
Response of Pluripotent Stem Cells to Environmental Stress and Its Application for Directed Differentiation



Simple Summary
Abstract
1. Introduction
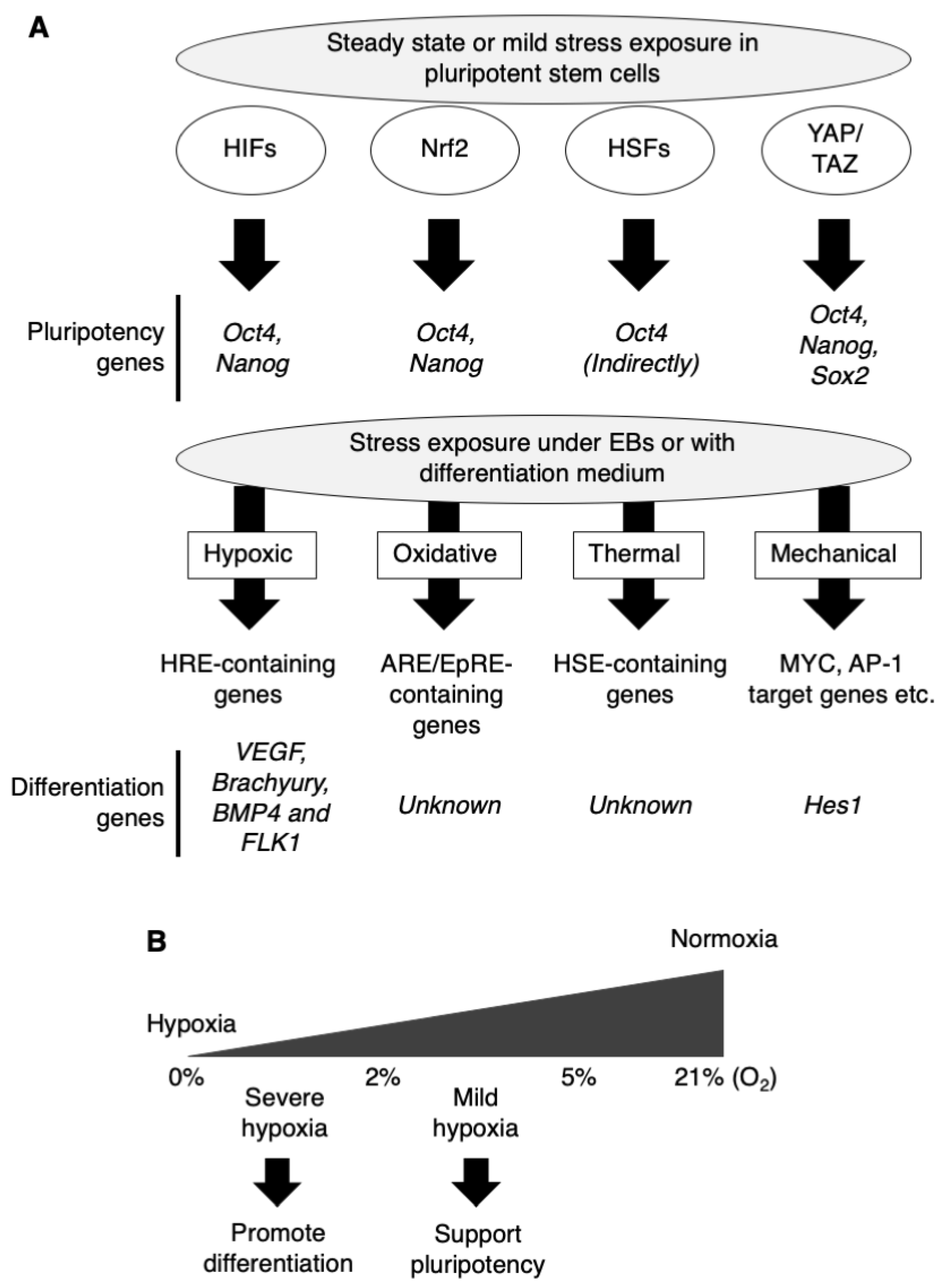
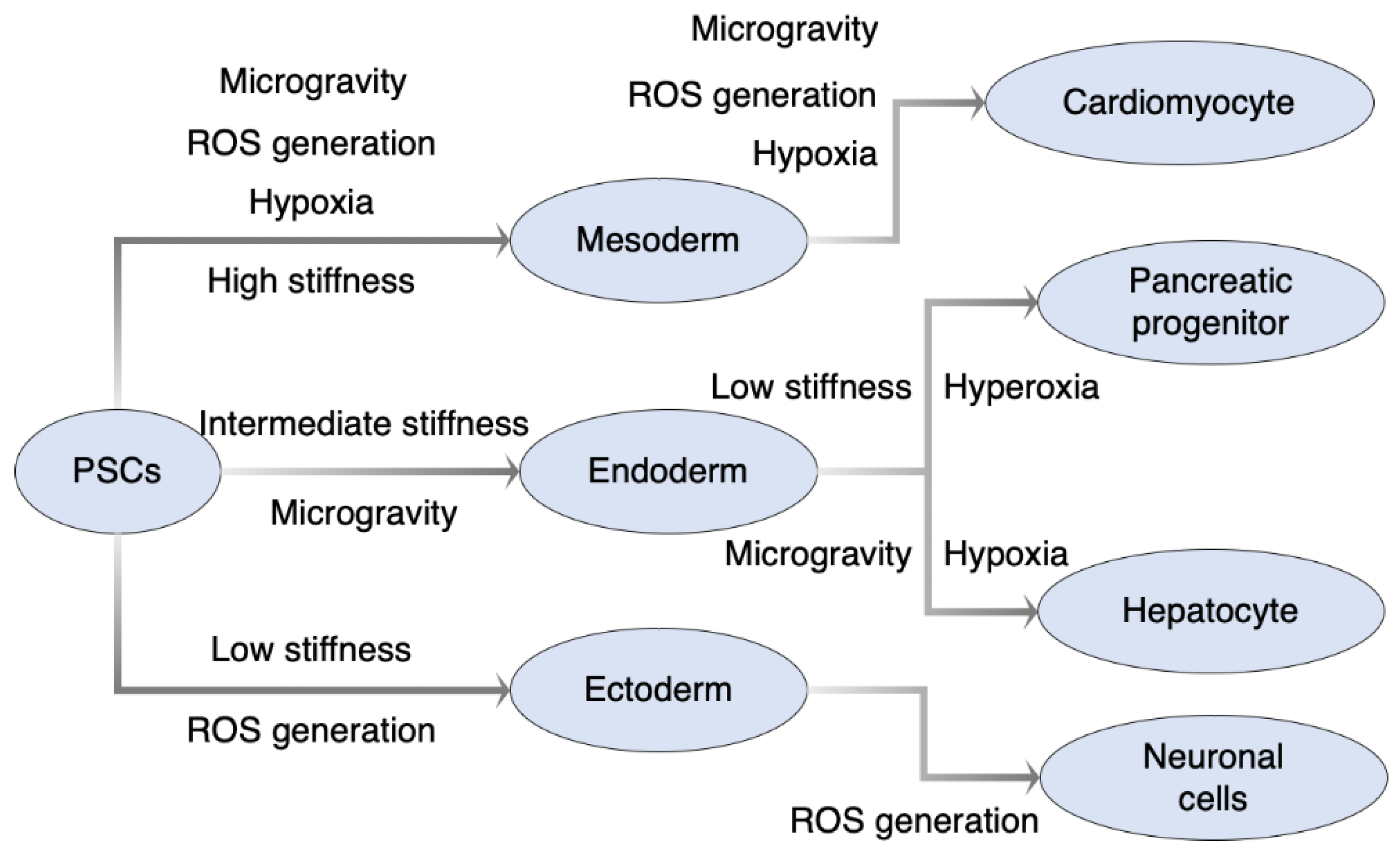
2. Hypoxic Stress for Directed Differentiation
2.1. Involvement of Hypoxia Signaling in Pluripotency of Pluripotent Stem Cells
2.2. Effect of Hypoxic Stress on Directed Differentiation of Pluripotent Stem Cells
3. Oxidative Stress for Directed Differentiation
3.1. Involvement of Oxidative Stress Signaling in Pluripotency of Pluripotent Stem Cells
3.2. Effect of Oxidative Stress on Directed Differentiation of Pluripotent Stem Cells
4. Thermal Stress for Directed Differentiation
4.1. Involvement of Thermal Stress Signaling in Pluripotency of Pluripotent Stem Cells
4.2. Effect of Thermal Stress on Directed Differentiation of Pluripotent Stem Cells
5. Mechanical Stress for Directed Differentiation
5.1. Involvement of Mechanical Stress Signaling in Pluripotency of Pluripotent Stem Cells
5.2. Effect of Mechanical Stress on Directed Differentiation of Pluripotent Stem Cells
6. Physical Stimulation for Directed Differentiation
6.1. Involvement of Physical Stimulation in Pluripotency of Pluripotent Stem Cells
6.2. Effect of Physical Stimuli on Directed Differentiation of Pluripotent Stem Cells
7. Other Candidates for the Method of Directed Differentiation with Environmental Stresses
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic Stem Cell Lines Derived from Human Blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, L.; Ayyash, M.; Novershtern, N.; Hanna, J.H. Dynamic Stem Cell States: Naive to Primed Pluripotency in Rodents and Humans. Nat. Rev. Mol. Cell Biol. 2016, 17, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Gaspar-Maia, A.; Alajem, A.; Meshorer, E.; Ramalho-Santos, M. Open Chromatin in Pluripotency and Reprogramming. Nat. Rev. Mol. Cell Biol. 2011, 12, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Ivan, M.; Kaelin, W.G. The EGLN-HIF O2-Sensing System: Multiple Inputs and Feedbacks. Mol. Cell 2017, 66, 772–779. [Google Scholar] [CrossRef] [PubMed]
- Kaelin, W.G.; Ratcliffe, P.J. Oxygen Sensing by Metazoans: The Central Role of the HIF Hydroxylase Pathway. Mol. Cell 2008, 30, 393–402. [Google Scholar] [CrossRef]
- Semenza, G.L. Hypoxia-Inducible Factors in Physiology and Medicine. Cell 2012, 148, 399–408. [Google Scholar] [CrossRef]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A.-L. The Keap1-Nrf2 Pathway: Mechanisms of Activation and Dysregulation in Cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef]
- Tebay, L.E.; Robertson, H.; Durant, S.T.; Vitale, S.R.; Penning, T.M.; Dinkova-Kostova, A.T.; Hayes, J.D. Mechanisms of Activation of the Transcription Factor Nrf2 by Redox Stressors, Nutrient Cues, and Energy Status and the Pathways through Which It Attenuates Degenerative Disease. Free Radic. Biol. Med. 2015, 88 Pt B, 108–146. [Google Scholar] [CrossRef]
- Gomez-Pastor, R.; Burchfiel, E.T.; Thiele, D.J. Regulation of Heat Shock Transcription Factors and Their Roles in Physiology and Disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 4–19. [Google Scholar] [CrossRef]
- Totaro, A.; Panciera, T.; Piccolo, S. YAP/TAZ Upstream Signals and Downstream Responses. Nat. Cell Biol. 2018, 20, 888–899. [Google Scholar] [CrossRef] [PubMed]
- Panciera, T.; Azzolin, L.; Cordenonsi, M.; Piccolo, S. Mechanobiology of YAP and TAZ in Physiology and Disease. Nat. Rev. Mol. Cell Biol. 2017, 18, 758–770. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, I.; McCollum, D. Control of Cellular Responses to Mechanical Cues through YAP/TAZ Regulation. J. Biol. Chem. 2019, 294, 17693–17706. [Google Scholar] [CrossRef]
- Simon, M.C.; Keith, B. The Role of Oxygen Availability in Embryonic Development and Stem Cell Function. Nat. Rev. Mol. Cell Biol. 2008, 9, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Akerfelt, M.; Morimoto, R.I.; Sistonen, L. Heat Shock Factors: Integrators of Cell Stress, Development and Lifespan. Nat. Rev. Mol. Cell Biol. 2010, 11, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.M.; Maltagliati, A.J. Nrf2 at the Heart of Oxidative Stress and Cardiac Protection. Physiol. Genom. 2018, 50, 77–97. [Google Scholar] [CrossRef]
