
Multi-Strain Probiotics: Synergy among Isolates Enhances Biological Activities


Abstract
Simple Summary
Abstract
1. Introduction
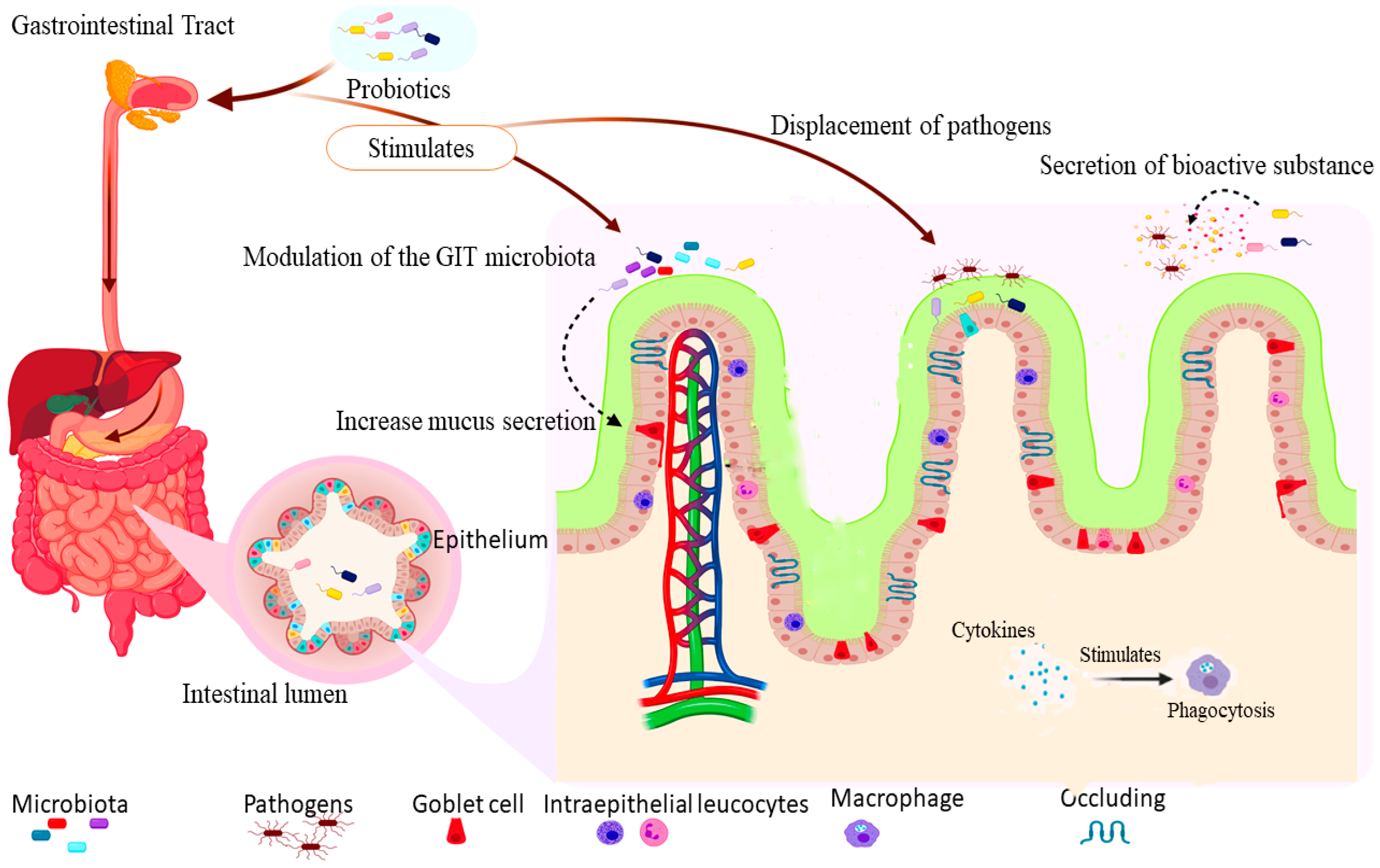
2. Mechanisms of Action of Probiotics
2.1. Stimulation of Bowel Microbiota
2.2. Immune Modulation
2.3. Stimulation of the Digestive Enzymes
2.4. Displacement of Possible Pathogens
2.5. Secretion of Bioactive Substances
2.6. Mechanisms of Action of Multi-Strain Probiotics
2.7. Antagonisms among Multi-Strain Probiotics
3. Applications and Biological Functions of Multi-Strain Probiotics
3.1. Treatment of Diseases
3.2. Inhibition of Pathogens
3.3. Improvement of Human Health
3.4. Multi-Strains Probiotics in Animal Husbandry
3.5. Poultry Farming
3.6. Aquaculture
3.7. Swine Production/Piggery
3.8. Ruminants Nutrition and Production
3.9. Synbiosis of Multi-Strain Probiotics with Other Biologically Active Molecules
4. Conclusion and Future Consideration
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506. [Google Scholar] [CrossRef]
- VidyaLaxme, B.; Rovetto, A.; Grau, R.; Agrawal, R. Synergistic effects of probiotic Leuconostoc mesenteroides and Bacillus subtilis in malted ragi (Eleucine corocana) food for antagonistic activity against V. cholerae and other beneficial properties. J. Food Sci. Technol. 2014, 51, 3072–3082. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bravo, J.A.; Julio-Pieper, M.; Forsythe, P.; Kunze, W.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Communication between gastrointestinal bacteria and the nervous system. Curr. Opin. Pharmacol. 2012, 12, 667–672. [Google Scholar] [CrossRef]
- Alagawany, M.; Abd El-Hack, M.E.; Farag, M.R.; Sachan, S.; Karthik, K.; Dhama, K. The use of probiotics as eco-friendly alternatives for antibiotics in poultry nutrition. Environ. Sci. Pollut. Res. 2018, 25, 10611–10618. [Google Scholar] [CrossRef]
- Liu, K.-F.; Chiu, C.-H.; Shiu, Y.-L.; Cheng, W.; Liu, C.-H. Effects of the probiotic, Bacillus subtilis E20, on the survival, development, stress tolerance, and immune status of white shrimp, Litopenaeus vannamei larvae. Fish Shellfish Immunol. 2010, 28, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-C.; Hu, S.-Y.; Chiu, C.-S.; Liu, C.-H. Multiple-strain probiotics appear to be more effective in improving the growth performance and health status of white shrimp, Litopenaeus vannamei, than single probiotic strains. Fish Shellfish Immunol. 2019, 84, 1050–1058. [Google Scholar] [CrossRef]
- Chaucheyras-Durand, F.; Durand, H. Probiotics in animal nutrition and health. Benef. Microbes 2010, 1, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Abudabos, A.; Al-Batshan, H.; Murshed, M. Effects of prebiotics and probiotics on the performance and bacterial colonization of broiler chickens. S. Afr. J. Anim. Sci. 2015, 45, 419–428. [Google Scholar] [CrossRef]
- Chang, C.H.; Teng, P.Y.; Lee, T.T.; Yu, B. The effects of the supplementation of multi-strain probiotics on intestinal microbiota, metabolites and inflammation of young SPF chickens challenged with Salmonella enterica subsp. enterica. Anim. Sci. J. 2019, 90, 737–746. [Google Scholar] [CrossRef]
- Nagpal, R.; Kumar, A.; Kumar, M.; Behare, P.V.; Jain, S.; Yadav, H. Probiotics, their health benefits and applications for developing healthier foods: A review. FEMS Microbiol. Lett. 2012, 334, 1–15. [Google Scholar] [CrossRef]
- Babot, J.; Argañaraz-Martínez, E.; Saavedra, L.; Apella, M.; Chaia, A.P. Compatibility and safety of five lectin-binding putative probiotic strains for the development of a multi-strain protective culture for poultry. Benef. Microbes 2018, 9, 927–935. [Google Scholar] [CrossRef]
- Cartman, S.T. Time to consider Clostridium probiotics? Future Microbiol. 2011, 6, 969–971. [Google Scholar] [CrossRef]
- Narushima, S.; Sugiura, Y.; Oshima, K.; Atarashi, K.; Hattori, M.; Suematsu, M.; Honda, K. Characterization of the 17 strains of regulatory T cell-inducing human-derived Clostridia. Gut Microbes 2014, 5, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Aalaei, M.; Khatibjoo, A.; Zaghari, M.; Taherpour, K.; Akbari Gharaei, M.; Soltani, M. Comparison of single- and multi-strain probiotics effects on broiler breeder performance, egg production, egg quality and hatchability. Br. Poult. Sci. 2018, 59, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Timmerman, H.; Koning, C.; Mulder, L.; Rombouts, F.; Beynen, A. Monostrain, multistrain and multispecies probiotics—A comparison of functionality and efficacy. Int. J. Food Microbiol. 2004, 96, 219–233. [Google Scholar] [CrossRef]
- Wilkins, T.; Sequoia, J. Probiotics for gastrointestinal conditions: A summary of the evidence. Am. Fam. Physician 2017, 96, 170–178. [Google Scholar] [PubMed]
- Chapman, C.; Gibson, G.; Todd, S.; Rowland, I. Comparative in vitro inhibition of urinary tract pathogens by single-and multi-strain probiotics. Eur. J. Nutr. 2013, 52, 1669–1677. [Google Scholar] [CrossRef]
- Adamberg, S.; Sumeri, I.; Uusna, R.; Ambalam, P.; Kondepudi, K.K.; Adamberg, K.; Wadström, T.; Ljungh, Å. Survival and synergistic growth of mixed cultures of bifidobacteria and lactobacilli combined with prebiotic oligosaccharides in a gastrointestinal tract simulator. Microb. Ecol. Health Dis. 2014, 25, 23062. [Google Scholar] [CrossRef]
- Trinder, M.; Bisanz, J.; Burton, J.; Reid, G. Probiotic lactobacilli: A potential prophylactic treatment for reducing pesticide absorption in humans and wildlife. Benef. Microbes 2015, 6, 841–847. [Google Scholar] [CrossRef]
- Astolfi, M.L.; Protano, C.; Schiavi, E.; Marconi, E.; Capobianco, D.; Massimi, L.; Ristorini, M.; Baldassarre, M.E.; Laforgia, N.; Vitali, M. A prophylactic multi-strain probiotic treatment to reduce the absorption of toxic elements: In-vitro study and biomonitoring of breast milk and infant stools. Environ. Int. 2019, 130, 104818. [Google Scholar] [CrossRef]
- Kinoshita, H. Biosorption of Heavy Metals by Lactic Acid Bacteria for Detoxification. In Lactic Acid Bacteria; Springer: Berlin/Heidelberg, Germany, 2019; pp. 145–157. [Google Scholar]
- Daisley, B.A.; Monachese, M.; Trinder, M.; Bisanz, J.E.; Chmiel, J.A.; Burton, J.P.; Reid, G. Immobilization of cadmium and lead by Lactobacillus rhamnosus GR-1 mitigates apical-to-basolateral heavy metal translocation in a Caco-2 model of the intestinal epithelium. Gut Microbes 2019, 10, 321–333. [Google Scholar] [CrossRef]
