
Identification and Functional Analysis of a Lysozyme Gene from Coridius chinensis (Hemiptera: Dinidoridae)


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Bacteria
2.2. Characteristic Analyses of CcLys2
2.3. RNA Extraction and cDNA Synthesis
2.4. Spatiotemporal Expression Profile of CcLys2
2.5. Analysis of Bacterial Infection
2.6. Construction of Recombinant Expression Vector
2.7. Expression and Purification of Recombinant Protein
2.8. Activity Assay of CcLys2
2.9. Antibacterial Assay of CcLys2
2.10. Data Analysis
3. Results
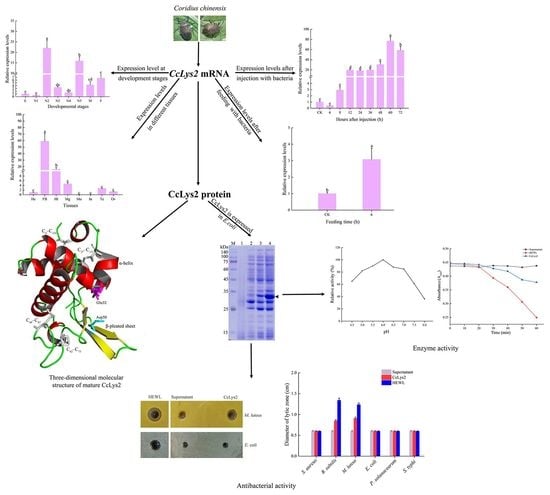
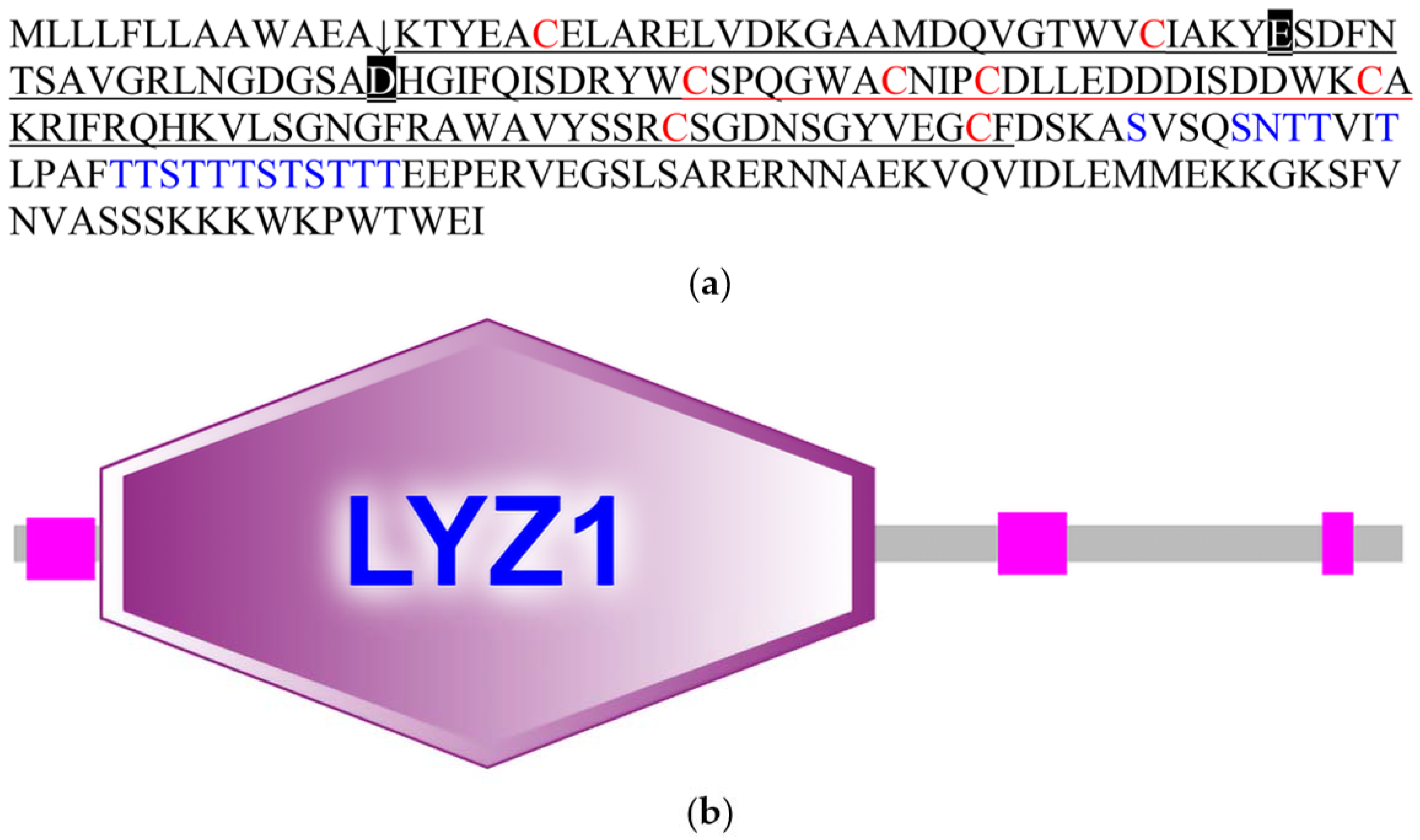
3.1. Characteristic Analyses of CcLys2
3.2. Homologous and Phylogenetic Analyses
3.3. Spatiotemporal Expression Patterns
3.4. Expression profile of CcLys2 after Bacterial Infection
3.5. Identification of Recombinant Strain
3.6. Muramidase Activity of CcLys2
3.7. Antimicrobial Activity of CcLys2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Callewaert, L.; Michiels, C. Lysozymes in the animal kingdom. J. Biosci. 2010, 35, 127–160. [Google Scholar] [CrossRef] [PubMed]
- Van Herreweghe, J.; Michiels, C. Invertebrate lysozymes: Diversity and distribution, molecular mechanism and in vivo function. J. Biosci. 2012, 37, 327–348. [Google Scholar] [CrossRef] [PubMed]
- Bachali, S.; Jager, M.; Hassanin, A.; Schoentgen, F.; Jollès, P.; Fiala-Medioni, A.; Deutsch, J. Phylogenetic Analysis of Invertebrate Lysozymes and the Evolution of Lysozyme Function. J. Mol. Evol. 2002, 54, 652–664. [Google Scholar] [CrossRef] [PubMed]
- Daffre, S.; Kylsten, P.; Samakovlis, C.; Hultmark, D. The lysozyme locus in Drosophila melanogaster: An expanded gene family adapted for expression in the digestive tract. Mol. Gen. Genet. MGG 1994, 242, 152–162. [Google Scholar] [CrossRef]
- Waterhouse, R.M.; Kriventseva, E.V.; Meister, S.; Xi, Z.; Alvarez, K.S.; Bartholomay, L.C.; Barillas-Mury, C.; Bian, G.; Blandin, S.; Christensen, B.M.; et al. Evolutionary dynamics of immune-related genes and pathways in disease-vector mosquitoes. Science 2007, 316, 1738–1743. [Google Scholar] [CrossRef] [PubMed]
- Regel, R.; Matioli, S.R.; Terra, W.R. Molecular adaptation of Drosophila melanogaster lysozymes to a digestive function. Insect Biochem. Mol. Biol. 1998, 28, 309–319. [Google Scholar] [CrossRef]
- Li, B.; Calvo, E.; Marinotti, O.; James, A.A.; Paskewitz, S.M. Characterization of the c-type lysozyme gene family in Anopheles gambiae. Gene 2005, 360, 131–139. [Google Scholar] [CrossRef]