- Discher, D.E.; Mooney, D.J.; Zandstra, P.W. Growth Factors, Matrices, and Forces Combine and Control Stem Cells. Science 2009, 324, 1673–1677. [Google Scholar] [CrossRef]
- Sridharan, S.; Varghese, R.; Venkatraj, V.; Datta, A. Hypoxia Stress Response Pathways: Modeling and Targeted Therapy. IEEE J. Biomed. Health Inform. 2017, 21, 875–885. [Google Scholar] [CrossRef]
- Dunwoodie, S.L. The Role of Hypoxia in Development of the Mammalian Embryo. Dev. Cell 2009, 17, 755–773. [Google Scholar] [CrossRef]
- Iyer, N.V.; Kotch, L.E.; Agani, F.; Leung, S.W.; Laughner, E.; Wenger, R.H.; Gassmann, M.; Gearhart, J.D.; Lawler, A.M.; Yu, A.Y.; et al. Cellular and Developmental Control of O2 Homeostasis by Hypoxia-Inducible Factor 1 Alpha. Genes Dev. 1998, 12, 149–162. [Google Scholar] [CrossRef]
- Arthur, S.A.; Blaydes, J.P.; Houghton, F.D. Glycolysis Regulates Human Embryonic Stem Cell Self-Renewal under Hypoxia through HIF-2α and the Glycolytic Sensors CTBPs. Stem Cell Rep. 2019, 12, 728–742. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-F.; Kuo, H.-C.; Lin, S.-P.; Chien, C.-L.; Chiang, M.-S.; Ho, H.-N. Hypoxic Culture Maintains Self-Renewal and Enhances Embryoid Body Formation of Human Embryonic Stem Cells. Tissue Eng. Part A 2010, 16, 2901–2913. [Google Scholar] [CrossRef] [PubMed]
- Ezashi, T.; Das, P.; Roberts, R.M. Low O2 Tensions and the Prevention of Differentiation of HES Cells. Proc. Natl. Acad. Sci. USA 2005, 102, 4783–4788. [Google Scholar] [CrossRef] [PubMed]
- Forristal, C.E.; Wright, K.L.; Hanley, N.A.; Oreffo, R.O.C.; Houghton, F.D. Hypoxia Inducible Factors Regulate Pluripotency and Proliferation in Human Embryonic Stem Cells Cultured at Reduced Oxygen Tensions. Reproduction 2010, 139, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.M.; Czepiel, M.; Cetinkaya, C.; Smigielska, K.; Weli, S.C.; Lysdahl, H.; Gabrielsen, A.; Petersen, K.; Ehlers, N.; Fink, T.; et al. Continuous Hypoxic Culturing Maintains Activation of Notch and Allows Long-Term Propagation of Human Embryonic Stem Cells without Spontaneous Differentiation. Cell Prolif. 2009, 42, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Westfall, S.D.; Sachdev, S.; Das, P.; Hearne, L.B.; Hannink, M.; Roberts, R.M.; Ezashi, T. Identification of Oxygen-Sensitive Transcriptional Programs in Human Embryonic Stem Cells. Stem Cells Dev. 2008, 17, 869–881. [Google Scholar] [CrossRef]
- Gustafsson, M.V.; Zheng, X.; Pereira, T.; Gradin, K.; Jin, S.; Lundkvist, J.; Ruas, J.L.; Poellinger, L.; Lendahl, U.; Bondesson, M. Hypoxia Requires Notch Signaling to Maintain the Undifferentiated Cell State. Dev. Cell 2005, 9, 617–628. [Google Scholar] [CrossRef]
- Petruzzelli, R.; Christensen, D.R.; Parry, K.L.; Sanchez-Elsner, T.; Houghton, F.D. HIF-2α Regulates NANOG Expression in Human Embryonic Stem Cells Following Hypoxia and Reoxygenation through the Interaction with an Oct-Sox Cis Regulatory Element. PLoS ONE 2014, 9, e108309. [Google Scholar] [CrossRef]
- Covello, K.L. HIF-2 Regulates Oct-4: Effects of Hypoxia on Stem Cell Function, Embryonic Development, and Tumor Growth. Genes Dev. 2006, 20, 557–570. [Google Scholar] [CrossRef]
- Jarecki, J.; Johnson, E.; Krasnow, M.A. Oxygen Regulation of Airway Branching in Drosophila Is Mediated by Branchless FGF. Cell 1999, 99, 211–220. [Google Scholar] [CrossRef]
- Provot, S.; Zinyk, D.; Gunes, Y.; Kathri, R.; Le, Q.; Kronenberg, H.M.; Johnson, R.S.; Longaker, M.T.; Giaccia, A.J.; Schipani, E. Hif-1alpha Regulates Differentiation of Limb Bud Mesenchyme and Joint Development. J. Cell Biol. 2007, 177, 451–464. [Google Scholar] [CrossRef] [PubMed]
- Schipani, E.; Ryan, H.E.; Didrickson, S.; Kobayashi, T.; Knight, M.; Johnson, R.S. Hypoxia in Cartilage: HIF-1alpha Is Essential for Chondrocyte Growth Arrest and Survival. Genes Dev. 2001, 15, 2865–2876. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Hammer, R.E.; Matsumoto, A.M.; Russell, D.W.; McKnight, S.L. The Hypoxia-Responsive Transcription Factor EPAS1 Is Essential for Catecholamine Homeostasis and Protection against Heart Failure during Embryonic Development. Genes Dev. 1998, 12, 3320–3324. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, V.; Choo-Wing, R.; Lee, C.G.; Zhu, Z.; Nedrelow, J.H.; Chupp, G.L.; Zhang, X.; Matthay, M.A.; Ware, L.B.; Homer, R.J.; et al. Hyperoxia Causes Angiopoietin 2-Mediated Acute Lung Injury and Necrotic Cell Death. Nat. Med. 2006, 12, 1286–1293. [Google Scholar] [CrossRef] [PubMed]
