Simple Summary
Plants are sessile organisms that are continuously exposed to adverse environmental factors, both abiotic and biotic. Plant immunity is an intricate system that involves a remarkable array of structural, chemical, and protein-based layers of defense, aiming to stop pathogens before they cause irreversible damages. Proteases are an integral part of plant defense systems, with several hubs of action, from pathogen recognition and priming to the activation of plant hypersensitive response. Within this wide group of proteolytic enzymes, aspartic proteases have been implicated in several plant development functions and are gaining more prominence due to their involvement in plant–pathogen interactions. In this review, we summarize the current knowledge on plant and pathogenic aspartic proteases and highlight the most recent findings on their participation on plant defense, as well as in pathogen infection strategies.
Abstract
Plant aspartic proteases (APs; E.C.3.4.23) are a group of proteolytic enzymes widely distributed among different species characterized by the conserved sequence Asp-Gly-Thr at the active site. With a broad spectrum of biological roles, plant APs are suggested to undergo functional specialization and to be crucial in developmental processes, such as in both biotic and abiotic stress responses. Over the last decade, an increasing number of publications highlighted the APs’ involvement in plant defense responses against a diversity of stresses. In contrast, few studies regarding pathogen-secreted APs and AP inhibitors have been published so far. In this review, we provide a comprehensive picture of aspartic proteases from plant and pathogenic origins, focusing on their relevance and participation in defense and offense strategies in plant–pathogen interactions.
1. The Past and the Present of Aspartic Proteases
Aspartic proteases (APs) were first discovered in animals during the nineteenth century. In 1836, Theodor Schwann described pepsin, which he identified during the study of animal gastric juices [1]. Later, in 1875, a pepsin-like proteinase was described in the pitcher plant (Nepenthes) after treatment of the plants’ digestive juice with sulfuric acid [2]. Almost a century after the discovery of the first AP, in 1930, the purification and crystallization of swine pepsin by John Northrop provided a substantial evidence that proteases were proteins [3]. In the following years, other proteases were crystallized and studied, including chymotrypsin, trypsin, and pepsinogen [4]. The conversion of pepsinogen to the active form of pepsin is an autocatalytic process that occurs at a low pH (1.5–5). Based on these findings, in 1962, the first step was taken towards the study of acidic proteinases [5]. In 1970, the discovery of pepstatin [6], a powerful inhibitor of aspartic proteases, encouraged its use as an immobilized compound for affinity purification of these proteases [7,8]. A major breakthrough occurred in 1972, when the complete amino acid sequence of the pig pepsin was uncovered [9]. Later, in the 1980s, the current terminology of the aspartic (or aspartyl) proteases was established, resulting from the observation that the carboxyl groups belonging to aspartate residues were involved in the catalytic process [10].
Although most studies about APs were performed in mammals, yeast, and fungi, some work has been developed in plants [11]. Plant APs were purified from the seeds of many organisms, such as Oryza sativa [12], Cucurbita maxima [13], Cucumis sativus [13], Triticum aestivum [14], and Hordeum vulgare [15], as well as from Lycopersicon esculentum leaves [16]. In 1991, the DNA sequence of the first plant aspartic protease, from barley (Hordeurn vulgare), was sequenced [17]. In the following years, APs from other organisms, including one from Arabidopsis thaliana, were isolated, providing more information about plant APs’ structure [7].
According to the MEROPS database (http://www.merops.ac.uk), aspartic proteases (EC 3.4.23) are grouped into 16 families, based on similarities of the amino acid sequences of the catalytic site. These families are clustered into five different clans that reflect a common evolutionary origin and similar tertiary structure [18]. Plant aspartic proteases are distributed among 12 of the 16 families: A1, A2, A3, A9, A11, A28, and A32 of clan AA; families A22 and A24 of clan AD; family A8 and A31 of clan AC and AE, respectively; and family A36 which has not yet been assigned to a clan [19]. A majority of plant APs belong to A1 family [20].
In 2004, with the completion of Arabidopsis genome, new perspectives have risen regarding plant APs’ diversity [21]. The first plant aspartic protease gene family to be described belonged to Arabidopsis with 51 known genes [22]. In the following years, plant APs have been found in increasing numbers [23] with 96 OsAP genes in rice (Oryza sativa) [24], 50 VvAP genes in grapevine (Vitis vinifera) [25], and 67 PtAP genes identified in black cottonwood (Populus trichocarpa) [26].
So far, it is known that plant APs are involved in several cell mechanisms, from developmental processes [27,28,29] to abiotic [30,31,32] and biotic stress responses [33,34,35]. Major milestones concerning aspartic proteases’ history and relation to pathogen resistance are presented in Figure 1.
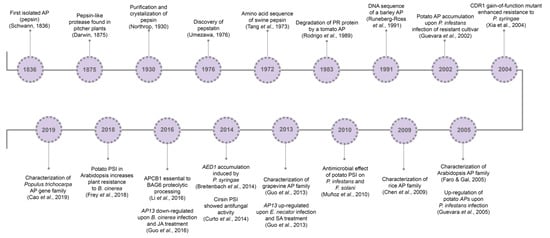
Figure 1.
