commun. 2016, 52, 9327–9342. [Google Scholar] [CrossRef]
- Sambanthamoorthy, K.; Sloup, R.E.; Parashar, V.; Smith, J.M.; Kim, E.E.; Semmelhack, M.F.; Neiditch, M.B.; Waters, C.M. Identification of small molecules that antagonize diguanylate cyclase enzymes to inhibit biofilm formation. Antimicrob. Agents Chemother. 2012, 56, 5202–5211. [Google Scholar] [CrossRef]
- Kimura, Y.; Okazaki, N.; Takegawa, K. Enzymatic characteristics of two novel Myxococcus xanthus enzymes, PdeA and PdeB, displaying 3′,5′- and 2′,3′-cAMP phosphodiesterase, and phosphatase activities. FEBS Lett. 2009, 583, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Topal, H.; Fulcher, N.B.; Bitterman, J.; Salazar, E.; Buck, J.; Levin, L.R.; Cann, M.J.; Wolfgang, M.C.; Steegborn, C. Crystal structure and regulation mechanisms of the CyaB adenylyl cyclase from the human pathogen pseudomonas aeruginosa. J. Mol. Biol. 2012, 416, 271–286. [Google Scholar] [CrossRef]
- Petrova, O.E.; Sauer, K. High-performance liquid chromatography (HPLC)-based detection and quantitation of cellular c-di-GMP. Methods Mol. Biol. 2017, 1657, 33–43. [Google Scholar] [CrossRef]
- Burhenne, H.; Kaever, V. Quantification of cyclic dinucleotides by reversed-phase LC-MS/MS. In Cyclic Nucleotide Signaling in Plants Methods and Protocols; Gehring, C., Ed.; Humana Press Inc.: Tortowa, NJ, USA, 2013; Volume 1016, pp. 27–37. ISBN 9781627034401. [Google Scholar]
- Shukla, S.K.; Rao, T.S. An Improved Crystal Violet Assay for Biofilm Quantification in 96-Well Microtitre Plate. bioRxiv 2017. [Google Scholar] [CrossRef]
- Wenderska, I.B.; Chong, M.; McNulty, J.; Wright, G.D.; Burrows, L.L. Palmitoyl-dl-Carnitine is a multitarget inhibitor of Pseudomonas aeruginosa biofilm development. ChemBioChem 2011, 12, 2759–2766. [Google Scholar] [CrossRef] [PubMed]
- Jenal, U.; Reinders, A.; Lori, C. Cyclic di-GMP: Second messenger extraordinaire. Nat. Rev. Microbiol. 2017, 15, 271–284. [Google Scholar] [CrossRef]
- Koestler, B.J.; Waters, C.M. Intestinal GPS: Bile and bicarbonate control cyclic di-GMP to provide Vibrio cholerae spatial cues within the small intestine. Gut Microbes 2014, 5, 775–780. [Google Scholar] [CrossRef]
- Evans, T.J. Small colony variants of Pseudomonas aeruginosa in chronic bacterial infection of the lung in cystic fibrosis. Future Microbiol. 2015, 10, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Rohde, M. Small-Colony Variants of Pseudomonas aeruginosa in Cystic Fibrosis. Clin. Infect. Dis. 1999, 29, 621–625. [Google Scholar] [CrossRef]
- Blanka, A.; Düvel, J.; Dötsch, A.; Klinkert, B.; Abraham, W.R.; Kaever, V.; Ritter, C.; Narberhaus, F.; Häussler, S. Constitutive production of c-di-GMP is associated with mutations in a variant of Pseudomonas aeruginosa with altered membrane composition. Sci. Signal. 2015, 8, ra36. [Google Scholar] [CrossRef]
- Malone, J.G.; Jaeger, T.; Spangler, C.; Ritz, D.; Spang, A.; Arrieumerlou, C.; Kaever, V.; Landmann, R.; Jenal, U. YfiBNR Mediates Cyclic di-GMP Dependent Small Colony Variant Formation and Persistence in Pseudomonas aeruginosa. PLoS Pathog. 2010, 6, e1000804. [Google Scholar] [CrossRef]