- Heinis, M.; Simon, M.-T.; Ilc, K.; Mazure, N.M.; Pouysségur, J.; Scharfmann, R.; Duvillié, B. Oxygen Tension Regulates Pancreatic Beta-Cell Differentiation through Hypoxia-Inducible Factor 1alpha. Diabetes 2010, 59, 662–669. [Google Scholar] [CrossRef]
- Fraker, C.A.; Alvarez, S.; Papadopoulos, P.; Giraldo, J.; Gu, W.; Ricordi, C.; Inverardi, L.; Domínguez-Bendala, J. Enhanced Oxygenation Promotes Beta-Cell Differentiation in Vitro. Stem Cells 2007, 25, 3155–3164. [Google Scholar] [CrossRef]
- Hakim, F.; Kaitsuka, T.; Raeed, J.M.; Wei, F.-Y.; Shiraki, N.; Akagi, T.; Yokota, T.; Kume, S.; Tomizawa, K. High Oxygen Condition Facilitates the Differentiation of Mouse and Human Pluripotent Stem Cells into Pancreatic Progenitors and Insulin-Producing Cells. J. Biol. Chem. 2014, 289, 9623–9638. [Google Scholar] [CrossRef]
- Lee, J.C.; Smith, S.B.; Watada, H.; Lin, J.; Scheel, D.; Wang, J.; Mirmira, R.G.; German, M.S. Regulation of the Pancreatic Pro-Endocrine Gene Neurogenin3. Diabetes 2001, 50, 928–936. [Google Scholar] [CrossRef]
- Lee, S.; Jeong, H.; Lee, J.; Yang, J.; Lee, E.J.; Kim, S.; Youn, S.; Lee, J.; Kim, W.J.; Kim, K.; et al. Hypoxic Priming of MESCs Accelerates Vascular-lineage Differentiation through HIF1-mediated Inverse Regulation of Oct4 and VEGF. EMBO Mol. Med. 2012, 4, 924–938. [Google Scholar] [CrossRef]
- Bianco, C.; Cotten, C.; Lonardo, E.; Strizzi, L.; Baraty, C.; Mancino, M.; Gonzales, M.; Watanabe, K.; Nagaoka, T.; Berry, C.; et al. Cripto-1 Is Required for Hypoxia to Induce Cardiac Differentiation of Mouse Embryonic Stem Cells. Am. J. Pathol. 2009, 175, 2146–2158. [Google Scholar] [CrossRef]
- Niebruegge, S.; Bauwens, C.L.; Peerani, R.; Thavandiran, N.; Masse, S.; Sevaptisidis, E.; Nanthakumar, K.; Woodhouse, K.; Husain, M.; Kumacheva, E.; et al. Generation of Human Embryonic Stem Cell-Derived Mesoderm and Cardiac Cells Using Size-Specified Aggregates in an Oxygen-Controlled Bioreactor. Biotechnol. Bioeng. 2009, 102, 493–507. [Google Scholar] [CrossRef]
- Koay, E.J.; Athanasiou, K.A. Hypoxic Chondrogenic Differentiation of Human Embryonic Stem Cells Enhances Cartilage Protein Synthesis and Biomechanical Functionality. Osteoarthr. Cartil. 2008, 16, 1450–1456. [Google Scholar] [CrossRef] [PubMed]
- Tsang, K.M.; Hyun, J.S.; Cheng, K.T.; Vargas, M.; Mehta, D.; Ushio-Fukai, M.; Zou, L.; Pajcini, K.V.; Rehman, J.; Malik, A.B. Embryonic Stem Cell Differentiation to Functional Arterial Endothelial Cells through Sequential Activation of ETV2 and NOTCH1 Signaling by HIF1α. Stem Cell Rep. 2017, 9, 796–806. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Bergeron, D.L.; Runge, A.; Dahl, K.D.C.; Fehling, H.J.; Keller, G.; Simon, M.C. Hypoxia Affects Mesoderm and Enhances Hemangioblast Specification during Early Development. Development 2004, 131, 4623–4634. [Google Scholar] [CrossRef] [PubMed]
- Ayabe, H.; Anada, T.; Kamoya, T.; Sato, T.; Kimura, M.; Yoshizawa, E.; Kikuchi, S.; Ueno, Y.; Sekine, K.; Camp, J.G.; et al. Optimal Hypoxia Regulates Human IPSC-Derived Liver Bud Differentiation through Intercellular TGFB Signaling. Stem Cell Rep. 2018, 11, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, A.; Raynaud, P.; Cordi, S.; Zong, Y.; Tronche, F.; Stanger, B.Z.; Jacquemin, P.; Pierreux, C.E.; Clotman, F.; Lemaigre, F.P. Intrahepatic Bile Ducts Develop According to a New Mode of Tubulogenesis Regulated by the Transcription Factor SOX9. Gastroenterology 2009, 136, 2325–2333. [Google Scholar] [CrossRef]
- Hu, Q.; Khanna, P.; Ee Wong, B.S.; Lin Heng, Z.S.; Subhramanyam, C.S.; Thanga, L.Z.; Sing Tan, S.W.; Baeg, G.H. Oxidative Stress Promotes Exit from the Stem Cell State and Spontaneous Neuronal Differentiation. Oncotarget 2018, 9, 4223–4238. [Google Scholar] [CrossRef]
- Ji, A.-R.; Ku, S.-Y.; Cho, M.S.; Kim, Y.Y.; Kim, Y.J.; Oh, S.K.; Kim, S.H.; Moon, S.Y.; Choi, Y.M. Reactive Oxygen Species Enhance Differentiation of Human Embryonic Stem Cells into Mesendodermal Lineage. Exp. Mol. Med. 2010, 42, 175–186. [Google Scholar] [CrossRef]