- Fredua-Agyeman, M.; Stapleton, P.; Basit, A.W.; Gaisford, S. Microcalorimetric evaluation of a multi-strain probiotic: Interspecies inhibition between probiotic strains. J. Funct. Foods 2017, 36, 357–361. [Google Scholar] [CrossRef]
- Yadav, R.; Shukla, P. An overview of advanced technologies for selection of probiotics and their expediency: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 3233–3242. [Google Scholar] [CrossRef]
- Bogucka, J.; Ribeiro, D.M.; Bogusławska-Tryk, M.; Dankowiakowska, A.; da Costa, R.P.R.; Bednarczyk, M. Microstructure of the small intestine in broiler chickens fed a diet with probiotic or synbiotic supplementation. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1785–1791. [Google Scholar] [CrossRef]
- Bajaj, B.K.; Razdan, K.; Claes, I.J.; Lebeer, S. Probiotic Attributes of the Newly Isolated Lactic Acid Bacteria from Infants’gut. J. Microbiol. Biotechnol. Food Sci. 2020, 2020, 109–115. [Google Scholar]
- Oak, S.J.; Jha, R. The effects of probiotics in lactose intolerance: A systematic review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1675–1683. [Google Scholar] [CrossRef]
- Wasilewska, E.; Zlotkowska, D.; Wroblewska, B. Yogurt starter cultures of Streptococcus thermophilus and Lactobacillus bulgaricus ameliorate symptoms and modulate the immune response in a mouse model of dextran sulfate sodium-induced colitis. J. Dairy Sci. 2019, 102, 37–53. [Google Scholar] [CrossRef] [PubMed]
- Drago, L.; Toscano, M.; Rodighiero, V.; De Vecchi, E.; Mogna, G. Cultivable and pyrosequenced fecal microflora in centenarians and young subjects. J. Clin. Gastroenterol. 2012, 46, S81–S84. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.-J.; Liu, Q.-Q.; Liao, S.; Fang, H.-H.; Yin, P.; Xie, S.-W.; Tian, L.-X.; Liu, Y.-J.; Niu, J. Effects of dietary mixed probiotics on growth, non-specific immunity, intestinal morphology and microbiota of juvenile pacific white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 90, 456–465. [Google Scholar] [CrossRef] [PubMed]
- McNulty, N.P.; Yatsunenko, T.; Hsiao, A.; Faith, J.J.; Muegge, B.D.; Goodman, A.L.; Henrissat, B.; Oozeer, R.; Cools-Portier, S.; Gobert, G. The impact of a consortium of fermented milk strains on the gut microbiome of gnotobiotic mice and monozygotic twins. Sci. Transl. Med. 2011, 3, 106ra106. [Google Scholar] [CrossRef]
- Forte, C.; Acuti, G.; Manuali, E.; Casagrande Proietti, P.; Pavone, S.; Trabalza-Marinucci, M.; Moscati, L.; Onofri, A.; Lorenzetti, C.; Franciosini, M. Effects of two different probiotics on microflora, morphology, and morphometry of gut in organic laying hens. Poult. Sci. 2016, 95, 2528–2535. [Google Scholar] [CrossRef]
- Chapman, C.; Gibson, G.; Rowland, I. Effects of single-and multi-strain probiotics on biofilm formation and in vitro adhesion to bladder cells by urinary tract pathogens. Anaerobe 2014, 27, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Foligné, B.; Parayre, S.; Cheddani, R.; Famelart, M.-H.; Madec, M.-N.; Plé, C.; Breton, J.; Dewulf, J.; Jan, G.; Deutsch, S.-M. Immunomodulation properties of multi-species fermented milks. Food Microbiol. 2016, 53, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Wróblewska, B.; Kaliszewska, A.; Kołakowski, P.; Pawlikowska, K.; Troszyńska, A. Impact of transglutaminase reaction on the immunoreactive and sensory quality of yoghurt starter. World J. Microbiol. Biotechnol. 2011, 27, 215–227. [Google Scholar] [CrossRef]
- Standen, B.; Rawling, M.; Davies, S.; Castex, M.; Foey, A.; Gioacchini, G.; Carnevali, O.; Merrifield, D. Probiotic Pediococcus acidilactici modulates both localised intestinal-and peripheral-immunity in tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2013, 35, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Villamil, L.; Reyes, C.; Martínez-Silva, M. In vivo and in vitro assessment of Lactobacillus acidophilus as probiotic for tilapia (Oreochromis niloticus, Perciformes: Cichlidae) culture improvement. Aquac. Res. 2014, 45, 1116–1125. [Google Scholar] [CrossRef]
- Daudelin, J.-F.; Lessard, M.; Beaudoin, F.; Nadeau, É.; Bissonnette, N.; Boutin, Y.; Brousseau, J.-P.; Lauzon, K.; Fairbrother, J.M. Administration of probiotics influences F4 (K88)-positive enterotoxigenic Escherichia coli attachment and intestinal cytokine expression in weaned pigs. Vet. Res. 2011, 42, 69. [Google Scholar] [CrossRef] [PubMed]
- Ragland, S.A.; Criss, A.K. From bacterial killing to immune modulation: Recent insights into the functions of lysozyme. PLoS Pathog. 2017, 13, e1006512. [Google Scholar] [CrossRef] [PubMed]
- Neveling, D.P.; van Emmenes, L.; Ahire, J.J.; Pieterse, E.; Smith, C.; Dicks, L. Effect of a Multi-Species Probiotic on the Colonisation of Salmonella in Broilers. Probiotics Antimicrob. Proteins 2019, 1–10. [Google Scholar] [CrossRef]
- Fasina, Y.; Holt, P.; Moran, E.; Moore, R.; Conner, D.; McKee, S. Intestinal cytokine response of commercial source broiler chicks to Salmonella typhimurium infection. Poult. Sci. 2008, 87, 1335–1346. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Tsen, H.-Y.; Lin, C.-L.; Yu, B.; Chen, C.-S. Oral administration of a combination of select lactic acid bacteria strains to reduce the Salmonella invasion and inflammation of broiler chicks. Poult. Sci. 2012, 91, 2139–2147. [Google Scholar] [CrossRef]
- Ray, A.; Ghosh, K.; Ringø, E. Enzyme-producing bacteria isolated from fish gut: A review. Aquac. Nutr. 2012, 18, 465–492. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Dhama, K.; Latheef, S.K.; Munjal, A.K.; Khandia, R.; Samad, H.A.; Iqbal, H.M.N.; Joshi, S.K. Probiotics in curing allergic and inflammatory conditions-research progress and futuristic vision. Recent Pat. Inflamm. Allergy Drug Discov. 2016, 10, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Huang, C.; He, T.; Harmsen, H. Effect of probiotics and yogurt on colonic microflora in subjects with lactose intolerance. J. Hyg. Res. 2006, 35, 587–591. [Google Scholar]
- Essa, M.A.; El-Serafy, S.; El-Ezabi, M.M.; Daboor, S.M.; Esmael, N.A.; Lall, S.P. Effect of different dietary probiotics on growth, feed utilization and digestive enzymes activities of Nile tilapia, Oreochromis niloticus. J. Arab. Aquac. Soc. 2010, 5, 143–162. [Google Scholar]
