
Capitoline Dolphins: Residency Patterns and Abundance Estimate of Tursiops truncatus at the Tiber River Estuary (Mediterranean Sea)

 ,
,  ,
,  , , , ,
, , , ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
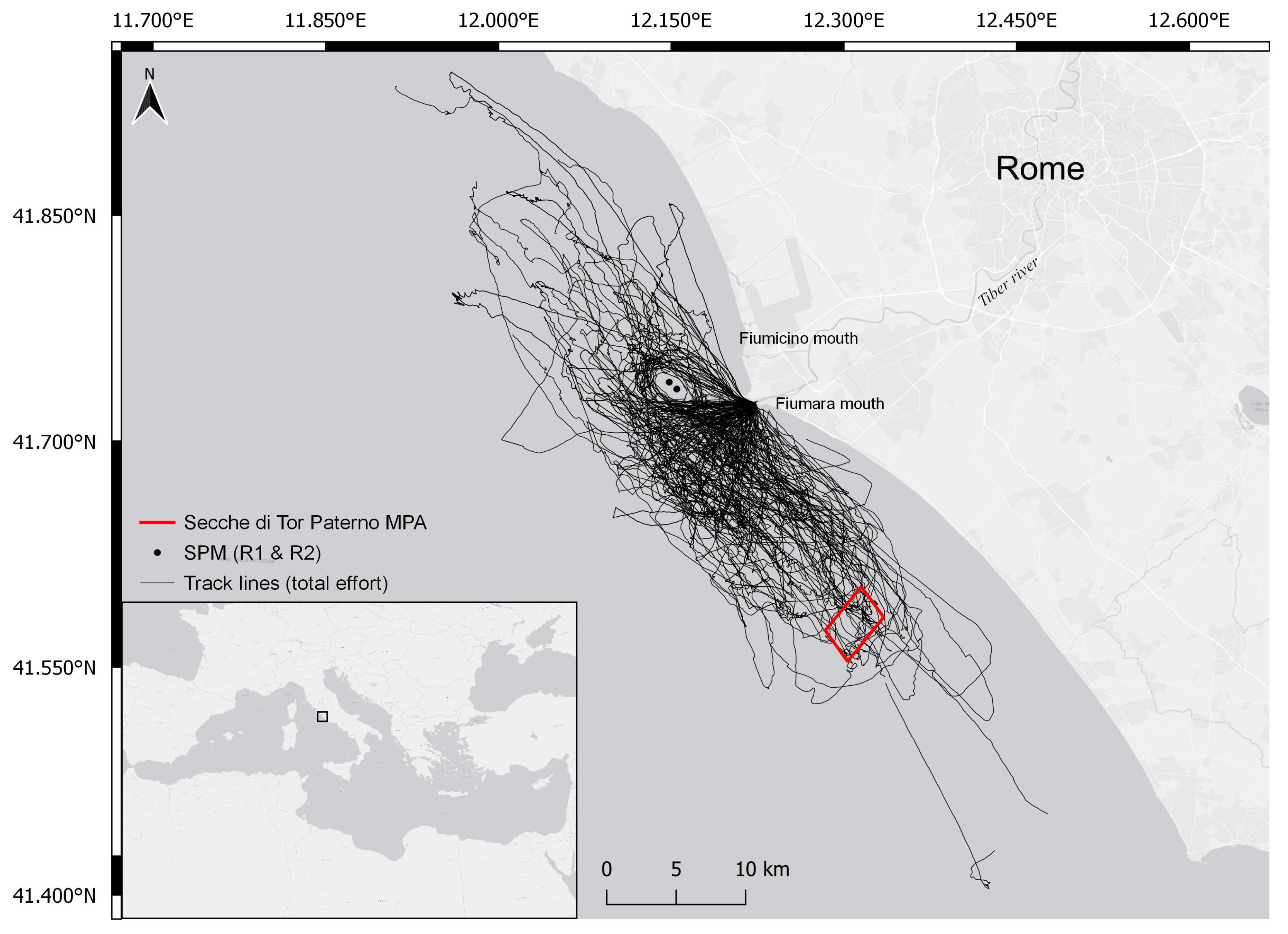
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis
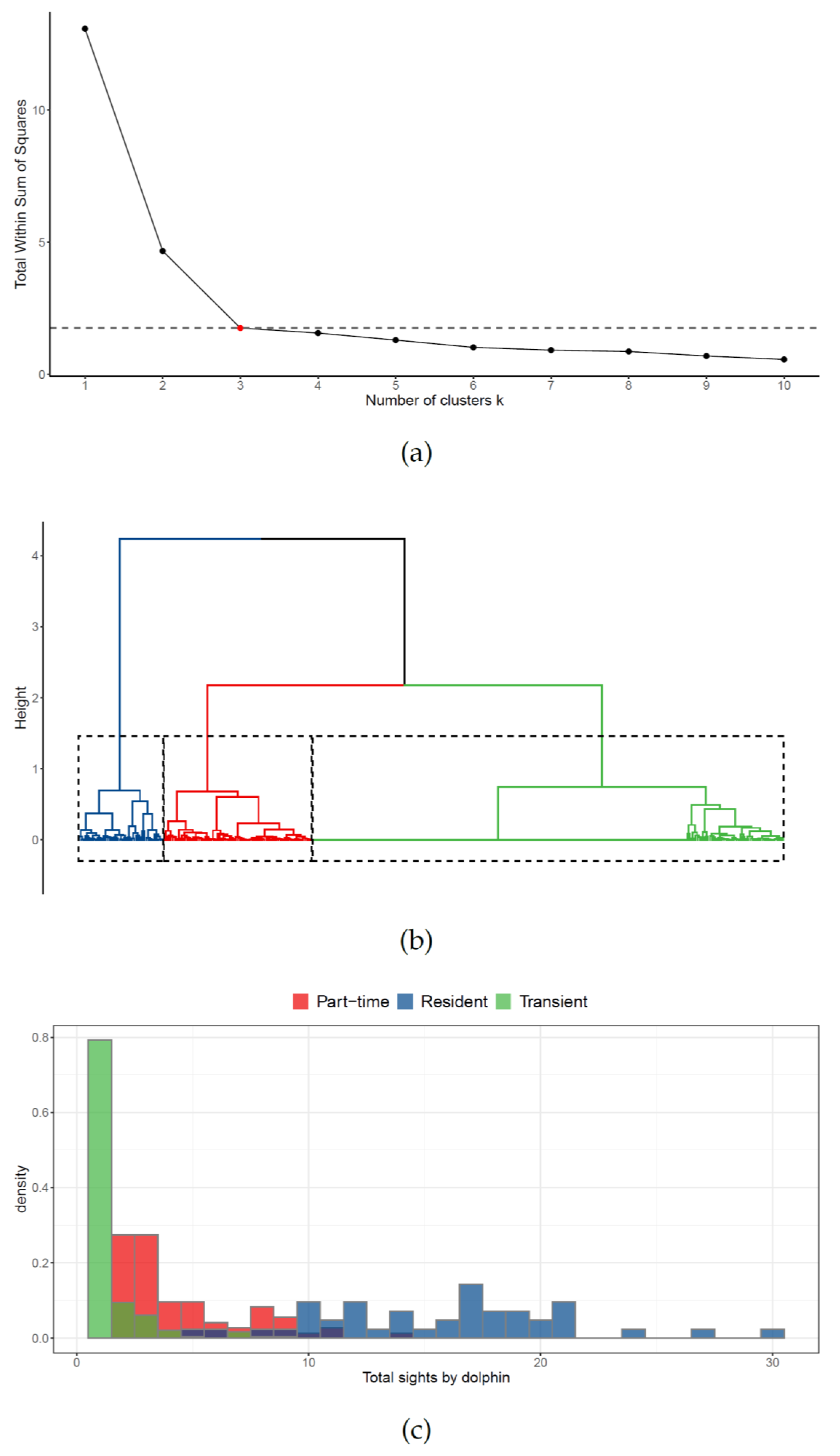
2.3.1. Photo-Identification
2.3.2. Site Fidelity
- Occurrence (Occ): The proportion of captures, determined by the number of times an individual was captured divided by the total number of capture occasions.