- Ding, L.; Liang, X.-G.; Hu, Y.; Zhu, D.-Y.; Lou, Y.-J. Involvement of P38MAPK and Reactive Oxygen Species in Icariin-Induced Cardiomyocyte Differentiation of Murine Embryonic Stem Cells in Vitro. Stem Cells Dev. 2008, 17, 751–760. [Google Scholar] [CrossRef]
- Li, J.; Stouffs, M.; Serrander, L.; Banfi, B.; Bettiol, E.; Charnay, Y.; Steger, K.; Krause, K.-H.; Jaconi, M.E. The NADPH Oxidase NOX4 Drives Cardiac Differentiation: Role in Regulating Cardiac Transcription Factors and MAP Kinase Activation. Mol. Biol. Cell 2006, 17, 3978–3988. [Google Scholar] [CrossRef]
- Jang, J.; Wang, Y.; Lalli, M.A.; Guzman, E.; Godshalk, S.E.; Zhou, H.; Kosik, K.S. Primary Cilium-Autophagy-Nrf2 (PAN) Axis Activation Commits Human Embryonic Stem Cells to a Neuroectoderm Fate. Cell 2016, 165, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Koga, T.; Shiraki, N.; Yano, S.; Suico, M.A.; Morino-Koga, S.; Sato, T.; Shuto, T.; Kume, S.; Kai, H. Mild Electrical Stimulation with Heat Shock Guides Differentiation of Embryonic Stem Cells into Pdx1-Expressing Cells within the Definitive Endoderm. BMC Biotechnol. 2017, 17, 14. [Google Scholar] [CrossRef]
- Yamamoto, K.; Sokabe, T.; Watabe, T.; Miyazono, K.; Yamashita, J.K.; Obi, S.; Ohura, N.; Matsushita, A.; Kamiya, A.; Ando, J. Fluid Shear Stress Induces Differentiation of Flk-1-Positive Embryonic Stem Cells into Vascular Endothelial Cells in Vitro. Am. J. Physiol. Heart Circ. Physiol. 2005, 288, H1915–H1924. [Google Scholar] [CrossRef]
- Wolfe, R.P.; Ahsan, T. Shear Stress during Early Embryonic Stem Cell Differentiation Promotes Hematopoietic and Endothelial Phenotypes. Biotechnol. Bioeng. 2013, 110, 1231–1242. [Google Scholar] [CrossRef] [PubMed]
- Adamo, L.; Naveiras, O.; Wenzel, P.L.; McKinney-Freeman, S.; Mack, P.J.; Gracia-Sancho, J.; Suchy-Dicey, A.; Yoshimoto, M.; Lensch, M.W.; Yoder, M.C.; et al. Biomechanical Forces Promote Embryonic Haematopoiesis. Nature 2009, 459, 1131–1135. [Google Scholar] [CrossRef] [PubMed]
- Richardson, T.; Barner, S.; Candiello, J.; Kumta, P.N.; Banerjee, I. Capsule Stiffness Regulates the Efficiency of Pancreatic Differentiation of Human Embryonic Stem Cells. Acta Biomater. 2016, 35, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Eroshenko, N.; Ramachandran, R.; Yadavalli, V.K.; Rao, R.R. Effect of Substrate Stiffness on Early Human Embryonic Stem Cell Differentiation. J. Biol. Eng. 2013, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Zoldan, J.; Karagiannis, E.D.; Lee, C.Y.; Anderson, D.G.; Langer, R.; Levenberg, S. The Influence of Scaffold Elasticity on Germ Layer Specification of Human Embryonic Stem Cells. Biomaterials 2011, 32, 9612–9621. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.L.; Najia, M.A.; Saeed, R.; McDevitt, T.C. Alginate Encapsulation Parameters Influence the Differentiation of Microencapsulated Embryonic Stem Cell Aggregates: Alginate Encapsulation Influences Differentiation of ESC Aggregates. Biotechnol. Bioeng. 2014, 111, 618–631. [Google Scholar] [CrossRef] [PubMed]
- Mamidi, A.; Prawiro, C.; Seymour, P.A.; de Lichtenberg, K.H.; Jackson, A.; Serup, P.; Semb, H. Mechanosignalling via Integrins Directs Fate Decisions of Pancreatic Progenitors. Nature 2018, 564, 114–118. [Google Scholar] [CrossRef] [PubMed]
- da Costa, R.M.; Rodrigues, D.; Pereira, C.A.; Silva, J.F.; Alves, J.V.; Lobato, N.S.; Tostes, R.C. Nrf2 as a Potential Mediator of Cardiovascular Risk in Metabolic Diseases. Front. Pharmacol. 2019, 10, 382. [Google Scholar] [CrossRef] [PubMed]
- Dröge, W. Free Radicals in the Physiological Control of Cell Function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef] [PubMed]
- Sart, S.; Song, L.; Li, Y. Controlling Redox Status for Stem Cell Survival, Expansion, and Differentiation. Oxidative Med. Cell. Longev. 2015, 2015, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Li, T.-S.; Marbán, E. Physiological Levels of Reactive Oxygen Species Are Required to Maintain Genomic Stability in Stem Cells. Stem Cells 2010, 28, 1178–1185. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.-L.; Chakraborty, S.; Rajan, S.S.; Wang, R.; Huang, F. Effects of Oxidative Stress on Mouse Embryonic Stem Cell Proliferation, Apoptosis, Senescence, and Self-Renewal. Stem Cells Dev. 2010, 19, 1321–1331. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Wang, Y.; Kim, H.-S.; Lalli, M.A.; Kosik, K.S. Nrf2, a Regulator of the Proteasome, Controls Self-Renewal and Pluripotency in Human Embryonic Stem Cells. Stem Cells 2014, 32, 2616–2625. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.E.; Hipp, M.S.; Bracher, A.; Hayer-Hartl, M.; Hartl, F.U. Molecular Chaperone Functions in Protein Folding and Proteostasis. Annu. Rev. Biochem. 2013, 82, 323–355. [Google Scholar] [CrossRef]