- Malone, J.G. Role of small colony variants in persistence of Pseudomonas aeruginosa infections in cystic fibrosis lungs. Infect. Drug Resist. 2015, 8, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cann, M.J.; Litvin, T.N.; Iourgenko, V.; Sinclair, M.L.; Levin, L.R.; Buck, J. Soluble adenylyl cyclase as an evolutionarily conserved bicarbonate sensor. Science 2000, 289, 625–628. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Buck, J.; Levin, L.R. Conservation of functional domain structure in bicarbonate-regulated “soluble” adenylyl cyclases in bacteria and eukaryotes. Dev. Genes Evol. 2004, 214, 503–509. [Google Scholar] [CrossRef]
- Ono, K.; Oka, R.; Toyofuku, M.; Sakaguchi, A.; Hamada, M.; Yoshida, S.; Nomura, N. CAMP signaling affects irreversible attachment during biofilm formation by pseudomonas aeruginosa PAo1. Microbes Environ. 2014, 29, 104–106. [Google Scholar] [CrossRef]
- Gutiérrez, D.; Hidalgo-Cantabrana, C.; Rodríguez, A.; García, P.; Ruas-Madiedo, P. Monitoring in Real Time the Formation and Removal of Biofilms from Clinical Related Pathogens Using an Impedance-Based Technology. PLoS ONE 2016, 11, e0163966. [Google Scholar] [CrossRef]
- Junka, A.F.; Janczura, A.; Smutnicka, D.; Mączyńska, B.; Secewicz, A.; Nowicka, J.; Bartoszewicz, M.; Gościniak, G. Use of the real time xCelligence system for purposes of medical microbiology. Polish J. Microbiol. 2012, 61, 191–197. [Google Scholar] [CrossRef]
- Ferrer, M.D.; Rodriguez, J.C.; Álvarez, L.; Artacho, A.; Royo, G.; Mira, A. Effect of antibiotics on biofilm inhibition and induction measured by real-time cell analysis. J. Appl. Microbiol. 2017, 122, 640–650. [Google Scholar] [CrossRef]
- Kho, D.; MacDonald, C.; Johnson, R.; Unsworth, C.; O’Carroll, S.; Mez, E.; Angel, C.; Graham, E. Application of xCELLigence RTCA Biosensor Technology for Revealing the Profile and Window of Drug Responsiveness in Real Time. Biosensors 2015, 5, 199–222. [Google Scholar] [CrossRef]
- Cao, H.; Lai, Y.; Bougouffa, S.; Xu, Z.; Yan, A. Comparative genome and transcriptome analysis reveals distinctive surface characteristics and unique physiological potentials of Pseudomonas aeruginosa ATCC 27853. BMC Genom. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Perez, L.R.R.; Costa, M.C.N.; Freitas, A.L.P.; Barth, A.L. Evaluation of biofilm production by pseudomonas aeruginosa isolates recovered fromcystic fibrosis and non-cystic fibrosis patients. Braz. J. Microbiol. 2011, 42, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Alhede, M.; Kragh, K.N.; Qvortrup, K.; Allesen-Holm, M.; van Gennip, M.; Christensen, L.D.; Jensen, P.Ø.; Nielsen, A.K.; Parsek, M.; Wozniak, D.; et al. Phenotypes of non-attached pseudomonas aeruginosa aggregates resemble surface attached biofilm. PLoS ONE 2011, 6, 27943. [Google Scholar] [CrossRef] [PubMed]