- Monthly rate (MR): Monthly average number of sights.
- Yearly rate (YR): Yearly average number of sights.
- Resight rate (RR): The average number of recaptures over all the recapture occasions.
- Periodicity (P): The recurrence of an individual, determined by the inverse of the average time between successive recaptures.
- Relative span-time (RS): The portion of the whole observation time elapsed between the first and last captures of the individual.
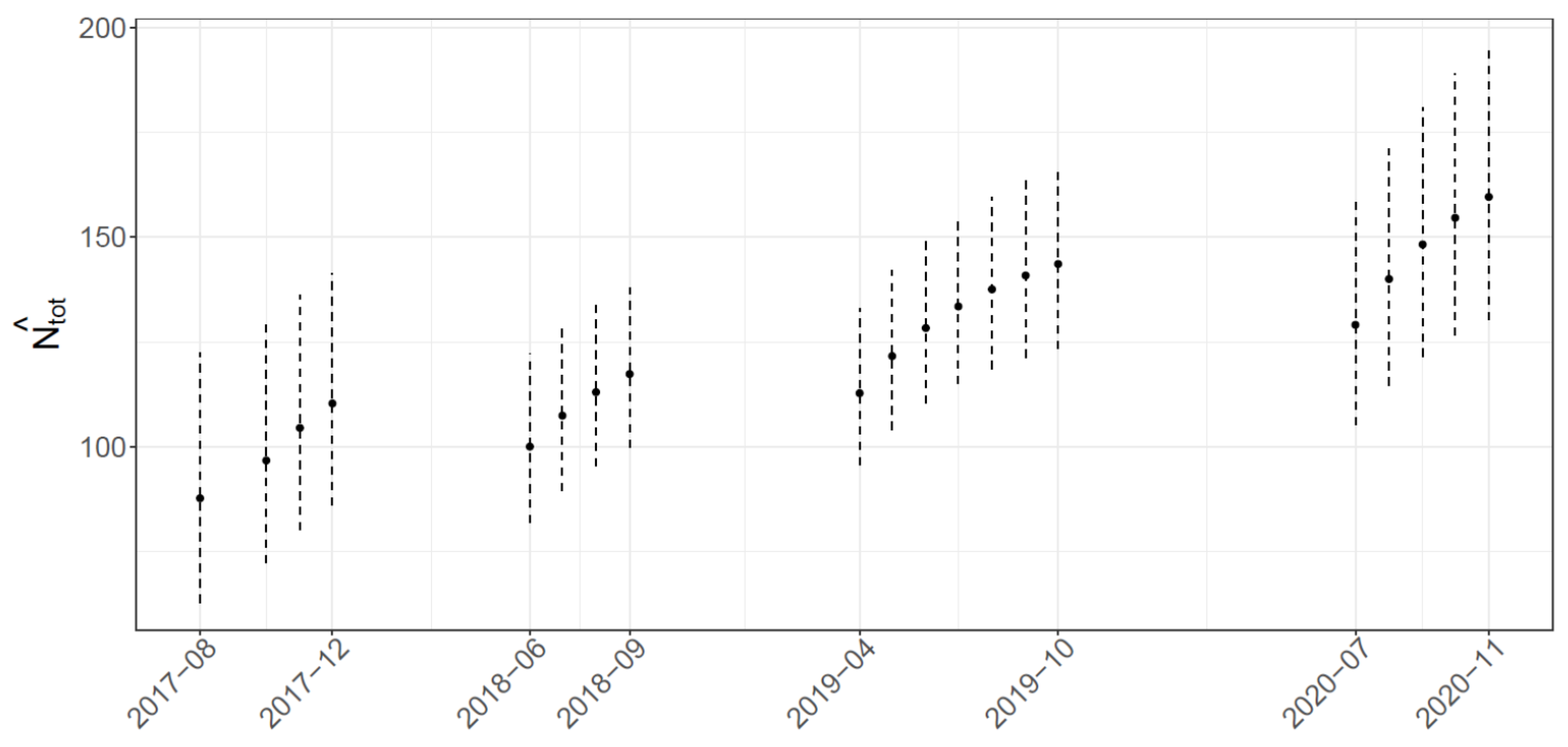
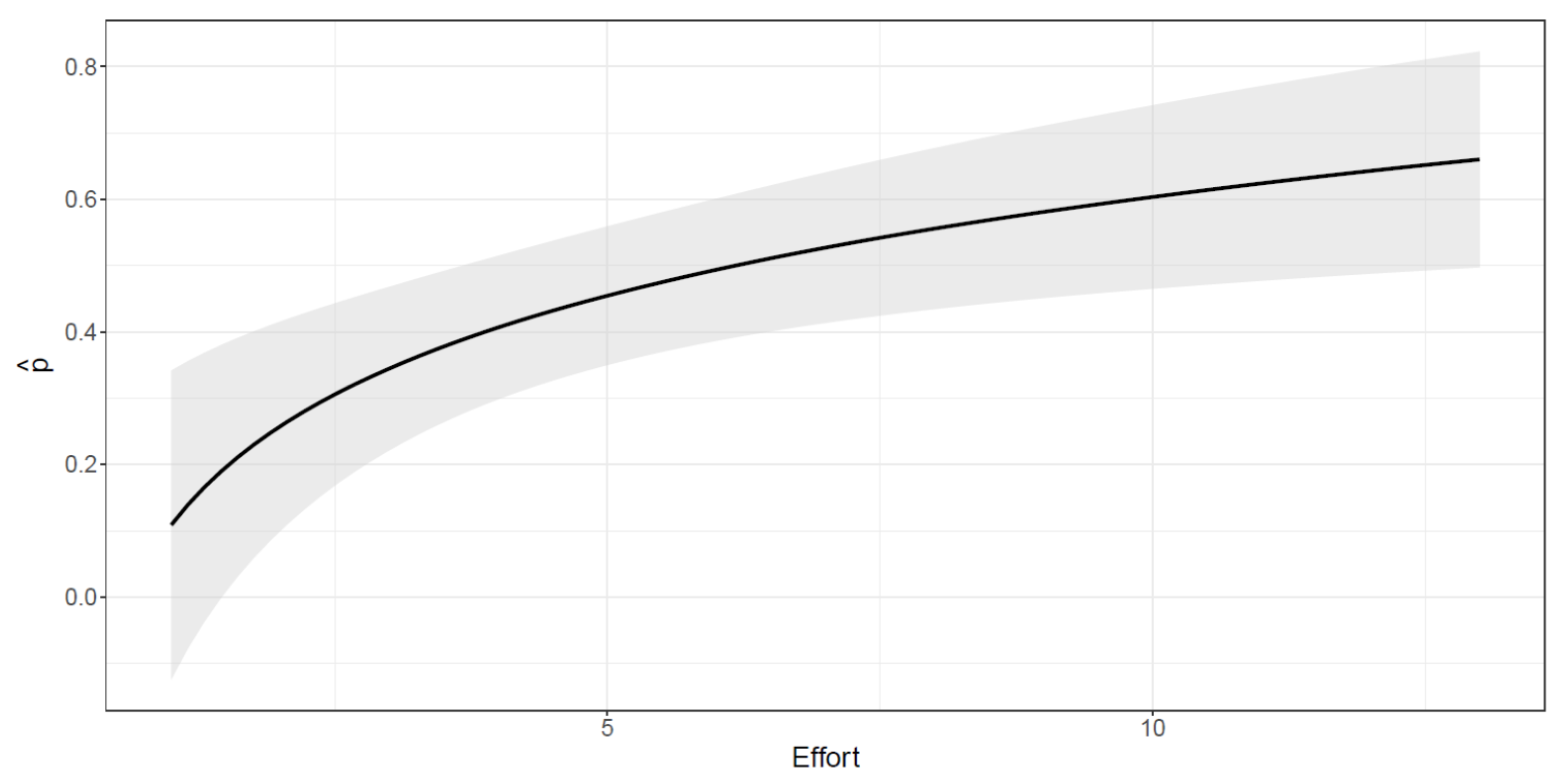
2.3.3. Abundance Estimate
- Captures are independent across individuals and along time.
- Both capture probability pj (i.e., detectability) and survival probability φj on each occasion are homogeneous among all individuals [62].
- Each individual from the superpopulation N enters the study area on occasion j with the same probability βj; then, it stays in the study area as long as it survives, with no chance of re-entrance once it has exited.
- , the capture probabilities of each occasion.
- , the survival probability between two subsequent sampling occasions; as migration/immigration phenomena are not distinguishable from natality/mortality, φ represents the apparent survival probability.
- , the probability that an individual from the superpopulation will join the population between two subsequent sampling occasions.
- Survival probability between two subsequent sampling occasions:
- −
- Constant: It does not vary across groups or over time: φ.
- −
- Occasion: It does not vary across groups but can vary over time: φt, t = 1, …, nm.
- −
- Group: It does not vary over time but can vary by group: φg, g = 1, …, G.
- Detectability at each sampling occasion:
- −
- Constant: It does not vary across groups or over time: p.
- −
- Log-effort: It does not vary over time but depends linearly on the logarithm of the effort: pl e f f, where l e f f = log(effort) and effort ∈
- −
- Occasion: It does not vary across groups but can vary over time: pt, t = 1, …, nm.
- Entry probability between two subsequent sampling occasions:
- −
- Constant: It does not vary across groups or over time: β.
- −
- Occasion: It does not vary across groups but can vary over time: βt, t = 1, …, nm.
- −
- Group: It does not vary over time but can vary by group: βg, g = 1, …, G.
- Marks are not lost over time, and individuals are identified with no error [63].
- Sampling is simultaneous, and each individual is released just after.
- Captures do not affect the detectability and survival probability of the captured individuals after release.
3. Results
3.1. Site Fidelity
3.2. Abundance Estimates
4. Discussion
4.1. Site Fidelity
4.2. Abundance Estimates
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berlow, E.L. Strong effects of weak interactions in ecological communities. Nature 1999, 398, 330–334. [Google Scholar] [CrossRef]