- Li, J.; Labbadia, J.; Morimoto, R.I. Rethinking HSF1 in Stress, Development, and Organismal Health. Trends Cell Biol. 2017, 27, 895–905. [Google Scholar] [CrossRef]
- Wu, C. Heat Shock Transcription Factors: Structure and Regulation. Annu. Rev. Cell Dev. Biol. 1995, 11, 441–469. [Google Scholar] [CrossRef]
- Anckar, J.; Sistonen, L. Regulation of H SF 1 Function in the Heat Stress Response: Implications in Aging and Disease. Annu. Rev. Biochem. 2011, 80, 1089–1115. [Google Scholar] [CrossRef]
- Zou, J.; Guo, Y.; Guettouche, T.; Smith, D.F.; Voellmy, R. Repression of Heat Shock Transcription Factor HSF1 Activation by HSP90 (HSP90 Complex) That Forms a Stress-Sensitive Complex with HSF1. Cell 1998, 94, 471–480. [Google Scholar] [CrossRef]
- Guo, Y.; Guettouche, T.; Fenna, M.; Boellmann, F.; Pratt, W.B.; Toft, D.O.; Smith, D.F.; Voellmy, R. Evidence for a Mechanism of Repression of Heat Shock Factor 1 Transcriptional Activity by a Multichaperone Complex. J. Biol. Chem. 2001, 276, 45791–45799. [Google Scholar] [CrossRef] [PubMed]
- Neef, D.W.; Jaeger, A.M.; Gomez-Pastor, R.; Willmund, F.; Frydman, J.; Thiele, D.J. A Direct Regulatory Interaction between Chaperonin TRiC and Stress-Responsive Transcription Factor HSF1. Cell Rep. 2014, 9, 955–966. [Google Scholar] [CrossRef] [PubMed]
- Labbadia, J.; Morimoto, R.I. The Biology of Proteostasis in Aging and Disease. Annu. Rev. Biochem. 2015, 84, 435–464. [Google Scholar] [CrossRef]
- Prinsloo, E.; Setati, M.M.; Longshaw, V.M.; Blatch, G.L. Chaperoning Stem Cells: A Role for Heat Shock Proteins in the Modulation of Stem Cell Self-Renewal and Differentiation? Bioessays 2009, 31, 370–377. [Google Scholar] [CrossRef]
- You, K.T.; Park, J.; Kim, V.N. Role of the Small Subunit Processome in the Maintenance of Pluripotent Stem Cells. Genes Dev. 2015, 29, 2004–2009. [Google Scholar] [CrossRef]
- Noormohammadi, A.; Khodakarami, A.; Gutierrez-Garcia, R.; Lee, H.J.; Koyuncu, S.; König, T.; Schindler, C.; Saez, I.; Fatima, A.; Dieterich, C.; et al. Somatic Increase of CCT8 Mimics Proteostasis of Human Pluripotent Stem Cells and Extends C. Elegans Lifespan. Nat. Commun. 2016, 7, 13649. [Google Scholar] [CrossRef]
- Lyu, X.; Rowley, M.J.; Corces, V.G. Architectural Proteins and Pluripotency Factors Cooperate to Orchestrate the Transcriptional Response of HESCs to Temperature Stress. Mol. Cell 2018, 71, 940–955. [Google Scholar] [CrossRef]
- Byun, K.; Kim, T.-K.; Oh, J.; Bayarsaikhan, E.; Kim, D.; Lee, M.Y.; Pack, C.-G.; Hwang, D.; Lee, B. Heat Shock Instructs HESCs to Exit from the Self-Renewal Program through Negative Regulation of OCT4 by SAPK/JNK and HSF1 Pathway. Stem Cell Res. 2013, 11, 1323–1334. [Google Scholar] [CrossRef]
- Weng, S.; Shao, Y.; Chen, W.; Fu, J. Mechanosensitive Subcellular Rheostasis Drives Emergent Single-Cell Mechanical Homeostasis. Nat. Mater. 2016, 15, 961–967. [Google Scholar] [CrossRef]
- Chen, C.S. Mechanotransduction—A Field Pulling Together? J. Cell. Sci. 2008, 121 Pt 20, 3285–3292. [Google Scholar] [CrossRef]
- Vogel, V.; Sheetz, M. Local Force and Geometry Sensing Regulate Cell Functions. Nat. Rev. Mol. Cell Biol. 2006, 7, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Pathak, M.M.; Nourse, J.L.; Tran, T.; Hwe, J.; Arulmoli, J.; Le, D.T.T.; Bernardis, E.; Flanagan, L.A.; Tombola, F. Stretch-Activated Ion Channel Piezo1 Directs Lineage Choice in Human Neural Stem Cells. Proc. Natl. Acad. Sci. USA 2014, 111, 16148–16153. [Google Scholar] [CrossRef] [PubMed]
- Hoey, D.A.; Tormey, S.; Ramcharan, S.; O’Brien, F.J.; Jacobs, C.R. Primary Cilia-Mediated Mechanotransduction in Human Mesenchymal Stem Cells. Stem Cells 2012, 30, 2561–2570. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.C.; Hoey, D.A.; Chua, M.; Bellon, R.; Jacobs, C.R. Mechanical Signals Promote Osteogenic Fate through a Primary Cilia-Mediated Mechanism. FASEB J. 2016, 30, 1504–1511. [Google Scholar] [CrossRef] [PubMed]