- Caiazza, N.C.; O’Toole, G.A. SadB is required for the transition from reversible to irreversible attachment during biofilm formation by Pseudomonas aeruginosa PA14. J. Bacteriol. 2004, 186, 4476–4485. [Google Scholar] [CrossRef]
- Friedman, L.; Kolter, R. Genes involved in matrix formation in Pseudomonas aeruginosa PA14 biofilms. Mol. Microbiol. 2004, 51, 675–690. [Google Scholar] [CrossRef]
- Peeters, E.; Nelis, H.J.; Coenye, T. Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 2008, 72, 157–165. [Google Scholar] [CrossRef]
- Stigliani, M.; Manniello, M.D.; Zegarra-Moran, O.; Galietta, L.; Minicucci, L.; Casciaro, R.; Garofalo, E.; Incarnato, L.; Aquino, R.P.; Del Gaudio, P.; et al. Rheological Properties of Cystic Fibrosis Bronchial Secretion and In Vitro Drug Permeation Study: The Effect of Sodium Bicarbonate. J. Aerosol Med. Pulm. Drug Deliv. 2016, 29, 337–345. [Google Scholar] [CrossRef]
- Ferrera, L.; Capurro, V.; Delpiano, L.; Gianotti, A.; Moran, O. The Application of Bicarbonate Recovers the Chemical-Physical Properties of Airway Surface Liquid in Cystic Fibrosis Epithelia Models. Biology 2021, 10, 278. [Google Scholar] [CrossRef] [PubMed]
- Gróf, I.; Bocsik, A.; Harazin, A.; Santa-Maria, A.R.; Vizsnyiczai, G.; Barna, L.; Kiss, L.; Fűr, G.; Rakonczay, Z.; Ambrus, R.; et al. The effect of sodium bicarbonate, a beneficial adjuvant molecule in cystic fibrosis, on bronchial epithelial cells expressing a wild-type or mutant CFTR channel. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Kis, A.; Toth, L.A.; Kunos, L.; Vasas, S.; Losonczy, G.; Mendes, E.; Wanner, A.; Horvath, G. The effect of airway alkalization by nebulized sodium bicarbonate on airway blood flow. Eur. Respir. J. 2012, 40, 2143. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruksakiet, K.; Stercz, B.; Tóth, G.; Jaikumpun, P.; Gróf, I.; Tengölics, R.; Lohinai, Z.M.; Horváth, P.; Deli, M.A.; Steward, M.C.; et al. Bicarbonate Evokes Reciprocal Changes in Intracellular Cyclic di-GMP and Cyclic AMP Levels in Pseudomonas aeruginosa. Biology 2021, 10, 519. https://doi.org/10.3390/biology10060519
Ruksakiet K, Stercz B, Tóth G, Jaikumpun P, Gróf I, Tengölics R, Lohinai ZM, Horváth P, Deli MA, Steward MC, et al. Bicarbonate Evokes Reciprocal Changes in Intracellular Cyclic di-GMP and Cyclic AMP Levels in Pseudomonas aeruginosa. Biology. 2021; 10(6):519. https://doi.org/10.3390/biology10060519
Chicago/Turabian StyleRuksakiet, Kasidid, Balázs Stercz, Gergő Tóth, Pongsiri Jaikumpun, Ilona Gróf, Roland Tengölics, Zsolt M. Lohinai, Péter Horváth, Mária A. Deli, Martin C. Steward, and et al. 2021. "Bicarbonate Evokes Reciprocal Changes in Intracellular Cyclic di-GMP and Cyclic AMP Levels in Pseudomonas aeruginosa" Biology 10, no. 6: 519. https://doi.org/10.3390/biology10060519
APA StyleRuksakiet, K., Stercz, B., Tóth, G., Jaikumpun, P., Gróf, I., Tengölics, R., Lohinai, Z. M., Horváth, P., Deli, M. A., Steward, M. C., Dobay, O., & Zsembery, Á. (2021). Bicarbonate Evokes Reciprocal Changes in Intracellular Cyclic di-GMP and Cyclic AMP Levels in Pseudomonas aeruginosa. Biology, 10(6), 519. https://doi.org/10.3390/biology10060519