- Wiesner, J.; Vilcinskas, A. Antimicrobial peptides the ancient arm of the human immune system. Virulence 2010, 1, 440–464. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, J. Innate Immunity of Insects. Curr. Opin. Immunol. 1995, 7, 4–10. [Google Scholar] [CrossRef]
- Cytryńska, M.; Wojda, I.; Jakubowicz, T. How Insects Combat Infections. In Lessons in Immunity; Ballarin, L., Cammarata, M., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 9, pp. 117–128. [Google Scholar] [CrossRef]
- Imoto, T.; Johnson, L.N.; North, A.C.T.; Phillips, D.C.; Rupley, J. 21 Vertebrate Lysozymes. In The Enzymes; Academic Press: Cambridge, MA, USA, 1972; Volume 7, pp. 665–868. [Google Scholar] [CrossRef]
- Beckert, A.; Wiesner, J.; Schmidtberg, H.; Lehmann, R.; Baumann, A.; Vogel, H.; Vilcinskas, A. Expression and characterization of a recombinant i-type lysozyme from the harlequin ladybird beetle Harmonia axyridis. Insect Mol. Biol. 2016, 25, 202–215. [Google Scholar] [CrossRef] [PubMed]
- Cançado, F.C.; Valério, A.A.; Marana, S.R.; Barbosa, J.A.R.G. The crystal structure of a lysozyme c from housefly Musca domestica, the first structure of a digestive lysozyme. J. Struct. Biol. 2007, 160, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Marana, S.R.; Cançado, F.C.; Valério, A.A.; Ferreira, C.; Terra, W.R.; Barbosa, J.A.R.G. Crystallization, data collection and phasing of two digestive lysozymes from Musca domestica. Acta Cryst. Sect. F Struct. Biol. Cryst. Commun. 2006, 62, 750–752. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-J.; Brey, P.T. Isolation and characterization of the lysozyme-encoding gene from the silkworm Bombyx mori. Gene 1995, 161, 199–203. [Google Scholar] [CrossRef]
- Parry, R.M.; Chandan, R.C.; Shahani, K.M. Isolation and characterization of human milk lysozyme. Arch. Biochem. Biophys. 1969, 130, 59–65. [Google Scholar] [CrossRef]
- Gandhe, A.; Janardhan, G.; Nagaraju, J. Immune upregulation of novel antibacterial proteins from silkmoth (Lepidoptera) that resemble lysozymes but lack muramidase activity. Insect Biochem. Mol. Biol. 2007, 37, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Mandal, A.; Klotz, K.; Shetty, J.; Jayes, F.; Wolkowicz, M.; Bolling, L.; Coonrod, S.; Black, M.; Diekman, A.; Haystead, T.; et al. SLLP1, A Unique, Intra-acrosomal, Non-bacteriolytic, c Lysozyme-Like Protein of Human Spermatozoa. Biol. Reprod. 2003, 68, 1525–1537. [Google Scholar] [CrossRef]