- Springer, A.M.; Estes, J.A.; Van Vliet, G.B.; Williams, T.M.; Doak, D.F.; Danner, E.M.; Forney, K.A.; Pfister, B. Sequential megafaunal collapse in the North Pacific Ocean: An ongoing legacy of industrial whaling? Proc. Natl. Acad. Sci. USA 2003, 100, 12223–12228. [Google Scholar] [CrossRef] [PubMed]
- Luypaert, T.; Hagan, J.G.; McCarthy, M.L.; Poti, M. Status of marine biodiversity in the Anthropocene. In YOUMARES 9-The Oceans: Our Research, Our Future; Springer: Cham, Switzerland, 2020; pp. 57–82. [Google Scholar]
- Parra, G.J. Resource partitioning in sympatric delphinids: Space use and habitat preferences of Australian snubfin and Indo-Pacific humpback dolphins. J. Anim. Ecol. 2006, 75, 862–874. [Google Scholar] [CrossRef]
- Bejder, L.; Hodgson, A.; Loneragan, N.; Allen, S. Coastal dolphins in north-western Australia: The need for re-evaluation of species listings and shortcomings in the Environmental Impact Assessment process. Pac. Conserv. Biol. 2012, 18, 22–25. [Google Scholar] [CrossRef]
- Pulcini, M.; Pace, D.S.; La Manna, G.; Triossi, F.; Fortuna, C.M. Distribution and abundance estimates of bottlenose dolphins (Tursiops truncatus) around Lampedusa Island (Sicily Channel, Italy): Implications for their management. J. Mar. Biol. Assoc. UK 2014, 94, 1175–1184. [Google Scholar] [CrossRef]
- Pace, D.S.; Mussi, B.; Gordon, J.C.; Wurtz, M. Foreword. Aquat. Conserv. Mar. Freshw. Ecosyst. 2014, 24, 1–3. [Google Scholar] [CrossRef]
- Pace, D.S.; Miragliuolo, A.; Mariani, M.; Vivaldi, C.; Mussi, B. Sociality of sperm whale off Ischia Island (Tyrrhenian Sea, Italy). Aquat. Conserv. Mar. Freshw. Ecosyst. 2014, 24, 71–82. [Google Scholar] [CrossRef]
- Pollock, K.H.; Nichols, J.D.; Simons, T.R.; Farnsworth, G.L.; Bailey, L.L.; Sauer, J.R. Large scale wildlife monitoring studies: Statistical methods for design and analysis. Environmetrics 2002, 13, 105–119. [Google Scholar] [CrossRef]
- Haughey, R.; Hunt, T.; Hanf, D.; Rankin, R.W.; Parra, G.J. Photographic capture-recapture analysis reveals a large population of Indo-Pacific bottlenose dolphins (Tursiops aduncus) with low site fidelity off the North West Cape, Western Australia. Front. Mar. Sci. 2020, 6, 781. [Google Scholar] [CrossRef]
- Williams, B.K.; Nichols, J.D.; Conroy, M.J. Analysis and Management of Animal Populations; Academic Press: Cambridge, MA, USA, 2002. [Google Scholar]
- Greenwood, P.J. Mating systems, philopatry and dispersal in birds and mammals. Anim. Behav. 1980, 28, 1140–1162. [Google Scholar] [CrossRef]
- Tschopp, A.; Ferrari, M.A.; Crespo, E.A.; Coscarella, M.A. Development of a site fidelity index based on population capture- recapture data. PeerJ 2018, 6, e4782. [Google Scholar] [CrossRef]
- Zanardo, N.; Parra, G.J.; Möller, L.M. Site fidelity, residency, and abundance of bottlenose dolphins (Tursiops sp.) in Adelaide’s coastal waters, South Australia. Mar. Mammal Sci. 2016, 32, 1381–1401. [Google Scholar] [CrossRef]