- Emami, N.K.; Samie, A.; Rahmani, H.; Ruiz-Feria, C. The effect of peppermint essential oil and fructooligosaccharides, as alternatives to virginiamycin, on growth performance, digestibility, gut morphology and immune response of male broilers. Anim. Feed Sci. Technol. 2012, 175, 57–64. [Google Scholar] [CrossRef]
- Standen, B.; Rodiles, A.; Peggs, D.; Davies, S.; Santos, G.; Merrifield, D. Modulation of the intestinal microbiota and morphology of tilapia, Oreochromis niloticus, following the application of a multi-species probiotic. Appl. Microbiol. Biotechnol. 2015, 99, 8403–8417. [Google Scholar] [CrossRef] [PubMed]
- Al-Fataftah, A.-R.; Abdelqader, A. Effects of dietary Bacillus subtilis on heat-stressed broilers performance, intestinal morphology and microflora composition. Anim. Feed Sci. Technol. 2014, 198, 279–285. [Google Scholar] [CrossRef]
- Burgain, J.; Scher, J.; Francius, G.; Borges, F.; Corgneau, M.; Revol-Junelles, A.; Cailliez-Grimal, C.; Gaiani, C. Lactic acid bacteria in dairy food: Surface characterization and interactions with food matrix components. Adv. Colloid Interface Sci. 2014, 213, 21–35. [Google Scholar] [CrossRef]
- Gueimonde, M.; Jalonen, L.; He, F.; Hiramatsu, M.; Salminen, S. Adhesion and competitive inhibition and displacement of human enteropathogens by selected lactobacilli. Food Res. Int. 2006, 39, 467–471. [Google Scholar] [CrossRef]
- Castagliuolo, I.; Galeazzi, F.; Ferrari, S.; Elli, M.; Brun, P.; Cavaggioni, A.; Tormen, D.; Sturniolo, G.C.; Morelli, L.; Palù, G. Beneficial effect of auto-aggregating Lactobacillus crispatus on experimentally induced colitis in mice. FEMS Immunol. Med. Microbiol. 2005, 43, 197–204. [Google Scholar] [CrossRef][Green Version]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef]
- Lee, Y.-K.; Puong, K.-Y. Competition for adhesion between probiotics and human gastrointestinal pathogens in the presence of carbohydrate. Br. J. Nutr. 2002, 88, S101–S108. [Google Scholar] [CrossRef] [PubMed]
- Ołdak, A.; Zielińska, D.; Łepecka, A.; Długosz, E.; Kołożyn-Krajewska, D. Lactobacillus plantarum strains isolated from Polish regional cheeses exhibit anti-staphylococcal activity and selected probiotic properties. Probiotics Antimicrob. Proteins 2020, 12, 1025–1038. [Google Scholar] [CrossRef]
- Guo, X.; Li, D.; Lu, W.; Piao, X.; Chen, X. Screening of Bacillus strains as potential probiotics and subsequent confirmation of the in vivo effectiveness of Bacillus subtilis MA139 in pigs. Antonie Van Leeuwenhoek 2006, 90, 139–146. [Google Scholar] [CrossRef]
- Alvarez-Sieiro, P.; Montalbán-López, M.; Mu, D.; Kuipers, O.P. Bacteriocins of lactic acid bacteria: Extending the family. Appl. Microbiol. Biotechnol. 2016, 100, 2939–2951. [Google Scholar] [CrossRef]
- Cavicchioli, V.; Camargo, A.; Todorov, S.; Nero, L. Novel bacteriocinogenic Enterococcus hirae and Pediococcus pentosaceus strains with antilisterial activity isolated from Brazilian artisanal cheese. J. Dairy Sci. 2017, 100, 2526–2535. [Google Scholar] [CrossRef]
- Heu, S.; Oh, J.; Kang, Y.; Ryu, S.; Cho, S.K.; Cho, Y.; Cho, M. gly gene cloning and expression and purification of glycinecin A, a bacteriocin produced by Xanthomonas campestris pv. glycines 8ra. Appl. Environ. Microbiol. 2001, 67, 4105–4110. [Google Scholar] [CrossRef] [PubMed]
- Deegan, L.H.; Cotter, P.D.; Hill, C.; Ross, P. Bacteriocins: Biological tools for bio-preservation and shelf-life extension. Int. Dairy J. 2006, 16, 1058–1071. [Google Scholar] [CrossRef]
- Olbrich dos Santos, K.M.; Silva Vieira, A.D.; Salles, H.O.; Oliveira, J.d.S.; Costa Rocha, C.R.; Borges, M.d.F.; Bruno, L.M.; de Melo Franco, B.D.G.; Todorov, S.D. Safety, beneficial and technological properties of Enterococcus faecium isolated from Brazilian cheeses. Braz. J. Microbiol. 2015, 46, 237. [Google Scholar] [CrossRef]
- Harnentis, H.; Nurmiati, N.; Marlida, Y.; Adzitey, F.; Huda, N. γ-Aminobutyric acid production by selected lactic acid bacteria isolate of an Indonesian indigenous fermented buffalo milk (dadih) origin. Vet. World 2019, 12, 1352. [Google Scholar] [CrossRef]
- Dhakal, R.; Bajpai, V.K.; Baek, K.-H. Production of GABA (γ-aminobutyric acid) by microorganisms: A review. Braz. J. Microbiol. 2012, 43, 1230–1241. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Chousalkar, K.K. Short-term feeding of probiotics and synbiotics modulates caecal microbiota during Salmonella Typhimurium infection but does not reduce shedding and invasion in chickens. Appl. Microbiol. Biotechnol. 2020, 104, 319–334. [Google Scholar] [CrossRef] [PubMed]
- McFarland, L.V. Efficacy of Single-Strain Probiotics Versus Multi-Strain Mixtures: Systematic Review of Strain and Disease Specificity. Dig. Dis. Sci. 2020. [Google Scholar] [CrossRef]
- Douillard, F.P.; Mora, D.; Eijlander, R.T.; Wels, M.; De Vos, W.M. Comparative genomic analysis of the multispecies probiotic-marketed product VSL# 3. PLoS ONE 2018, 13, e0192452. [Google Scholar]
- Kankainen, M.; Paulin, L.; Tynkkynen, S.; von Ossowski, I.; Reunanen, J.; Partanen, P.; Satokari, R.; Vesterlund, S.; Hendrickx, A.P.; Lebeer, S. Comparative genomic analysis of Lactobacillus rhamnosus GG reveals pili containing a human-mucus binding protein. Proc. Natl. Acad. Sci. USA 2009, 106, 17193–17198. [Google Scholar] [CrossRef]
- Motherway, M.O.C.; Zomer, A.; Leahy, S.C.; Reunanen, J.; Bottacini, F.; Claesson, M.J.; O’Brien, F.; Flynn, K.; Casey, P.G.; Munoz, J.A.M. Functional genome analysis of Bifidobacterium breve UCC2003 reveals type IVb tight adherence (Tad) pili as an essential and conserved host-colonization factor. Proc. Natl. Acad. Sci. USA 2011, 108, 11217–11222. [Google Scholar] [CrossRef]
- Sengupta, R.; Altermann, E.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Roy, N.C. The role of cell surface architecture of lactobacilli in host-microbe interactions in the gastrointestinal tract. Mediat. Inflamm. 2013, 2013, 237921. [Google Scholar] [CrossRef]
- Fredriksen, L.; Moen, A.; Adzhubei, A.A.; Mathiesen, G.; Eijsink, V.G.; Egge-Jacobsen, W. Lactobacillus plantarum WCFS1 O-linked protein glycosylation: An extended spectrum of target proteins and modification sites detected by mass spectrometry. Glycobiology 2013, 23, 1439–1451. [Google Scholar] [CrossRef]