- Dupont, S.; Morsut, L.; Aragona, M.; Enzo, E.; Giulitti, S.; Cordenonsi, M.; Zanconato, F.; Le Digabel, J.; Forcato, M.; Bicciato, S.; et al. Role of YAP/TAZ in Mechanotransduction. Nature 2011, 474, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Lian, I.; Kim, J.; Okazawa, H.; Zhao, J.; Zhao, B.; Yu, J.; Chinnaiyan, A.; Israel, M.A.; Goldstein, L.S.B.; Abujarour, R.; et al. The Role of YAP Transcription Coactivator in Regulating Stem Cell Self-Renewal and Differentiation. Genes Dev. 2010, 24, 1106–1118. [Google Scholar] [CrossRef]
- Souilhol, C.; Serbanovic-Canic, J.; Fragiadaki, M.; Chico, T.J.; Ridger, V.; Roddie, H.; Evans, P.C. Endothelial Responses to Shear Stress in Atherosclerosis: A Novel Role for Developmental Genes. Nat. Rev. Cardiol. 2020, 17, 52–63. [Google Scholar] [CrossRef]
- Vining, K.H.; Mooney, D.J. Mechanical Forces Direct Stem Cell Behaviour in Development and Regeneration. Nat. Rev. Mol. Cell Biol. 2017, 18, 728–742. [Google Scholar] [CrossRef]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix Elasticity Directs Stem Cell Lineage Specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, C.S.; Fu, J. Forcing Stem Cells to Behave: A Biophysical Perspective of the Cellular Microenvironment. Annu. Rev. Biophys. 2012, 41, 519–542. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, F.; Na, S.; Li, D.; Poh, Y.-C.; Tanaka, T.S.; Wang, F.; Wang, N. Material Properties of the Cell Dictate Stress-Induced Spreading and Differentiation in Embryonic Stem Cells. Nat. Mater. 2010, 9, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Wehland, M.; Corydon, T.J.; Richter, P.; Prasad, B.; Bauer, J.; Egli, M.; Kopp, S.; Lebert, M.; Krüger, M. The Effects of Microgravity on Differentiation and Cell Growth in Stem Cells and Cancer Stem Cells. Stem Cells Transl. Med. 2020, 9, 882–894. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Dong, X.; Zhang, F.; Zhu, H.; Hao, T.; Jiang, X.; Zheng, W.; Zhang, T.; Wang, P.; Li, H.; et al. Real Microgravity Condition Promoted Regeneration Capacity of Induced Pluripotent Stem Cells during the TZ-1 Space Mission. Cell Prolif. 2019, 52, e12574. [Google Scholar] [CrossRef] [PubMed]
- Blaber, E.A.; Finkelstein, H.; Dvorochkin, N.; Sato, K.Y.; Yousuf, R.; Burns, B.P.; Globus, R.K.; Almeida, E.A.C. Microgravity Reduces the Differentiation and Regenerative Potential of Embryonic Stem Cells. Stem Cells Dev. 2015, 24, 2605–2621. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Deng, Z.; Zhang, H.; Zhao, H.; Zhou, J.; Liu, S.; Chen, Q.; Ning, L.; Cao, Y.; Wang, X.; et al. Rotary Suspension Culture Enhances Mesendoderm Differentiation of Embryonic Stem Cells through Modulation of Wnt/β-Catenin Pathway. Stem Cell Rev. Rep. 2014, 10, 526–538. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Deng, Z.; Duan, E. Uniform Embryoid Body Production and Enhanced Mesendoderm Differentiation with Murine Embryonic Stem Cells in a Rotary Suspension Bioreactor. Methods Mol. Biol. 2016, 1502, 63–75. [Google Scholar] [CrossRef]
- Li, H.; Zhu, H.; Zhang, F.; Dong, X.; Hao, T.; Jiang, X.; Zheng, W.; Zhang, T.; Chen, X.; Wang, P.; et al. Spaceflight Promoted Myocardial Differentiation of Induced Pluripotent Stem Cells: Results from Tianzhou-1 Space Mission. Stem Cells Dev. 2019, 28, 357–360. [Google Scholar] [CrossRef]
- Jha, R.; Wu, Q.; Singh, M.; Preininger, M.K.; Han, P.; Ding, G.; Cho, H.C.; Jo, H.; Maher, K.O.; Wagner, M.B.; et al. Simulated Microgravity and 3D Culture Enhance Induction, Viability, Proliferation and Differentiation of Cardiac Progenitors from Human Pluripotent Stem Cells. Sci. Rep. 2016, 6, 30956. [Google Scholar] [CrossRef]