- Karthik, V.; Vijayan, K.; Thomas, A.; Sudheer, N.S.; Singh, B.; Narayanan, R. Functional Characterization of a c-type Lysozyme from Indian Shrimp Fenneropenaeus indicus. Probiotics Antimicrob. Proteins 2013, 6. [Google Scholar] [CrossRef]
- Yu, L.-P.; Sun, B.-G.; Li, J.; Sun, L. Characterization of a c-type lysozyme of Scophthalmus maximus: Expression, activity, and antibacterial effect. Fish Shellfish Immunol. 2013, 34, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Shang-Wei, L.I.; Bai-Song, Z.; Juan, D.U. Isolation, purification, and detection of the antimicrobial activity of the antimicrobial peptide CcAMP1 from Coridius chinensis (Hemiptera:Dinidoridae). Acta Entomol. Sin. 2015, 58, 610–616. [Google Scholar] [CrossRef]
- Su, T.; Mulla, M.S. Quantitative Determination of Free Amino Acids in the Hemolymph of Autogenous and Anautogenous Strains of Culex tarsalis (Diptera: Culicidae). J. Med. Entomol. 1997, 34, 729–734. [Google Scholar] [CrossRef]
- Houser, M. Improved turbidimetric assay for lysozyme in urine. Clin. Chem. 1983, 29, 1488–1493. [Google Scholar] [CrossRef] [PubMed]
- Stelzner, A.; Klein, U.; Klein, Y.M.; Holtz, H. Optimization of lysozyme determination: Comparative study of preparations of test cultures of Micrococcus luteus (M. lysodeikticus Fleming). Zent. Bakteriol. Mikrobiol. Hyg. A 1982, 253, 253–264. [Google Scholar] [CrossRef]
- Malcolm, B.; Rosenberg, S.; Corey, M.; Allen, J.; Baetselier, A.; Kirsch, J. Site-directed mutagenesis of the catalytic residues Asp-52 and Glu-35 of chicken egg white lysozyme. Proc. Natl. Acad. Sci. USA 1989, 86, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Vocadlo, D.; Davies, G.; Laine, R.A.; Withers, S. Catalysis by hen egg-white lysozyme proceeds via a covalent intermediate. Nature 2001, 412, 835–838. [Google Scholar] [CrossRef] [PubMed]
- Kyomuhendo, P.; Myrnes, B.; Nilsen, I.W. A cold-active salmon goose-type lysozyme with high heat tolerance. Cell. Mol. Life Sci. CMLS 2007, 64, 2841–2847. [Google Scholar] [CrossRef] [PubMed]
- Grunclová, L.; Fouquier, H.; Hypša, V.; Kopáček, P. Lysozyme from the gut of the soft tick Ornithodoros moubata: The sequence, phylogeny and post-feeding regulation. Dev. Comp. Immunol. 2003, 27, 651–660. [Google Scholar] [CrossRef]
- Broersen, K.; Voragen, A.; Hamer, R.; Jongh, H. Glycoforms of β-Lactoglobulin with Improved Thermostability and Preserved Structural Packing. Biotechnol. Bioeng. 2004, 86, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Jafari-Aghdam, J.; Khajeh, K.; Ranjbar, B.; Nemat-Gorgani, M. Deglycosylation of glucoamylase from Aspergillus niger: Effects on structure, activity and stability. Biochim. Biophys. Acta 2005, 1750, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Solá, R.; Griebenow, K. Effects of Glycosylation on the Stability of Protein Pharmaceuticals. J. Pharm. Sci. 2009, 98, 1223–1245. [Google Scholar] [CrossRef] [PubMed]