- Tytgat, H.L.; De Vos, W.M. Sugar coating the envelope: Glycoconjugates for microbe–host crosstalk. Trends Microbiol. 2016, 24, 853–861. [Google Scholar] [CrossRef]
- Cazzola, M.; Tompkins, T.A.; Matera, M.G. Immunomodulatory impact of a synbiotic in Th1 and Th2 models of infection. Ther. Adv. Respir. Dis. 2010, 4, 259–270. [Google Scholar] [CrossRef]
- Tien, M.-T.; Girardin, S.E.; Regnault, B.; Le Bourhis, L.; Dillies, M.-A.; Coppée, J.-Y.; Bourdet-Sicard, R.; Sansonetti, P.J.; Pédron, T. Anti-inflammatory effect of Lactobacillus casei on Shigella-infected human intestinal epithelial cells. J. Immunol. 2006, 176, 1228–1237. [Google Scholar] [CrossRef]
- Di Cagno, R.; De Angelis, M.; Limitone, A.; Minervini, F.; Simonetti, M.C.; Buchin, S.; Gobbetti, M. Cell–cell communication in sourdough lactic acid bacteria: A proteomic study in Lactobacillus sanfranciscensis CB1. Proteomics 2007, 7, 2430–2446. [Google Scholar] [CrossRef]
- Xavier, K.B.; Bassler, B.L. LuxS quorum sensing: More than just a numbers game. Curr. Opin. Microbiol. 2003, 6, 191–197. [Google Scholar] [CrossRef]
- Nakayama, J.; Cao, Y.; Horii, T.; Sakuda, S.; Akkermans, A.D.; De Vos, W.M.; Nagasawa, H. Gelatinase biosynthesis-activating pheromone: A peptide lactone that mediates a quorum sensing in Enterococcus faecalis. Mol. Microbiol. 2001, 41, 145–154. [Google Scholar] [CrossRef]
- Sturme, M.H.; Francke, C.; Siezen, R.J.; de Vos, W.M.; Kleerebezem, M. Making sense of quorum sensing in lactobacilli: A special focus on Lactobacillus plantarum WCFS1. Microbiology 2007, 153, 3939–3947. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; De Angelis, M.; Coda, R.; Minervini, F.; Gobbetti, M. Molecular adaptation of sourdough Lactobacillus plantarum DC400 under co-cultivation with other lactobacilli. Res. Microbiol. 2009, 160, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Diep, D.; Mathiesen, G.; Eijsink, V.; Nes, I. Use of lactobacilli and their pheromone-based regulatory mechanism in gene expression and drug delivery. Curr. Pharm. Biotechnol. 2009, 10, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, A.; Ruiz-Barba, J.L.; Jiménez-Díaz, R. Production of plantaricin NC8 by Lactobacillus plantarum NC8 is induced in the presence of different types of gram-positive bacteria. Arch. Microbiol. 2004, 181, 8–16. [Google Scholar] [CrossRef]
- Di Cagno, R.; De Angelis, M.; Calasso, M.; Vincentini, O.; Vernocchi, P.; Ndagijimana, M.; De Vincenzi, M.; Dessì, M.R.; Guerzoni, M.E.; Gobbetti, M. Quorum sensing in sourdough Lactobacillus plantarum DC400: Induction of plantaricin A (PlnA) under co-cultivation with other lactic acid bacteria and effect of PlnA on bacterial and Caco-2 cells. Proteomics 2010, 10, 2175–2190. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Hu, J. Positive effect of probiotics on constipation in children: A systematic review and meta-analysis of six randomized controlled trials. Front. Cell. Infect. Microbiol. 2017, 7, 153. [Google Scholar] [CrossRef]
- Algburi, A.; Alazzawi, S.A.; Al-Ezzy, A.I.A.; Weeks, R.; Chistyakov, V.; Chikindas, M.L. Potential Probiotics Bacillus subtilis KATMIRA1933 and Bacillus amyloliquefaciens B-1895 Co-Aggregate with Clinical Isolates of Proteus mirabilis and Prevent Biofilm Formation. Probiotics Antimicrob. Proteins 2020, 12, 1471–1483. [Google Scholar] [CrossRef] [PubMed]
- Parker, E.A.; Roy, T.; D’Adamo, C.R.; Wieland, L.S. Probiotics and gastrointestinal conditions: An overview of evidence from the Cochrane Collaboration. Nutrition 2018, 45, 125–134.e111. [Google Scholar] [CrossRef]
- Zhang, Z.; Kim, I. Effects of multistrain probiotics on growth performance, apparent ileal nutrient digestibility, blood characteristics, cecal microbial shedding, and excreta odor contents in broilers. Poult. Sci. 2014, 93, 364–370. [Google Scholar] [CrossRef]
- Bernstein, C.N. Antibiotics, probiotics and prebiotics in IBD. In Nutrition, Gut Microbiota and Immunity: Therapeutic Targets for IBD; Karger Publishers: Basel, Switzerland, 2014; Volume 79, pp. 83–100. [Google Scholar]
- Hajela, N.; Ramakrishna, B.; Nair, G.B.; Abraham, P.; Gopalan, S.; Ganguly, N.K. Gut microbiome, gut function, and probiotics: Implications for health. Indian J. Gastroenterol. 2015, 34, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Frech, T.M.; Khanna, D.; Maranian, P.; Frech, E.J.; Sawitzke, A.D.; Murtaugh, M.A. Probiotics for the treatment of systemic sclerosis-associated gastrointestinal bloating/distention. Clin. Exp. Rheumatol. 2011, 29, S22. [Google Scholar]
- Yoon, J.S.; Sohn, W.; Lee, O.Y.; Lee, S.P.; Lee, K.N.; Jun, D.W.; Lee, H.L.; Yoon, B.C.; Choi, H.S.; Chung, W.S. Effect of multispecies probiotics on irritable bowel syndrome: A randomized, double-blind, placebo-controlled trial. J. Gastroenterol. Hepatol. 2014, 29, 52–59. [Google Scholar] [CrossRef]
- Leventogiannis, K.; Gkolfakis, P.; Spithakis, G.; Tsatali, A.; Pistiki, A.; Sioulas, A.; Giamarellos-Bourboulis, E.J.; Triantafyllou, K. Effect of a preparation of four probiotics on symptoms of patients with irritable bowel syndrome: Association with intestinal bacterial overgrowth. Probiotics Antimicrob. Proteins 2019, 11, 627–634. [Google Scholar] [CrossRef]
- Gareau, M.G. Microbiota-gut-brain axis and cognitive function. In Microbial Endocrinology: The Microbiota-Gut-Brain Axis in Health and Disease; Springer: Berlin/Heidelberg, Germany, 2014; pp. 357–371. [Google Scholar]
- Akbari, E.; Asemi, Z.; Daneshvar Kakhaki, R.; Bahmani, F.; Kouchaki, E.; Tamtaji, O.R.; Hamidi, G.A.; Salami, M. Effect of probiotic supplementation on cognitive function and metabolic status in Alzheimer’s disease: A randomized, double-blind and controlled trial. Front. Aging Neurosci. 2016, 8, 256. [Google Scholar] [CrossRef]
- Sabico, S.; Al-Mashharawi, A.; Al-Daghri, N.M.; Wani, K.; Amer, O.E.; Hussain, D.S.; Ansari, M.G.A.; Masoud, M.S.; Alokail, M.S.; McTernan, P.G. Effects of a 6-month multi-strain probiotics supplementation in endotoxemic, inflammatory and cardiometabolic status of T2DM patients: A randomized, double-blind, placebo-controlled trial. Clin. Nutr. 2019, 38, 1561–1569. [Google Scholar] [CrossRef]