- Möller, L.; Allen, S.; Harcourt, R. Group characteristics, site fidelity and seasonal abundance of bottlenosed dolphins (Tursiops aduncus) in Jervis Bay and Port Stephens, South-Eastern Australia. Aust. Mammal 2002, 24, 11–22. [Google Scholar] [CrossRef]
- Foote, A.D.; Similä, T.; Víkingsson, G.A.; Stevick, P.T. Movement, site fidelity and connectivity in a top marine predator, the killer whale. Evol. Ecol. 2010, 24, 803–814. [Google Scholar] [CrossRef]
- Clavel, J.; Robert, A.; Devictor, V.; Julliard, R. Abundance estimation with a transient model under the robust design. J. Wildl. Manag. 2008, 72, 1203–1210. [Google Scholar] [CrossRef]
- Conn, P.B.; Gorgone, A.M.; Jugovich, A.R.; Byrd, B.L.; Hansen, L.J. Accounting for transients when estimating abundance of bottlenose dolphins in Choctawhatchee Bay, Florida. J. Wildl. Manag. 2011, 75, 569–579. [Google Scholar] [CrossRef]
- Zanardo, N.; Parra, G.J.; Passadore, C.; Möller, L.M. Ensemble modelling of southern Australian bottlenose dolphin Tursiops sp. distribution reveals important habitats and their potential ecological function. Mar. Ecol. Prog. Ser. 2017, 569, 253–266. [Google Scholar] [CrossRef]
- La Manna, G.; Ronchetti, F.; Sarà, G.; Ruiu, A.; Ceccherelli, G. Common Bottlenose Dolphin Protection and Sustainable Boating: Species Distribution Modeling for Effective Coastal Planning. Front. Mar. Sci. 2020, 7, 955. [Google Scholar] [CrossRef]
- La Manna, G.; Ronchetti, F.; Sarà, G. Predicting common bottlenose dolphin habitat preference to dynamically adapt management measures from a Marine Spatial Planning perspective. Ocean. Coast. Manag. 2016, 130, 317–327. [Google Scholar] [CrossRef]
- Marini, C.; Fossa, F.; Paoli, C.; Bellingeri, M.; Gnone, G.; Vassallo, P. Predicting bottlenose dolphin distribution along Liguria coast (northwestern Mediterranean Sea) through different modeling techniques and indirect predictors. J. Environ. Manag. 2015, 150, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Azzellino, A.; Panigada, S.; Lanfredi, C.; Zanardelli, M.; Airoldi, S.; di Sciara, G.N. Predictive habitat models for managing marine areas: Spatial and temporal distribution of marine mammals within the Pelagos Sanctuary (Northwestern Mediterranean Sea). Ocean. Coast. Manag. 2012, 67, 63–74. [Google Scholar] [CrossRef]
- Bonizzoni, S.; Furey, N.B.; Santostasi, N.L.; Eddy, L.; Valavanis, V.D.; Bearzi, G. Modelling dolphin distribution within an Important Marine Mammal Area in Greece to support spatial management planning. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 1665–1680. [Google Scholar] [CrossRef]
- Hammond, P.; Bearzi, G.; Bjørge, A.; Forney, K.; Karkzmarski, L.; Kasuya, T.; Perrin, W.; Scott, M.; Wang, J.; Wells, R.; et al. Tursiops truncatus. The IUCN Red List of Threatened Species 2012: e. T22563A17347397. 2018. Available online: https://www.iucnredlist.org/species/22563/2782611 (accessed on 12 February 2021).