- Arnold, J.; Wormald, M.; Sim, R.; Rudd, P.; Dwek, R. The Impact of Glycosylation on the Biological Function and Structure of Human Immunoglobulins. Annu. Rev. Immunol. 2007, 25, 21–50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Sun, C.; Sun, S.-S.; Zhao, X.-F.; Wang, J.-X. Functional analysis of two invertebrate-type lysozymes from red swamp crayfish, Procambarus clarkii. Fish Shellfish Immunol. 2010, 29, 1066–1072. [Google Scholar] [CrossRef]
- Kollien, A.H.; Fechner, S.; Waniek, P.J.; Schaub, G.A. Isolation and characterization of a cDNA encoding for a lysozyme from the gut of the reduviid bug Triatoma infestans. Arch. Insect Biochem. Physiol. 2003, 53, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Boman, H.G. Gene-Encoded Peptide Antibiotics and the Concept of Innate Immunity: An Update Review. Scand. J. Immunol. 1998, 48, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Imler, J.-L.; Bulet, P. Mechanisms of Epithelial Defense. Chem. Immunol. Allergy 2005, 86, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Kanost, M.R.; Jiang, H.; Yu, X.-Q. Innate immune responses of a lepidopteran insect, Manduca sexta. Immunol. Rev. 2004, 198, 97–105. [Google Scholar] [CrossRef]
- Dobson, D.E.; Prager, E.M.; Wilson, A.C. Stomach lysozymes of ruminants. I. Distribution and catalytic properties. J. Biol. Chem. 1984, 259, 11607–11616. [Google Scholar] [CrossRef]
- McHenery, J.; Birkbeck, T.; Allen, J. The occurrence of lysozyme in marine bivalves. Comp. Biochem. Physiol. Part B Comp. Biochem. 1979, 63, 25–28. [Google Scholar] [CrossRef]
- Lemos, F.J.A.; Ribeiro, A.F.; Terra, W.R. A bacteria-digesting midgut-lysozyme from Musca domestica (diptera) larvae. Purification, properties and secretory mechanism. Insect Biochem. Mol. Biol. 1993, 23, 533–541. [Google Scholar] [CrossRef]
- Herbert, S.; Bera, A.; Nerz, C.; Kraus, D.; Peschel, A.; Goerke, C.; Meehl, M.; Cheung, A.; Götz, F. Molecular basis of resistance to muramidase and cationic antimicrobial peptide activity of lysozyme in staphylococci. PLoS Pathog. 2007, 3, e102. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.; Kitaoka, M.; Ohmiya, K.; Hayashi, K. Substrate Specificity of the N,6-O-Diacetylmuramidase from Streptomyces globisporus. J. Biosci. Bioeng. 2003, 95, 313–316. [Google Scholar] [CrossRef]