- Oss-Ronen, L.; Redden, R.A.; Lelkes, P.I. Enhanced Induction of Definitive Endoderm Differentiation of Mouse Embryonic Stem Cells in Simulated Microgravity. Stem Cells Dev. 2020, 29, 1275–1284. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Zhang, S.; Peng, G.; Liu, T.; Li, Y.; Xiang, D.; Wassler, M.J.; Shelat, H.S.; Geng, Y. Rotating Microgravity-Bioreactor Cultivation Enhances the Hepatic Differentiation of Mouse Embryonic Stem Cells on Biodegradable Polymer Scaffolds. Tissue Eng. Part A 2012, 18, 2376–2385. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.; Choi, H.; Park, H.; Cho, B.; Kim, S.; Kim, J. Effects of a Hypomagnetic Field on DNA Methylation during the Differentiation of Embryonic Stem Cells. Sci. Rep. 2019, 9, 1333. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Chen, C.; Deng, P.; Zhu, G.; Lin, M.; Zhang, L.; Xu, S.; He, M.; Lu, Y.; Duan, W.; et al. Extremely Low-Frequency Electromagnetic Fields Promote In Vitro Neuronal Differentiation and Neurite Outgrowth of Embryonic Neural Stem Cells via Up-Regulating TRPC1. PLoS ONE 2016, 11, e0150923. [Google Scholar] [CrossRef]
- Huang, J.; Wang, D.; Chen, J.; Liu, W.; Duan, L.; You, W.; Zhu, W.; Xiong, J.; Wang, D. Osteogenic Differentiation of Bone Marrow Mesenchymal Stem Cells by Magnetic Nanoparticle Composite Scaffolds under a Pulsed Electromagnetic Field. Saudi Pharm. J. 2017, 25, 575–579. [Google Scholar] [CrossRef]
- Sauer, H.; Rahimi, G.; Hescheler, J.; Wartenberg, M. Effects of Electrical Fields on Cardiomyocyte Differentiation of Embryonic Stem Cells. J. Cell. Biochem. 1999, 75, 710–723. [Google Scholar] [CrossRef]
- Bekhite, M.M.; Finkensieper, A.; Abou–Zaid, F.A.; El-Shourbagy, I.K.; Omar, K.M.; Figulla, H.-R.; Sauer, H.; Wartenberg, M. Static Electromagnetic Fields Induce Vasculogenesis and Chondro-Osteogenesis of Mouse Embryonic Stem Cells by Reactive Oxygen Species-Mediated Up-Regulation of Vascular Endothelial Growth Factor. Stem Cells Dev. 2010, 19, 731–743. [Google Scholar] [CrossRef]
- Sasai, Y. Next-Generation Regenerative Medicine: Organogenesis from Stem Cells in 3D Culture. Cell Stem Cell 2013, 12, 520–530. [Google Scholar] [CrossRef]
- Takebe, T.; Sekine, K.; Enomura, M.; Koike, H.; Kimura, M.; Ogaeri, T.; Zhang, R.-R.; Ueno, Y.; Zheng, Y.-W.; Koike, N.; et al. Vascularized and Functional Human Liver from an IPSC-Derived Organ Bud Transplant. Nature 2013, 499, 481–484. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, H.; Ko, U.H.; Oh, Y.; Lim, A.; Sohn, J.-W.; Shin, J.H.; Kim, H.; Han, Y.-M. Islet-like Organoids Derived from Human Pluripotent Stem Cells Efficiently Function in the Glucose Responsiveness in Vitro and in Vivo. Sci. Rep. 2016, 6, 35145. [Google Scholar] [CrossRef]
- Liu, H.; Ren, C.; Zhu, B.; Wang, L.; Liu, W.; Shi, J.; Lin, J.; Xia, X.; Zeng, F.; Chen, J.; et al. High-Efficient Transfection of Human Embryonic Stem Cells by Single-Cell Plating and Starvation. Stem Cells Dev. 2016, 25, 477–491. [Google Scholar] [CrossRef]
- Charruyer, A.; Ghadially, R. Influence of PH on Skin Stem Cells and Their Differentiation. In Current Problems in Dermatology; Surber, C., Abels, C., Maibach, H., Eds.; S. Karger AG: Basel, Switzerland, 2018; Volume 54, pp. 71–78. [Google Scholar] [CrossRef]
- Li, X.; Karki, P.; Lei, L.; Wang, H.; Fliegel, L. Na+/H+ Exchanger Isoform 1 Facilitates Cardiomyocyte Embryonic Stem Cell Differentiation. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H159–H170. [Google Scholar] [CrossRef] [PubMed]
- Ulmschneider, B.; Grillo-Hill, B.K.; Benitez, M.; Azimova, D.R.; Barber, D.L.; Nystul, T.G. Increased Intracellular PH Is Necessary for Adult Epithelial and Embryonic Stem Cell Differentiation. J. Cell Biol. 2016, 215, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Minami, N.; Yamada, M.; Imai, H. Immobilized PH in Culture Reveals an Optimal Condition for Somatic Cell Reprogramming and Differentiation of Pluripotent Stem Cells. Reprod. Med. Biol. 2017, 16, 58–66. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Stress | Cell Type | Directed Cell Type | Mechanism | References |
|---|---|---|---|---|
| Hypoxic stress | ||||
| Hyperoxia (60% O2) | Mouse ESCs 1, human iPSCs 2 | Pancreatic beta cell | Inhibition of HIF-1αfollowed by Hes1 repression | [37] |
| Hypoxia (1% O2) | Mouse ESCs | Vascular lineage | HIF-1-mediated inverse regulation of Oct4 (down) and VEGF (up) | [39] |
| Hypoxia (0.5% O2) | Mouse ESCs | Mesoderm and cardiomyocyte | HIF-1α mediated Cripto-1 expression | [40] |
| Hypoxia (2% O2) | Human ESCs | Chondrocyte | Undescribed | [42] |
| Hypoxia (1% O2) | Mouse ESCs | Arterial endothelial cells | Activation of ETV2 and NOTCH1 signaling by HIF-1α | [43] |
| Hypoxia (4% O2) | Human ESCs | Cardiomyocytes | Undescribed | [41] |