- Pace, D.S.; Giacomini, G.; Campana, I.; Paraboschi, M.; Pellegrino, G.; Silvestri, M.; Alessi, J.; Angeletti, D.; Cafaro, V.; Pavan, G.; et al. An integrated approach for cetacean knowledge and conservation in the central Mediterranean Sea using research and social media data sources. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 1302–1323. [Google Scholar] [CrossRef]
- Montuori, P.; Aurino, S.; Garzonio, F.; Sarnacchiaro, P.; Polichetti, S.; Nardone, A.; Triassi, M. Estimates of Tiber River organophosphate pesticide loads to the Tyrrhenian Sea and ecological risk. Sci. Total Environ. 2016, 559, 218–231. [Google Scholar] [CrossRef] [PubMed]
- Montuori, P.; Aurino, S.; Garzonio, F.; Nardone, A.; Triassi, M. Estimation of heavy metal loads from Tiber River to the Tyrrhenian Sea and environmental quality assessment. Environ. Sci. Pollut. Res. 2016, 23, 23694–23713. [Google Scholar] [CrossRef] [PubMed]
- Ventura, D.; Jona Lasinio, G.; Ardizzone, G. Temporal partitioning of microhabitat use among four juvenile fish species of the genus Diplodus (Pisces: Perciformes, Sparidae). Mar. Ecol. 2015, 36, 1013–1032. [Google Scholar] [CrossRef]
- Ardizzone, G.; Belluscio, A.; Criscoli, A. Atlante degli Habitat dei Fondali Marini del Lazio; Dipartimento di Biologia Ambientale, Sapienza Università di Roma, Sapienza Università Editrice: Roma, Italy, 2018. [Google Scholar]
- Bonifazi, A.; Lezzi, M.; Ventura, D.; Lisco, S.; Cardone, F.; Gravina, M.F. Macrofaunal biodiversity associated with different developmental phases of a threatened Mediterranean Sabellaria alveolata (Linnaeus, 1767) reef. Mar. Environ. Res. 2019, 145, 97–111. [Google Scholar] [CrossRef]
- Casoli, E.; Bonifazi, A.; Ardizzone, G.; Gravina, M.F.; Russo, G.F.; Sandulli, R.; Donnarumma, L. Comparative Analysis of Mollusc Assemblages from Different Hard Bottom Habitats in the Central Tyrrhenian Sea. Diversity 2019, 11, 74. [Google Scholar] [CrossRef]
- Triossi, F.; Willis, T.J.; Pace, D.S. Occurrence of bottlenose dolphins Tursiops truncatus in natural gas fields of the northwestern Adriatic Sea. Mar. Ecol. 2013, 34, 373–379. [Google Scholar] [CrossRef]
- Blanco, C.; Salomón, O.; Raga, J. Diet of the bottlenose dolphin (Tursiops truncatus) in the western Mediterranean Sea. Mar. Biol. Assoc. UK J. Mar. Biol. Assoc. UK 2001, 81, 1053–1058. [Google Scholar] [CrossRef]
- Pace, D.S.; Pulcini, M.; Triossi, F. Anthropogenic food patches and association patterns of Tursiops truncatus at Lampedusa island, Italy. Behav. Ecol. 2012, 23, 254–264. [Google Scholar] [CrossRef]
- Bonizzoni, S.; Furey, N.B.; Bearzi, G. Bottlenose dolphins (Tursiops truncatus) in the north-western Adriatic Sea: Spatial distribution and effects of trawling. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 635–650. [Google Scholar] [CrossRef]
- Mann, J. Behavioral sampling methods for cetaceans: A review and critique. Mar. Mammal Sci. 1999, 15, 102–122. [Google Scholar] [CrossRef]
- Gowans, S.; Whitehead, H. Photographic identification of northern bottlenose whales (Hyperoodon ampullatus): Sources of heterogeneity from natural marks. Mar. Mammal Sci. 2001, 17, 76–93. [Google Scholar] [CrossRef]
- Nicholson, K.; Bejder, L.; Allen, S.J.; Krützen, M.; Pollock, K.H. Abundance, survival and temporary emigration of bottlenose dolphins (Tursiops sp.) off Useless Loop in the western gulf of Shark Bay, Western Australia. Mar. Freshw. Res. 2012, 63, 1059–1068. [Google Scholar] [CrossRef]
- Wursig, B.; Jefferson, T.A. Methods of photo-identification for small cetaceans. Rep. Int. Whal. Comm. 1990, 12, 43–52. [Google Scholar]
- Urian, K.; Gorgone, A.; Read, A.; Balmer, B.; Wells, R.S.; Berggren, P.; Durban, J.; Eguchi, T.; Rayment, W.; Hammond, P.S. Recommendations for photo-identification methods used in capture-recapture models with cetaceans. Mar. Mammal Sci. 2015, 31, 298–321. [Google Scholar] [CrossRef]
- Mariani, M.; Miragliuolo, A.; Mussi, B.; Russo, G.F.; Ardizzone, G.; Pace, D.S. Analysis of the natural markings of Risso’s dolphins (Grampus griseus) in the central Mediterranean Sea. J. Mammal 2016, 97, 1512–1524. [Google Scholar] [CrossRef]
- Pollock, K.H.; Nichols, J.D.; Brownie, C.; Hines, J.E. Statistical inference for capture-recapture experiments. Wildl. Monogr. 1990, 107, 3–97. [Google Scholar]
- Tyne, J.A.; Pollock, K.H.; Johnston, D.W.; Bejder, L. Abundance and survival rates of the Hawai’i Island associated spinner dolphin (Stenella longirostris) stock. PLoS ONE 2014, 9, e86132. [Google Scholar] [CrossRef]
- Morteo, E.; Rocha-Olivares, A.; Morteo, R. Sensitivity analysis of residency and site fidelity estimations to variations in sampling effort and individual catchability. Rev. Mex. Biodivers. 2012, 83, 487–495. [Google Scholar] [CrossRef]
- Balmer, B.; Wells, R.; Nowacek, S.; Nowacek, D.; Schwacke, L.; McLellan, W.; Scharf, F. 157 Seasonal abundance and distribution patterns of common bottlenose dolphins (Tursiops truncatus) near St. Joseph Bay, Florida, USA. J. Cetacean Res. Manag. 2008, 10, 157–167. [Google Scholar]
- Simões-Lopes, P.C.; Fabian, M.E. Residence patterns and site fidelity in bottlenose dolphins, Tursiops truncatus (Montagu) (Cetacea, Delphinidae) off Southern Brazil. Rev. Bras. Zool. 1999, 16, 1017–1024. [Google Scholar] [CrossRef]
- Gower, J.C. A general coefficient of similarity and some of its properties. Biometrics 1971, 27, 857–871. [Google Scholar] [CrossRef]
- Hennig, C. Clustering strategy and method selection. arXiv 2015, arXiv:1503.02059. [Google Scholar]