- Allen, S.J.; Jordan, S.; Storey, M.; Thornton, C.A.; Gravenor, M.B.; Garaiova, I.; Plummer, S.F.; Wang, D.; Morgan, G. Probiotics in the prevention of eczema: A randomised controlled trial. Arch. Dis. Child. 2014, 99, 1014–1019. [Google Scholar] [CrossRef]
- Jadhav, S.R.; Shandilya, U.K.; Kansal, V.K. Immunoprotective effect of probiotic dahi containing Lactobacillus acidophilus and Bifidobacterium bifidum on dextran sodium sulfate-induced ulcerative colitis in mice. Probiotics Antimicrob. Proteins 2012, 4, 21–26. [Google Scholar] [CrossRef]
- Chen, Y.; Schwack, W. High-performance thin-layer chromatography screening of multi class antibiotics in animal food by bioluminescent bioautography and electrospray ionization mass spectrometry. J. Chromatogr. A 2014, 1356, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Ambalam, P.; Kondepudi, K.K.; Balusupati, P.; Nilsson, I.; Wadström, T.; Ljungh, Å. Prebiotic preferences of human lactobacilli strains in co-culture with bifidobacteria and antimicrobial activity against Clostridium difficile. J. Appl. Microbiol. 2015, 119, 1672–1682. [Google Scholar] [CrossRef] [PubMed]
- Kondepudi, K.K.; Ambalam, P.; Karagin, P.H.; Nilsson, I.; Wadström, T.; Ljungh, Å. A novel multi-strain probiotic and synbiotic supplement for prevention of Clostridium difficile infection in a murine model. Microbiol. Immunol. 2014, 58, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Khanna, S.; Pardi, D.S. Clostridium difficile infection: Management strategies for a difficult disease. Ther. Adv. Gastroenterol. 2014, 7, 72–86. [Google Scholar] [CrossRef] [PubMed]
- Ouwehand, A.C.; Forssten, S.; Hibberd, A.A.; Lyra, A.; Stahl, B. Probiotic approach to prevent antibiotic resistance. Ann. Med. 2016, 48, 246–255. [Google Scholar] [CrossRef]
- Lakhtin, M.; Alyoshkin, V.; Lakhtin, V.; Afanasyev, S.; Pozhalostina, L.; Pospelova, V. Probiotic lactobacillus and bifidobacterial lectins against Candida albicans and Staphylococcus aureus clinical strains: New class of the pathogen biofilm destructors. Probiotics Antimicrob. Proteins 2010, 2, 186–196. [Google Scholar] [CrossRef]
- Mansour, N.M.; Elkhatib, W.F.; Aboshanab, K.M.; Bahr, M.M. Inhibition of Clostridium difficile in mice using a mixture of potential probiotic strains Enterococcus faecalis NM815, E. faecalis NM915, and E. faecium NM1015: Novel candidates to control C. difficile infection (CDI). Probiotics Antimicrob. Proteins 2018, 10, 511–522. [Google Scholar] [CrossRef]
- Reid, G.; Bruce, A.W. Selection of Lactobacillus strains for urogenital probiotic applications. J. Infect. Dis. 2001, 183, S77–S80. [Google Scholar] [CrossRef]
- Zhong, J.; Zhang, F.; Peng, Y.; Ji, Z.; Li, H.; Li, S.; Zhang, X.; Shi, Q.; Zhang, J. Mixed culture of probiotics on a solid-state medium: An efficient method to produce an affordable probiotic feed additive. Biotechnol. Bioprocess Eng. 2017, 22, 758–766. [Google Scholar] [CrossRef]
- Karimi, G.; Jamaluddin, R.; Mohtarrudin, N.; Ahmad, Z.; Khazaai, H.; Parvaneh, M. Single-species versus dual-species probiotic supplementation as an emerging therapeutic strategy for obesity. Nutr. Metab. Cardiovasc. Dis. 2017, 27, 910–918. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Zhang, S.; Liu, X.; Huo, Y.; Su, B.; Li, X. Effects of a probiotic intervention on Escherichia coli and high-fat diet-induced intestinal microbiota imbalance. Appl. Microbiol. Biotechnol. 2020, 104, 1243–1257. [Google Scholar] [CrossRef] [PubMed]
- Wolff, B.J.; Price, T.K.; Joyce, C.J.; Wolfe, A.J.; Mueller, E.R. Oral probiotics and the female urinary microbiome: A double-blinded randomized placebo-controlled trial. Int. Urol. Nephrol. 2019, 1–11. [Google Scholar] [CrossRef]
- Bidossi, A.; De Grandi, R.; Toscano, M.; Bottagisio, M.; De Vecchi, E.; Gelardi, M.; Drago, L. Probiotics Streptococcus salivarius 24SMB and Streptococcus oralis 89a interfere with biofilm formation of pathogens of the upper respiratory tract. BMC Infect. Dis. 2018, 18, 653. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Leonhard, M.; Moser, D.; Ma, S.; Schneider-Stickler, B. Inhibitory effect of probiotic lactobacilli supernatants on single and mixed non-albicans Candida species biofilm. Arch. Oral Biol. 2018, 85, 40–45. [Google Scholar] [CrossRef]
- Ahmed, S.; Mun, H.-S.; Islam, M.; Kim, S.-S.; Hwang, J.-A.; Kim, Y.-J.; Yang, C.-J. Effects of Citrus junos by-products fermented with multistrain probiotics on growth performance, immunity, caecal microbiology and meat oxidative stability in broilers. Br. Poult. Sci. 2014, 55, 540–547. [Google Scholar] [CrossRef]
- Sabetkish, N.; Sabetkish, S.; Mohseni, M.J.; Kajbafzadeh, A.-M. Prevention of renal scarring in acute pyelonephritis by probiotic therapy: An experimental study. Probiotics Antimicrob. Proteins 2019, 11, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Redweik, G.A.; Stromberg, Z.R.; Van Goor, A.; Mellata, M. Protection against avian pathogenic Escherichia coli and Salmonella Kentucky exhibited in chickens given both probiotics and live Salmonella vaccine. Poult. Sci. 2020, 99, 752–762. [Google Scholar] [CrossRef]
- Piyadeatsoontorn, S.; Taharnklaew, R.; Upathanpreecha, T.; Sornplang, P. Encapsulating viability of multi-strain Lactobacilli as potential probiotic in pigs. Probiotics Antimicrob. Proteins 2019, 11, 438–446. [Google Scholar] [CrossRef]
- Sarjapuram, N.; Mekala, N.; Singh, M.; Tatu, U. The potential of Lactobacillus casei and Entercoccus faecium combination as a preventive probiotic against Entamoeba. Probiotics Antimicrob. Proteins 2017, 9, 142–149. [Google Scholar] [CrossRef]
- Zambori, C.; Morvay, A.A.; Sala, C.; Licker, M.; Gurban, C.; Tanasie, G.; Tîrziu, E. Antimicrobial effect of probiotics on bacterial species from dental plaque. J. Infect. Dev. Ctries. 2016, 10, 214–221. [Google Scholar] [CrossRef][Green Version]
- Ahmed, S.T.; Hoon, J.; Mun, H.-S.; Yang, C.-J. Evaluation of Lactobacillus and Bacillus-based probiotics as alternatives to antibiotics in enteric microbial challenged weaned piglets. Afr. J. Microbiol. Res. 2014, 8, 96–104. [Google Scholar]