- Callewaert, L.; Aertsen, A.; Deckers, D.; Vanoirbeek, K.G.A.; Vanderkelen, L.; Van Herreweghe, J.M.; Masschalck, B.; Nakimbugwe, D.; Robben, J.; Michiels, C.W. A New Family of Lysozyme Inhibitors Contributing to Lysozyme Tolerance in Gram-Negative Bacteria. PLoS Pathog. 2008, 4, e1000019. [Google Scholar] [CrossRef] [PubMed]
- Supungul, P.; Rimphanitchayakit, V.; Aoki, T.; Hirono, I.; Tassanakajon, A. Molecular characterization and expression analysis of a c-type and two novel muramidase-deficient i-type lysozymes from Penaeus monodon. Fish Shellfish Immunol. 2010, 28, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Hultmark, D. Insect lysozymes. EXS 1996, 75, 87–102. [Google Scholar] [CrossRef] [PubMed]
- Prager, E. Adaptive evolution of lysozyme: Changes in amino acid sequence, regulation of expression and gene number. EXS 1996, 75, 323–345. [Google Scholar] [CrossRef] [PubMed]
- Beckert, A.; Wiesner, J.; Baumann, A.; Pöppel, A.-K.; Vogel, H.; Vilcinskas, A. Two c-type lysozymes boost the innate immune system of the invasive ladybird Harmonia axyridis. Dev. Comp. Immunology 2015, 49, 303–312. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence | Primer Usage |
|---|---|---|
| T7-F | 5′-TAATACGACTCACTATAGG-3′ | Clone |
| T7-R | 5′-GCTAGTTATTGCTCAGCGG-3′ | |
| Lys-qF | 5′-CTCTTGGAGGACGACGACATCT-3′ | RT-qPCR |
| Lys-qR | 5′-TGACTGTGGTGTTGGACTGTGA-3′ | |
| Actin-F | 5′-ACCGCTGAGAGGGAAATCG-3′ | |
| Actin-R | 5′-CAAGAAGGAAGGCTGGAAGAG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, H.; Du, J.; Li, S.-W.; Gong, T. Identification and Functional Analysis of a Lysozyme Gene from Coridius chinensis (Hemiptera: Dinidoridae). Biology 2021, 10, 330. https://doi.org/10.3390/biology10040330
Huang H, Du J, Li S-W, Gong T. Identification and Functional Analysis of a Lysozyme Gene from Coridius chinensis (Hemiptera: Dinidoridae). Biology. 2021; 10(4):330. https://doi.org/10.3390/biology10040330
Chicago/Turabian StyleHuang, Hai, Juan Du, Shang-Wei Li, and Tao Gong. 2021. "Identification and Functional Analysis of a Lysozyme Gene from Coridius chinensis (Hemiptera: Dinidoridae)" Biology 10, no. 4: 330. https://doi.org/10.3390/biology10040330
APA StyleHuang, H., Du, J., Li, S.-W., & Gong, T. (2021). Identification and Functional Analysis of a Lysozyme Gene from Coridius chinensis (Hemiptera: Dinidoridae). Biology, 10(4), 330. https://doi.org/10.3390/biology10040330