| Hypoxia (3% O2) | Mouse ESCs | Mesoderm and hemangioblast | Accelerated expression of Brachyury, BMP4 and FLK1 via Arnt | [44] |
| Mild hypoxia (10% O2) | Human iPSCs | Hepatocyte | TGFB signal inhibition | [45] |
| Oxidative stress | ||||
| Paraquat (25 µM) | Human ESCs | Neuronal cells | ROS 3 and activation of MAPK-ERK1/2 | [47] |
| Buthionine sulfoximine (0.2 mM) | Human ESCs | Mesodermal and endodermal lineages | Inactivation of p38 and AKT as well as concomitant transient increase in JNK and ERK signaling | [48] |
| Icariin | Mouse ESCs | Cardiomyocyte | ROS generation and the subsequent activation of p38 MAPK | [49] |
| H2O2 (1~100 nM) | Mouse ESCs | Cardiomyocyte | p38 activation and MEF2C nuclear translocation | [50] |
| Nrf2 shRNA | Human iPSCs | Neuroectoderm | Suppression of Nrf2 binding to pluripotency genes OCT4 and NANOG | [51] |
| Thermal stress | ||||
| Heat shock with mild electrical stimulation (42 °C, 55 pps) | Mouse ESCs | Pdx1-expressing pancreatic progenitors from definitive endoderm | Upregulation of Hsp72 and activation of Akt, ERK, p38 and JNK (putative). | [52] |
| Mechanical stress | ||||
| Fluid shear stress | Mouse ESCs | Vascular endothelial cell | Flk-1 activation and VEGF production | [53] |
| Fluid shear stress | Mouse ESCs | Endothelial and hematopoietic cell | Flk1 activation | [54] |
| Fluid shear stress | Mouse ESCs | Hematopoietic cell | Increased Runx1 expression | [55] |
| High stiffness (BAlg 4 capsule, ~22 kPa) | Human ESCs | Definitive endoderm | Increase in pSMAD/pAkt | [56] |
| Low stiffness (BAlg capsule, ~4 kPa) | Human ESCs | Pancreatic progenitor | Decrease in SHH signaling | [56] |
| High stiffness | Human ESCs | Mesoderm | Undescribed | [57] |
| High stiffness (3D scaffold, 1.5–6 MPa) | Human ESCs | Mesoderm | Undescribed (similar elasticity during gastrulation could be related) | [58] |
| Intermediate stiffness (3D scaffold, 0.1–1 MPa) | Human ESCs | Endoderm | Undescribed (similar elasticity during gastrulation could be related) | [58] |
| Low stiffness (3D scaffold, <0.1 MPa) | Human ESCs | Ectoderm | Undescribed (similar elasticity during gastrulation could be related) | [58] |
| Low stiffness (encapsulated by alginate microbeads) | Mouse ESCs | Endoderm | Undescribed | [59] |
| Confinement (~300 µm2) | Human ESCs | Pancreatic endocrine progenitor | Inhibition of YAP1 | [60] |
| Stress | Cell Type | Directed Cell Type | Mechanism | References |
|---|---|---|---|---|
| Microgravity | ||||
| Rotary suspension culture | Mouse ESCs 1 | Mesoderm | Enhancement of Wnt/β-catenin signaling | [96,97] |
| Spaceflight | Mouse iPSCs 2 | Cardiomyocyte | Undescribed | [98] |
| Simulated microgravity and 3D culture | Human ESCs and iPSCs | Cardiomyocyte | Increased proliferation and viability of cardiac progenitors via up-regulation of heat shock proteins and BIRC5 | [99] |
| Simulated microgravity in rotary bioreactor | Mouse ESCs (embryoid body) | Definitive endoderm | Undescribed | [100] |
| Simulated microgravity in rotary bioreactor | Mouse ESCs | Hepatocyte | Undescribed | [101] |
| EMF3 | ||||
| Single electrical field (500 V/m) | Mouse ESCs | Cardiomyocyte | Intracellular ROS 4 generation and NF-κB 5 activation | [105] |
| Static EMF (0.4–2 mT) | Mouse ESCs | Vasculogenesis and chondro-osteogenesis | Intracellular ROS generation and VEGF 6 induction | [106] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaitsuka, T.; Hakim, F. Response of Pluripotent Stem Cells to Environmental Stress and Its Application for Directed Differentiation. Biology 2021, 10, 84. https://doi.org/10.3390/biology10020084
Kaitsuka T, Hakim F. Response of Pluripotent Stem Cells to Environmental Stress and Its Application for Directed Differentiation. Biology. 2021; 10(2):84. https://doi.org/10.3390/biology10020084
Chicago/Turabian StyleKaitsuka, Taku, and Farzana Hakim. 2021. "Response of Pluripotent Stem Cells to Environmental Stress and Its Application for Directed Differentiation" Biology 10, no. 2: 84. https://doi.org/10.3390/biology10020084
APA StyleKaitsuka, T., & Hakim, F. (2021). Response of Pluripotent Stem Cells to Environmental Stress and Its Application for Directed Differentiation. Biology, 10(2), 84. https://doi.org/10.3390/biology10020084