- Tejero-Sariñena, S.; Barlow, J.; Costabile, A.; Gibson, G.R.; Rowland, I. Antipathogenic activity of probiotics against Salmonella Typhimurium and Clostridium difficile in anaerobic batch culture systems: Is it due to synergies in probiotic mixtures or the specificity of single strains? Anaerobe 2013, 24, 60–65. [Google Scholar] [CrossRef]
- Lollo, P.C.B.; de Moura, C.S.; Morato, P.N.; Cruz, A.G.; de Freitas Castro, W.; Betim, C.B.; Nisishima, L.; José de Assis, F.F.; Junior, M.M.; Fernandes, C.O. Probiotic yogurt offers higher immune-protection than probiotic whey beverage. Food Res. Int. 2013, 54, 118–124. [Google Scholar] [CrossRef]
- de Souza, B.M.S.; Borgonovi, T.F.; Casarotti, S.N.; Todorov, S.D.; Penna, A.L.B. Lactobacillus casei and Lactobacillus fermentum strains isolated from mozzarella cheese: Probiotic potential, safety, acidifying kinetic parameters and viability under gastrointestinal tract conditions. Probiotics Antimicrob. Proteins 2019, 11, 382–396. [Google Scholar] [CrossRef] [PubMed]
- Pawłowska, J.; Klewicka, E.; Czubkowski, P.; Motyl, I.; Jankowska, I.; Libudzisz, Z.; Teisseyre, M.; Gliwicz, D.; Cukrowska, B. Effect of Lactobacillus casei DN-114001 application on the activity of fecal enzymes in children after liver transplantation. In Transplantation Proceedings; Elsevier: Amsterdam, The Netherlands, 2007; pp. 3219–3221. [Google Scholar]
- Schwenger, E.M.; Tejani, A.M.; Loewen, P.S. Probiotics for preventing urinary tract infections in adults and children. Cochrane Database Syst. Rev. 2015. [Google Scholar] [CrossRef] [PubMed]
- Sharif, A.; Kashani, H.H.; Nasri, E.; Soleimani, Z.; Sharif, M.R. The role of probiotics in the treatment of dysentery: A randomized double-blind clinical trial. Probiotics Antimicrob. Proteins 2017, 9, 380–385. [Google Scholar] [CrossRef]
- Guarner, F.; Malagelada, J.-R. Gut flora in health and disease. Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef]
- Lan, R.; Lee, S.; Kim, I. Effects of multistrain probiotics on growth performance, nutrient digestibility, blood profiles, faecal microbial shedding, faecal score and noxious gas emission in weaning pigs. J. Anim. Physiol. Anim. Nutr. 2016, 100, 1130–1138. [Google Scholar] [CrossRef]
- Asai, T.; Masani, K.; Sato, C.; Hiki, M.; Usui, M.; Baba, K.; Ozawa, M.; Harada, K.; Aoki, H.; Sawada, T. Phylogenetic groups and cephalosporin resistance genes of Escherichia coli from diseased food-producing animals in Japan. Acta Vet. Scand. 2011, 53, 52. [Google Scholar] [CrossRef]
- Deniz, G.; Orman, A.; Cetinkaya, F.; Gencoglu, H.; Meral, Y.; Turkmen, I. Effects of probiotic (Bacillus subtilis DSM 17299) supplementation on the caecal microflora and performance in broiler chickens. Rev. Méd. Vét. 2011, 162, 538–545. [Google Scholar]
- Mountzouris, K.; Tsirtsikos, P.; Kalamara, E.; Nitsch, S.; Schatzmayr, G.; Fegeros, K. Evaluation of the efficacy of a probiotic containing Lactobacillus, Bifidobacterium, Enterococcus, and Pediococcus strains in promoting broiler performance and modulating cecal microflora composition and metabolic activities. Poult. Sci. 2007, 86, 309–317. [Google Scholar] [CrossRef]
- Yang, C.; Cao, G.; Ferket, P.; Liu, T.; Zhou, L.; Zhang, L.; Xiao, Y.; Chen, A. Effects of probiotic, Clostridium butyricum, on growth performance, immune function, and cecal microflora in broiler chickens. Poult. Sci. 2012, 91, 2121–2129. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, P.; Sahoo, J.; Panda, P. Effect of probiotic (Lacto-Sacc) feeding, packaging methods and seasons on the microbial and organoleptic qualities of chicken meat balls during refrigerated storage. J. Food Sci. Technol. (Mysore) 2000, 37, 67–71. [Google Scholar]
- Lee, K.; Lillehoj, H.S.; Siragusa, G.R. Direct-fed microbials and their impact on the intestinal microflora and immune system of chickens. J. Poult. Sci. 2010. [Google Scholar] [CrossRef]
- Kazemi, S.A.; Ahmadi, H.; Karimi Torshizi, M.A. Evaluating two multistrain probiotics on growth performance, intestinal morphology, lipid oxidation and ileal microflora in chickens. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1399–1407. [Google Scholar] [CrossRef]
- Gadde, U.; Oh, S.; Lee, Y.; Davis, E.; Zimmerman, N.; Rehberger, T.; Lillehoj, H.S. The effects of direct-fed microbial supplementation, as an alternative to antibiotics, on growth performance, intestinal immune status, and epithelial barrier gene expression in broiler chickens. Probiotics Antimicrob. Proteins 2017, 9, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Balevi, T.; Ucan, U.; Coşun, B.; Kurtoğu, V.; Cetingül, I. Effect of dietary probiotic on performance and humoral immune response in layer hens. Br. Poult. Sci. 2001, 42, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Gil De Los Santos, J.; Storch, O.; Gil-Turnes, C. Bacillus cereus var. toyoii and Saccharomyces boulardii increased feed efficiency in broilers infected with Salmonella enteritidis. Br. Poult. Sci. 2005, 46, 494–497. [Google Scholar] [CrossRef]
- Mountzouris, K.; Tsitrsikos, P.; Palamidi, I.; Arvaniti, A.; Mohnl, M.; Schatzmayr, G.; Fegeros, K. Effects of probiotic inclusion levels in broiler nutrition on growth performance, nutrient digestibility, plasma immunoglobulins, and cecal microflora composition. Poult. Sci. 2010, 89, 58–67. [Google Scholar] [CrossRef]
- Bairagi, A.; Sarkar Ghosh, K.; Sen, S.; Ray, A. Evaluation of the nutritive value of Leucaena leucocephala leaf meal, inoculated with fish intestinal bacteria Bacillus subtilis and Bacillus circulans in formulated diets for rohu, Labeo rohita (Hamilton) fingerlings. Aquac. Res. 2004, 35, 436–446. [Google Scholar] [CrossRef]
- Hong, H.A.; Duc, L.H.; Cutting, S.M. The use of bacterial spore formers as probiotics. FEMS Microbiol. Rev. 2005, 29, 813–835. [Google Scholar] [CrossRef] [PubMed]
- Standen, B.; Peggs, D.; Rawling, M.; Foey, A.; Davies, S.; Santos, G.; Merrifield, D. Dietary administration of a commercial mixed-species probiotic improves growth performance and modulates the intestinal immunity of tilapia, Oreochromis niloticus. Fish Shellfish Immunol. 2016, 49, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Yeh, S.P.; Chiu, C.H.; Shiu, Y.L.; Huang, Z.L.; Liu, C.H. Effects of diets supplemented with either individual or combined probiotics, B acillus subtilis E20 and L actobacillus plantarum 7-40, on the immune response and disease resistance of the mud crab, S cylla paramamosain (E stampador). Aquac. Res. 2014, 45, 1164–1175. [Google Scholar] [CrossRef]