- Murtagh, F.; Legendre, P. Ward’s hierarchical agglomerative clustering method: Which algorithms implement Ward’s criterion? J. Classif. 2014, 31, 274–295. [Google Scholar] [CrossRef]
- Allaire, J. RStudio: Integrated Development Environment for R; RStudio: Boston, MA, USA, 2012; Volume 770, p. 394. [Google Scholar]
- Stanley, T.R.; Richards, J.D. Software Review: A program for testing capture-recapture data for closure. Wildl. Soc. Bull. 2005, 33, 782–785. [Google Scholar] [CrossRef]
- Stanley, T.R.; Burnham, K.P. A closure test for time-specific capture-recapture data. Environ. Ecol. Stat. 1999, 6, 197–209. [Google Scholar] [CrossRef]
- Otis, D.L.; Burnham, K.P.; White, G.C.; Anderson, D.R. Statistical inference from capture data on closed animal populations. Wildl. Monogr. 1978, 62, 3–135. [Google Scholar]
- Arnason, A.N.; Schwarz, C.J. POPAN-4: Enhancements to a system for the analysis of mark-recapture data from open populations. J. Appl. Stat. 1995, 22, 785–800. [Google Scholar] [CrossRef]
- Schwarz, C.J.; Arnason, A.N. A general methodology for the analysis of capture-recapture experiments in open populations. Biometrics 1996, 52, 860–873. [Google Scholar] [CrossRef]
- Félix, F.; Calderón, A.; Vintimilla, M.; Bayas-Rea, R.A. Decreasing population trend in coastal bottlenose dolphin (Tursiops truncatus) from the Gulf of Guayaquil, Ecuador. Aquat. Conserv. Mar. Freshw. Ecosyst. 2017, 27, 856–866. [Google Scholar] [CrossRef]
- Papale, E.; Ceraulo, M.; Giardino, G.; Buffa, G.; Filiciotto, F.; Grammauta, R.; Maccarrone, V.; Mazzola, S.; Buscaino, G. Association patterns and population dynamics of bottlenose dolphins in the Strait of Sicily (Central Mediterranean Sea): Implication for management. Popul. Ecol. 2017, 59, 55–64. [Google Scholar] [CrossRef]
- Bearzi, G.; Bonizzoni, S.; Riley, M.A.; Santostasi, N.L. Bottlenose dolphins in the north-western Adriatic Sea: Abundance and management implications. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 651–664. [Google Scholar] [CrossRef]
- Nichols, J. Modern open-population capture-recapture models. In Handbook of Capture-Recapture Analysis; Princeton University Press: Princeton, NJ, USA, 2005; pp. 88–123. [Google Scholar]
- Reisinger, R.; Karczmarski, L. Population size estimate of Indo-Pacific bottlenose dolphins in the Algoa Bay region, South Africa. Mar. Mammal Sci. 2010, 26, 86–97. [Google Scholar] [CrossRef]
- Schwarz, C.J.; Seber, G.A. Estimating animal abundance: Review III. Stat. Sci. 1999, 14, 427–456. [Google Scholar] [CrossRef]
- Friday, N.A.; Smith, T.D.; Stevick, P.T.; Allen, J.; Fernald, T. Balancing bias and precision in capture-recapture estimates of abundance. Mar. Mammal Sci. 2008, 24, 253–275. [Google Scholar] [CrossRef]
- Anderson, D.; Burnham, K.; White, G. AIC model selection in overdispersed capture-recapture data. Ecology 1994, 75, 1780–1793. [Google Scholar] [CrossRef]
- Lebreton, J.; Burnham, K.; Clobert, J.; Anderson, D. Modeling survival and testing biological hypotheses using marked animals: A unified approach with case studies. Ecol. Monogr. 1992, 62, 67–118. [Google Scholar] [CrossRef]
- Cooch, E.; White, G. Program MARK: A Gentle Introduction; 13th ed.; Colorado State University: Fort Collins, CO, USA, 2014. [Google Scholar]
- Wilson, B.; Hammond, P.S.; Thompson, P.M. Estimating size and assessing trends in a coastal bottlenose dolphin population. Ecol. Appl. 1999, 9, 288–300. [Google Scholar] [CrossRef]
- Burham, K.; Anderson, D.; White, G.; Brownie, C.; Pollock, K. Design and analysis of fish survival experiments based on release-recapture data. Am. Fish. Soc. Monogr. 1987, 5, 1–437. [Google Scholar]
- Defran, R.; Weller, D.W. Occurrence, distribution, site fidelity, and school size of bottlenose dolphins (Tursiops truncatus) off San Diego, California. Mar. Mammal Sci. 1999, 15, 366–380. [Google Scholar] [CrossRef]
- Wells, R.S.; Schwacke, L.H.; Rowles, T.K.; Balmer, B.C.; Zolman, E.; Speakman, T.; Townsend, F.I.; Tumlin, M.C.; Barleycorn, A.; Wilkinson, K.A. Ranging patterns of common bottlenose dolphins Tursiops truncatus in Barataria Bay, Louisiana, following the Deepwater Horizon oil spill. Endanger. Species Res. 2017, 33, 159–180. [Google Scholar] [CrossRef]
- Sprogis, K.R.; Raudino, H.C.; Rankin, R.; MacLeod, C.D.; Bejder, L. Home range size of adult Indo-Pacific bottlenose dolphins (Tursiops aduncus) in a coastal and estuarine system is habitat and sex-specific. Mar. Mammal Sci. 2016, 32, 287–308. [Google Scholar] [CrossRef]
- Passadore, C.; Möller, L.; Diaz-Aguirre, F.; Parra, G.J. High site fidelity and restricted ranging patterns in southern Australian bottlenose dolphins. Ecol. Evol. 2018, 8, 242–256. [Google Scholar] [CrossRef]
- Silva, M.A.; Magalhães, S.; Prieto, R.; Santos, R.S.; Hammond, P.S. Estimating survival and abundance in a bottlenose dolphin population taking into account transience and temporary emigration. Mar. Ecol. Prog. Ser. 2009, 392, 263–276. [Google Scholar] [CrossRef]
- Brown, A.M.; Bejder, L.; Pollock, K.H.; Allen, S.J. Site-specific assessments of the abundance of three inshore dolphin species to inform conservation and management. Front. Mar. Sci. 2016, 3, 4. [Google Scholar] [CrossRef]
- Fury, C.A.; Harrison, P.L. Abundance, site fidelity and range patterns of Indo-Pacific bottlenose dolphins (Tursiops aduncus) in two Australian subtropical estuaries. Mar. Freshw. Res. 2008, 59, 1015–1027. [Google Scholar] [CrossRef]