- Dlamini, Z.; Langa, R.; Aiyegoro, O.; Okoh, A. Effects of probiotics on growth performance, blood parameters, and antibody stimulation in piglets. S. Afr. J. Anim. Sci. 2017, 47, 765–776. [Google Scholar] [CrossRef]
- Castillo, M.; Martín-Orúe, S.M.; Nofrarías, M.; Manzanilla, E.G.; Gasa, J. Changes in caecal microbiota and mucosal morphology of weaned pigs. Vet. Microbiol. 2007, 124, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Rhouma, M.; Fairbrother, J.M.; Beaudry, F.; Letellier, A. Post weaning diarrhea in pigs: Risk factors and non-colistin-based control strategies. Acta Vet. Scand. 2017, 59, 31. [Google Scholar] [CrossRef]
- Luppi, A.; Gibellini, M.; Gin, T.; Vangroenweghe, F.; Vandenbroucke, V.; Bauerfeind, R.; Bonilauri, P.; Labarque, G.; Hidalgo, Á. Prevalence of virulence factors in enterotoxigenic Escherichia coli isolated from pigs with post-weaning diarrhoea in Europe. Porc. Health Manag. 2016, 2, 20. [Google Scholar] [CrossRef]
- Schlee, M.; Harder, J.; Köten, B.; Stange, E.; Wehkamp, J.; Fellermann, K. Probiotic lactobacilli and VSL# 3 induce enterocyte β-defensin 2. Clin. Exp. Immunol. 2008, 151, 528–535. [Google Scholar]
- Lee, S.; Lillehoj, H.; Dalloul, R.; Park, D.; Hong, Y.; Lin, J. Influence of Pediococcus-based probiotic on coccidiosis in broiler chickens. Poult. Sci. 2007, 86, 63–66. [Google Scholar] [CrossRef]
- Brousseau, J.-P.; Talbot, G.; Beaudoin, F.; Lauzon, K.; Roy, D.; Lessard, M. Effects of probiotics Pediococcus acidilactici strain MA18/5M and Saccharomyces cerevisiae subsp. boulardii strain SB-CNCM I-1079 on fecal and intestinal microbiota of nursing and weanling piglets. J. Anim. Sci. 2015, 93, 5313–5326. [Google Scholar] [CrossRef]
- Laskowska, E.; Jarosz, Ł.; Grądzki, Z. Effect of multi-microbial probiotic formulation Bokashi on pro-and anti-inflammatory cytokines profile in the serum, colostrum and milk of sows, and in a culture of polymorphonuclear cells isolated from colostrum. Probiotics Antimicrob. Proteins 2019, 11, 220–232. [Google Scholar] [CrossRef]
- El-Tawab, M.A.; Youssef, I.; Bakr, H.; Fthenakis, G.; Giadinis, N. Role of probiotics in nutrition and health of small ruminants. Pol. J. Vet. Sci. 2016, 19, 893–906. [Google Scholar] [CrossRef][Green Version]
- Adetoye, A.; Pinloche, E.; Adeniyi, B.A.; Ayeni, F.A. Characterization and anti-salmonella activities of lactic acid bacteria isolated from cattle faeces. BMC Microbiol. 2018, 18, 96. [Google Scholar] [CrossRef]
- Renaud, D.; Kelton, D.; Weese, J.; Noble, C.; Duffield, T. Evaluation of a multispecies probiotic as a supportive treatment for diarrhea in dairy calves: A randomized clinical trial. J. Dairy Sci. 2019, 102, 4498–4505. [Google Scholar] [CrossRef] [PubMed]
- Signorini, M.; Soto, L.; Zbrun, M.; Sequeira, G.; Rosmini, M.; Frizzo, L. Impact of probiotic administration on the health and fecal microbiota of young calves: A meta-analysis of randomized controlled trials of lactic acid bacteria. Res. Vet. Sci. 2012, 93, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Timmerman, H.; Mulder, L.; Everts, H.; Van Espen, D.; Van Der Wal, E.; Klaassen, G.; Rouwers, S.; Hartemink, R.; Rombouts, F.; Beynen, A. Health and growth of veal calves fed milk replacers with or without probiotics. J. Dairy Sci. 2005, 88, 2154–2165. [Google Scholar] [CrossRef]
- Deng, Q.; Odhiambo, J.; Farooq, U.; Lam, T.; Dunn, S.; Ametaj, B. Intravaginal probiotics modulated metabolic status and improved milk production and composition of transition dairy cows. J. Anim. Sci. 2016, 94, 760–770. [Google Scholar] [CrossRef]
- Olchowy, T.; Soust, M.; Alawneh, J. The effect of a commercial probiotic product on the milk quality of dairy cows. J. Dairy Sci. 2019, 102, 2188–2195. [Google Scholar] [CrossRef]
- Karamali, M.; Nasiri, N.; Shavazi, N.T.; Jamilian, M.; Bahmani, F.; Tajabadi-Ebrahimi, M.; Asemi, Z. The effects of synbiotic supplementation on pregnancy outcomes in gestational diabetes. Probiotics Antimicrob. Proteins 2018, 10, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Güney-Varal, İ.; Köksal, N.; Özkan, H.; Bağcı, O.; Doğan, P. The effect of early administration of combined multi-strain and multi-species probiotics on gastrointestinal morbidities and mortality in preterm infants: A randomized controlled trial in a tertiary care unit. Turk. J. Pediatr. 2017, 59, 13–19. [Google Scholar] [CrossRef]
- Kovachev, S.M.; Vatcheva-Dobrevska, R.S. Local probiotic therapy for vaginal Candida albicans infections. Probiotics Antimicrob. Proteins 2015, 7, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.; Zaneb, H.; Masood, S.; Khan, R.U.; Ashraf, S.; Sikandar, A.; Rehman, H.F.U.; Rehman, H.U. Effect of dietary supplementation of zinc and multi-microbe probiotic on growth traits and alteration of intestinal architecture in broiler. Probiotics Antimicrob. Proteins 2019, 11, 931–937. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Probiotics Mixture | Conditions | Mechanism of Actions | References |
|---|---|---|---|
| B. bifidum W23, B. lactis W52, L. acidophilus W37, L. brevis W63, L. casei W56, L. salivarius W24, Lactococcus lactis W19 and L. lactis W58 | Endotoxins | Improvement of endothelial barrier, inhibition of mast cell, activation of proinflammatory cytokines, and decrease endotoxin | [94] |
| L. acidophilus, L. casei, B. bifidum, and L. fermentum | Cognitive function in Alzheimer’s disease | [93] | |
| L. paracasei DSM 24,733, L. plantarum DSM 24,730, L. acidophilus DSM 24,735, and L. delbrueckii subspecies bulgaricus DSM 24,734), Bifidobacteria (B. longum DSM 24,736, B. breve DSM 24,732, and B. infantis DSM 24,737), and Streptococcus (S. thermophilus DSM 24,731) | Systemic sclerosis-associated gastrointestinal disease | Improvement of GI reflux and intestinal microbiota alpha diversity | [89] |
| L. acidophilus LaVK2 and B. bifidum Bbvk3 | Dextran sodium- sulphate salt-induced ulcerative colitis in mice | Reduction in myeloperoxidase activity, levels of TNF-α, IL-6, and IFN-γ | [96] |
| L. bulgaricus 151 and S. thermophilus MK-10 | Dextran sodium- sulphate salt-induced colitis | Modulation of intestinal microbiota, decrease the content of putrefactive short-chain fatty acid, enhanced production of cytokines | [28] |