- Rossman, S.; Ostrom, P.H.; Stolen, M.; Barros, N.B.; Gandhi, H.; Stricker, C.A.; Wells, R.S. Individual specialization in the foraging habits of female bottlenose dolphins living in a trophically diverse and habitat rich estuary. Oecologia 2015, 178, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Lidgard, D.; Bowen, W.; Iverson, S. Sex-differences in fine-scale home-range use in an upper-trophic level marine predator. Mov. Ecol. 2020, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Engelhaupt, D.; Rus Hoelzel, A.; Nicholson, C.; Frantzis, A.; Mesnick, S.; Gero, S.; Whitehead, H.; Rendell, L.; Miller, P.; De Stefanis, R.; et al. Female philopatry in coastal basins and male dispersion across the North Atlantic in a highly mobile marine species, the sperm whale (Physeter macrocephalus). Mol. Ecol. 2009, 18, 4193–4205. [Google Scholar] [CrossRef]
- Servidio, A.; Pérez-Gil, E.; Pérez-Gil, M.; Cañadas, A.; Hammond, P.S.; Martín, V. Site fidelity and movement patterns of short-finned pilot whales within the Canary Islands: Evidence for resident and transient populations. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 227–241. [Google Scholar] [CrossRef]
- Wilson, B.; Reid, J.R.; Grellier, K.; Thompson, P.M.; Hammond, P.S. Considering the temporal when managing the spatial: A population range expansion impacts protected area-based management for bottlenose dolphins. Anim. Conserv. 2004, 7, 331–338. [Google Scholar] [CrossRef]
- Lusseau, D.; Wilson, B.; Hammond, P.S.; Grellier, K.; Durban, J.W.; Parsons, K.M.; Barton, T.R.; Thompson, P.M. Quantifying the influence of sociality on population structure of bottlenose dolphins. J. Anim. Ecol. 2006, 75, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Watson-Capps, J.J.; Mann, J. The effects of aquaculture on bottlenose dolphin (Tursiops sp.) ranging in Shark Bay, Western Australia. Biol. Conserv. 2005, 124, 519–526. [Google Scholar] [CrossRef]
- Pleslić, G.; Rako Gospić, N.; Mackelworth, P.; Wiemann, A.; Holcer, D.; Fortuna, C. The abundance of common bottlenose dolphins (Tursiops truncatus) in the former special marine reserve of the Cres-Lošinj Archipelago, Croatia. Aquat. Conserv. Mar. Freshw. Ecosyst. 2015, 25, 125–137. [Google Scholar] [CrossRef]
- Tezanos-Pinto, G.; Constantine, R.; Brooks, L.; Jackson, J.A.; Mourão, F.; Wells, S.; Scott Baker, C. Decline in local abundance of bottlenose dolphins (Tursiops truncatus) in the Bay of Islands, New Zealand. Mar. Mammal Sci. 2013, 29, 390–410. [Google Scholar] [CrossRef]
- Berrow, S.; O’Brien, J.; Groth, L.; Foley, A.; Voigt, K. Abundance estimate of bottlenose dolphins (Tursiops truncatus) in the Lower River Shannon candidate Special Area of Conservation, Ireland. Aquat. Mamm. 2012, 38, 136–144. [Google Scholar] [CrossRef]
- Pleslić, G.; Rako-Gospić, N.; Holcer, D. Bottlenose dolphins (Tursiops truncatus) in North Dalmatia, Croatia: Occurrence and demographic parameters. Mar. Mammal Sci. 2021, 37, 142–161. [Google Scholar] [CrossRef]
- Balmer, B.; Watwood, S.; Quigley, B.; Speakman, T.; Barry, K.; Mullin, K.; Rosel, P.; Sinclair, C.; Zolman, E.; Schwacke, L. Common bottlenose dolphin (Tursiops truncatus) abundance and distribution patterns in St Andrew Bay, Florida, USA. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 486–498. [Google Scholar] [CrossRef]
- Santostasi, N.L.; Bonizzoni, S.; Bearzi, G.; Eddy, L.; Gimenez, O. A robust design capture-recapture analysis of abundance, survival and temporary emigration of three Odontocete species in the Gulf of Corinth, Greece. PLoS ONE 2016, 11, e0166650. [Google Scholar] [CrossRef]
- Bearzi, G.; Agazzi, S.; Gonzalvo, J.; Bonizzoni, S.; Costa, M.; Petroselli, A. Biomass removal by dolphins and fisheries in a Mediterranean Sea coastal area: Do dolphins have an ecological impact on fisheries? Aquat. Conserv. Mar. Freshw. Ecosyst. 2010, 20, 549–559. [Google Scholar] [CrossRef]
- Gnone, G.; Bellingeri, M.; Dhermain, F.; Dupraz, F.; Nuti, S.; Bedocchi, D.; Moulins, A.; Rosso, M.; Alessi, J.; McCrea, R.S.; et al. Distribution, abundance, and movements of the bottlenose dolphin (Tursiops truncatus) in the Pelagos Sanctuary MPA (north-west Mediterranean Sea). Aquat. Conserv. Mar. Freshw. Ecosyst. 2011, 21, 372–388. [Google Scholar] [CrossRef]
- Bejder, L.; Samuels, A.; Whitehead, H.; Gales, N.; Mann, J.; Connor, R.; Heithaus, M.; Watson-Capps, J.; Flaherty, C.; Krützen, M. Decline in relative abundance of bottlenose dolphins exposed to long-term disturbance. Conserv. Biol. 2006, 20, 1791–1798. [Google Scholar] [CrossRef]
- Speakman, T.R.; Lane, S.M.; Schwacke, L.H.; Fair, P.A.; Zolman, E.S. Mark-recapture estimates of seasonal abundance and survivorship for bottlenose dolphins (Tursiops truncatus) near Charleston, South Carolina, USA. J. Cetacean Res. Manag. 2010, 11, 153–162. [Google Scholar]
- Smith, H.C.; Pollock, K.; Waples, K.; Bradley, S.; Bejder, L. Use of the robust design to estimate seasonal abundance and demographic parameters of a coastal bottlenose dolphin (Tursiops truncatus) population. PLoS ONE 2013, 8, e76574. [Google Scholar] [CrossRef]
- Gregorietti, M.; Papale, E.; Ceraulo, M.; de Vita, C.; Pace, D.S.; Tranchida, G.; Mazzola, S.; Buscaino, G. Acoustic Presence of Dolphins through Whistles Detection in Mediterranean Shallow Waters. J. Mar. Sci. Eng. 2021, 9, 78. [Google Scholar] [CrossRef]
- March, D.; Metcalfe, K.; Tintoré, J.; Godley, B. Tracking the Global Reduction of Marine Traffic during the COVID-19 Pandemic. 2020. Available online: https://www.researchsquare.com/article/rs-47243/v1 (accessed on 30 January 2021).
- Millefiori, L.M.; Braca, P.; Zissis, D.; Spiliopoulos, G.; Marano, S.; Willett, P.K.; Carniel, S. Covid-19 Impact on Global Maritime Mobility. Available online: https://arxiv.org/abs/2009.06960v2 (accessed on 30 January 2021).
- Thomson, D.J.; Barclay, D.R. Real-time observations of the impact of COVID-19 on underwater noise. J. Acoust. Soc. Am. 2020, 147, 3390–3396. [Google Scholar] [CrossRef] [PubMed]
- Pace, D.S.; Arcangeli, A.; Mussi, B.; Vivaldi, C.; Ledon, C.; Lagorio, S.; Giacomini, G.; Pavan, G.; Ardizzone, G. Habitat suitability modeling in different sperm whale social groups. J. Wildl. Manag. 2018, 82, 1062–1073. [Google Scholar] [CrossRef]
- Hartman, K.L.; Fernandez, M.; Azevedo, J.M. Spatial segregation of calving and nursing Risso’s dolphins (Grampus griseus) in the Azores, and its conservation implications. Mar. Biol. 2014, 161, 1419–1428. [Google Scholar] [CrossRef]
- Cafaro, V.; Angeletti, D.; Bellisario, B.; Macali, A.; Carere, C.; Alessi, J. Habitat overlap between bottlenose dolphins and seabirds: A pilot study to identify high-presence coastal areas in the Tyrrhenian Sea. J. Mar. Biol. Assoc. UK 2016, 96, 891–901. [Google Scholar] [CrossRef]
- Crosti, R.; Arcangeli, A.; Campana, I.; Paraboschi, M.; González-Fernández, D. ‘Down to the river’: Amount, composition, and economic sector of litter entering the marine compartment, through the Tiber river in the Western Mediterranean Sea. Rend. Lincei Sci. Fis. Nat. 2018, 29, 859–866. [Google Scholar] [CrossRef]
- Inghilesi, R.; Ottolenghi, L.; Orasi, A.; Pizzi, C.; Bignami, F.; Santoleri, R. Fate of river Tiber discharge investigated through numerical simulation and satellite monitoring. Ocean Sci. 2012, 8, 773–786. [Google Scholar] [CrossRef]
- Nykänen, M.; Dillane, E.; Englund, A.; Foote, A.D.; Ingram, S.N.; Louis, M.; Mirimin, L.; Oudejans, M.; Rogan, E. Quantifying dispersal between marine protected areas by a highly mobile species, the bottlenose dolphin, Tursiops truncatus. Ecol. Evol. 2018, 8, 9241–9258. [Google Scholar] [CrossRef] [PubMed]
- Mann, J.; Connor, R.C.; Barre, L.M.; Heithaus, M.R. Female reproductive success in bottlenose dolphins (Tursiops sp.): Life history, habitat, provisioning, and group-size effects. Behav. Ecol. 2000, 11, 210–219. [Google Scholar] [CrossRef]
- Vermeulen, E.; Bräger, S. Demographics of the disappearing bottlenose dolphin in Argentina: A common species on its way out? PLoS ONE 2015, 10, e0119182. [Google Scholar] [CrossRef] [PubMed]
- Vella, A.; Murphy, S.; Giménez, J.; de Stephanis, R.; Mussi, B.; Vella, J.G.; Larbi Doukara, K.; Pace, D.S. The conservation of the endangered Mediterranean common dolphin (Delphinus delphis): Current knowledge and research priorities. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 110–136. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cluster | No. of Dolphins | Occ | MR | YR | RR | P | RS |
|---|---|---|---|---|---|---|---|
| Resident | 42 | 0.167 | 0.416 | 0.851 | 0.173 | 0.017 | 0.738 |
| Part-time | 73 | 0.048 | 0.160 | 0.541 | 0.052 | 0.008 | 0.382 |
| Transient | 232 | 0.016 | 0.062 | 0.250 | 0.013 | 0.019 | 0.008 |
| Model | K | QAICc | ΔQAICc | Cluster | ||
|---|---|---|---|---|---|---|
| 1: Φg, pt, βg | 27 | 648.02 | 0 | Resident | 45 (39, 52) | 77 (65, 91) |
| Part-time | 65 (59, 72) | 111 (97, 128) | ||||
| Transient | 202 (173, 235) | 354 (288, 312) | ||||
| Total | 312 (276, 353) | 533 (458, 621) | ||||
| 2: Φg, pl e f f, βg | 9 | 651.88 | 3.86 | Resident | 47 (40, 54) | 80 (67, 95) |
| Part-time | 67 (60, 75) | 114 (100, 132) | ||||
| Transient | 196 (169, 227) | 335 (282, 397) | ||||
| Total | 310 (275, 349) | 529 (456, 614) | ||||
| 3: Φg, pt, βt | 43 | 683.86 | 35.84 | Resident | 38 (31, 46) | 65 (53, 80) |
| Part-time | 58 (51, 66) | 99 (85, 116) | ||||
| Transient | 196 (164, 236) | 336 (274, 412) | ||||
| Total | 292 (252, 339) | 500 (421, 593) | ||||
| 4: Φg, pl e f f, β | 7 | 696.56 | 48.55 | Resident | 47 (42, 52) | 80 (69, 92) |
| Part-time | 67 (62, 73) | 115 (102, 129) | ||||
| Transient | 187 (163, 216) | 320 (271, 379) | ||||
| Total | 301 (271, 335) | 514 (448, 591) |
| Overall | 2017 | 2018 | 2019 | 2020 | |
|---|---|---|---|---|---|
| 0.585 | 0.786 | 0.752 | 0.64 | 0.534 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pace, D.S.; Di Marco, C.; Giacomini, G.; Ferri, S.; Silvestri, M.; Papale, E.; Casoli, E.; Ventura, D.; Mingione, M.; Alaimo Di Loro, P.; et al. Capitoline Dolphins: Residency Patterns and Abundance Estimate of Tursiops truncatus at the Tiber River Estuary (Mediterranean Sea). Biology 2021, 10, 275. https://doi.org/10.3390/biology10040275
Pace DS, Di Marco C, Giacomini G, Ferri S, Silvestri M, Papale E, Casoli E, Ventura D, Mingione M, Alaimo Di Loro P, et al. Capitoline Dolphins: Residency Patterns and Abundance Estimate of Tursiops truncatus at the Tiber River Estuary (Mediterranean Sea). Biology. 2021; 10(4):275. https://doi.org/10.3390/biology10040275
Chicago/Turabian StylePace, Daniela Silvia, Chiara Di Marco, Giancarlo Giacomini, Sara Ferri, Margherita Silvestri, Elena Papale, Edoardo Casoli, Daniele Ventura, Marco Mingione, Pierfrancesco Alaimo Di Loro, and et al. 2021. "Capitoline Dolphins: Residency Patterns and Abundance Estimate of Tursiops truncatus at the Tiber River Estuary (Mediterranean Sea)" Biology 10, no. 4: 275. https://doi.org/10.3390/biology10040275
APA StylePace, D. S., Di Marco, C., Giacomini, G., Ferri, S., Silvestri, M., Papale, E., Casoli, E., Ventura, D., Mingione, M., Alaimo Di Loro, P., Jona Lasinio, G., & Ardizzone, G. (2021). Capitoline Dolphins: Residency Patterns and Abundance Estimate of Tursiops truncatus at the Tiber River Estuary (Mediterranean Sea). Biology, 10(4), 275. https://doi.org/10.3390/biology10040275