| B. bifidum (KCTC 12199BP), B. lactis (KCTC 11904BP), B. longum (KCTC 12200BP), L. acidophilus (KCTC 11906BP), L. rhamnosus (KCTC 12202BP) and S. thermophilus (KCTC 11870BP) | Irritable Bowel Syndrome (IBS) | Alleviation of IBS symptoms and improvement of intestinal microbiota | [90] |
| B. longum and L. casei strain Shirota | Treatment of obesity | Decreased weight and triglyceride in rats fed with the high-fat diet. | [106] |
| S. boulardii, L. acidophilus, L. plantarum, B. lactis | IBS associated with bacterial overgrowth and constipation | Improvement in bloating, and pain associated with constipation | [91] |
| L. plantarum, B. breve, and L. fermentum | high-dietary fat-induced obesity and E. coli challenged | Causes reduced Lipopolysaccharide and IL-1β, improved the structure of intestinal flora and increased the fecal short-chain fatty acid (SCFA) content | [107] |
| Multi-Strain Probiotics Isolates | Pathogenic Bacteria | Host | References |
|---|---|---|---|
| B. subtilis and L. mesentroides | Vibrio cholereae | In-vitro agar diffusion test | [2] |
| L. plantarum F44, L. paracasei F8, B. breve 46 and B. lactis | Clostridium difficile | Mice | [99] |
| S. oralis and S. salivarius | Biofilm (S. aureus, S. epidermidis, S. pneumoniae, S. pyogenes, Propionibacterium acnes and Moraxella catarrhalis | Dogs | [109] |
| L. acidophilus LAP5, L. fermentum P2, P. acidophilus LS, and L. casei L21 | S. enterica subspecies Enterica | Chickens | [9] |
| L. acidophilus LA-5 and B. bifidum BB-12 | P. stomatis, P. multocida, P. canis, N. animaloris, and N. zoodegmatis | [116] | |
| P. acidilactici and S. cerevisiae boulardii | Enterotoxigenic E. coli (ETEC) F4 | Pigs | [38] |
| L. acidophilus NCIMB 30184, L. fermentum NCIMB 30226, L. plantarum NCIMB 30187, and L. rhamnosus NCIMB 30188 | Pathogenic E. coli and E. faecalis | [33] | |
| S. cerevisiae, E. faecium, L. acidophilus and Bacillus subtilis | E. coli | Chickens (broilers) | [117] |
| L. acidophilus NCIMB 30184, L. rhamnosus NCIMB 30188, L. plantarum NCIMB 30187, L. delbrueckii ssp. bulgaricus NCIMB 30186, L. casei NCIMB 30185, L. lactis NCIMB 30222, L. salivarius NCIMB 30225, L. fermentum NCIMB 30226, L. helveticus NCIMB 30224, B. bifidum NCIMB 30179, B. breve NCIMB 30180, B. infantis NCIMB 30181, B. longum NCIMB 30182, S. thermophilus NCIMB 30189 B. subtilis NCIMB 30223 | S. typhimurium, C. difficile | In-vitro distal colon model | [118] |
| L. acidophilus NCIMB 30184, L. fermentum NCIMB 30188, L. plantarum NCIMB 30187 and L. rhamnosus NCIMB 30226 | E. faecalis NCTC 0075 and E. coli NCTC 9001 | In-vitro agar diffusion test | [17] |
| L. rhamnosus and L. reuteri | Vaginal coliforms and yeast | Human (female) | [104] |
| L. crispatus, L. salivarius, L. gallinarum, L. johnsonii, E. faecalis and B. amyloliquefaciens | Salmonella Enteritidis A9 | Chickens (broiler) | [40] |
| L. acidophilus, L. fermentum, L. plantarum and E. faecium | Salmonella enterica | Chickens (broiler) | [42] |
| B. amyloliquefaciens B-1895 and B. subtilis KATMIRA1933 | Inhibits Proteus mirabilis biofilm formation | Invitro | [84] |
| E. faecalis (strains NM815, and NM915) and E. faecium NM1015 | C. difficile infection | Mice | [103] |
| L. acidophilus (LA-5), and B. animalis subspecies Lactis (Bb12) | E. coli induced pyelonephritis | Sprague-Dawley rat | [112] |
| L. casei and E. faecium | Entamoeba invadens | Invitro | [115] |
| B. subtilis, L. acidophilus, P. acidilactici, P. pentosus, Saccharomyces pastorianus | Avian pathogenic E. coli and Salmonella Kentucky | White leg-horn chicks | [113] |
| L. gasseri and L. rhamnosus | Non-Candida albicans biofilm formation | In-vitro | [110] |
| Synbiotics | Actions | Host | References |
|---|---|---|---|
| L. acidophilus strain T16, L. casei strain T2) and B. bifidum strain T1 plus 800mg inulin (HPX) | decreased the incidence of cesarean section rate and newborn’s hyperbilirubinemia and hospitalization | Human (pregnant women) | [156] |
| L. acidophilus, L. rhamnosus, S. thermophilus, and L. delbrueckii subspecies Bulgaricus plus fluconazole | Enhance the treatment of Vaginal candidiasis caused Candida albicans | humans | [158] |
| L. plantarum, L. acidophilus, L. delbrueckii subspecies bulgaricus, B. bifidum, L. rhamnosus, E. faecium, S. salivarius subspecies thermophilus, Aspergillus oryza, and Candida pintolopesii plus Zinc | Enhances growth performance, better feed utilization, increase in villus height in the duodenum and ileum | Chicken (broiler) | [159] |
| Synbiotics A: Enterococcus sp., Pediococcus sp., Bifidobacterium sp., Lactobacillus sp. plus fructooligosaccharides Synbiotic B: L. acidophilus, L. casei, L. salivarius, L. plantarum, L. rhamnosus, L. brevis, B. bifidum, B. lactis, S. thermophilus, prebiotic inulin (chicory root extract), protease, amylase, cellulase, hemicellulase, lipase, papain and bromelain | Modulate the caecal microbiota without any effects on Salmonella Typhimurium shedding | Chickens (layers) | [65] |
| Probiotics; (L. rhamnosus, L. casei L. plantarum B. animalis) prebiotics (383 mg of fructooligosaccharides and 100 mg of galactooligosaccharides) | Improved gastrointestinal complications, sepsis, and mortality in premature infants | Preterm infants | [157] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwoji, I.D.; Aiyegoro, O.A.; Okpeku, M.; Adeleke, M.A. Multi-Strain Probiotics: Synergy among Isolates Enhances Biological Activities. Biology 2021, 10, 322. https://doi.org/10.3390/biology10040322
Kwoji ID, Aiyegoro OA, Okpeku M, Adeleke MA. Multi-Strain Probiotics: Synergy among Isolates Enhances Biological Activities. Biology. 2021; 10(4):322. https://doi.org/10.3390/biology10040322
Chicago/Turabian StyleKwoji, Iliya D., Olayinka A. Aiyegoro, Moses Okpeku, and Matthew A. Adeleke. 2021. "Multi-Strain Probiotics: Synergy among Isolates Enhances Biological Activities" Biology 10, no. 4: 322. https://doi.org/10.3390/biology10040322
APA StyleKwoji, I. D., Aiyegoro, O. A., Okpeku, M., & Adeleke, M. A. (2021). Multi-Strain Probiotics: Synergy among Isolates Enhances Biological Activities. Biology, 10(4), 322. https://doi.org/10.3390/biology10040322