Aspartic proteases historical breakthroughs. AP, aspartic protease; CDR1, constitutive disease resistance 1; PSI, plant-specific insert; AP13, aspartic protease 13; SA, salicylic acid; AED1, apoplastic enhanced disease susceptibility 1; APCB1, aspartyl protease cleaving bcl-2 associated athanogene; BAG6, BCL-2 associated athanogene protein 6; SA, salicylic acid; JA, jasmonic acid; MeJA, methyl jasmonate.
2. The Features of Plant Aspartic Proteases
2.1. Structure and Classification
In the past years, with the study of phytepsin (AP from barley seeds) and cardosin (AP from the flowers of Cynara cardunculus), relevant information has been generated about plant aspartic proteases [22,36,37]. Plant APs, mostly belonging to family A1, are generally active at acid pH (pH 2–6), are specifically inhibited by pepstatin A, and comprise two aspartic acid residues essential for the catalytic activity [20,38]. The catalytic motifs of plant aspartic proteases from A1 family are usually Asp-Thr-Gly (DTG) or Asp-Ser-Gly (DSG) [20]. Although the general structure of the plant APs has similarities to that of mammals and microorganisms, plant APs contain a plant-specific insert (PSI) in the C-terminal region [7].
Most of the knowledge about A1 family plant APs comes from the study of typical APs, such as phytepsin and cardosin A and B [20]. Typical APs possess a signal peptide, a prosegment, and a PSI, and the catalytic site is composed by hydrophobic-hydrophobic-DTG-Ser-Ser residues (Figure 2). Exceptions to the structure of typical aspartic proteases were already described, as in the case of nucellin [39], in chloroplast nucleoid DNA-binding protein (CND41) [40], and in the constitutive disease resistance 1 (CDR1) protease [33]. These structural exceptions gave rise to three different categories, depending on the putative domain organization and active site sequence motifs: typical, nucellin-like, and atypical aspartic proteases [22]. Atypical and nucellin-like APs have distinct features on primary structure organization that differ from typical APs. The nucellin-like APs lack the prosegment and the PSI and comprise proteins similar to nucellin [20] with a characteristic sequence of residues: acidic-hydrophobic-DTG-serine-acidic residues around the catalytic site [22]. Atypical APs have intermediate features between typical and nucellin-like, and the active site is composed by hydrophobic-hydrophobic-DTG-Ser-acidic residues [20]. Both atypical and nucellin-like APs have a cysteine-rich region designated nepenthesin-type AP (NAP) specific insertion [38].
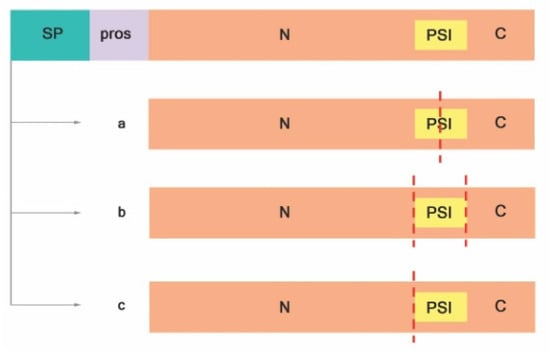
Figure 2.
Proteolytic activation of typical APs adapted from Reference [41]. (a) PSI is digested at the midsection, (b) PSI is entirely removed, and (c) PSI and C-terminal are removed. Signal peptide (SP); prosegment (pros); plant-specific insert (PSI); N-terminal domain (N); C-terminal domain (C); red dashed lines indicate cleavage sites.
Detailed information on structure organization of plant aspartic proteases has been extensively reviewed in Reference [38].
2.2. Activation of Aspartic Proteases and Their Subcellular Localization
Proteolytic cleavage is crucial for active proteases. It starts with the removal of the signal sequence upon translocation to the ER lumen resulting in proproteins (zymogens). Usually, processing zymogens of typical plant APs involves the removal of the prosegment and partial or total deletion of PSI in an autocatalytic manner at the low pH of the vacuole [20,41]. Cheung and colleagues have proposed that, after proteolytic cleavage and activation, typical plant APs are either heterodimeric, where the PSI is partially digested (Figure 2a) [42,43,44] or entirely removed (Figure 2b; Reference [45]), or monomeric, without PSI (Figure 2c), as was observed in sweet potato SPAP1 [46].
There is evidence that two monomeric APs from potato tuber and leaves, StAsp1 [47] and StAsp3 [48], respectively, have the PSI in their mature form. However, the proteolytic mechanisms behind that process are still unknown [49,50]. In contrast, two atypical aspartic proteases, CDR1 and its rice homolog, have shown activity without the removal of the putative prosegment [51,52]. More studies have to be conducted to fully understand the inactivation mechanisms of plant APs. Soares and colleagues have recently proposed that the inactive form of APs occurs because the active site is blocked by the prosegment alone or by the prosegment together with the mature N-terminal and the flap. In contrast, precursors of cardosin A and B are active before undergoing the proteolytic process that removes prosegment, hence probably do not share the inactivation mechanism described above [20].
Considering the APs’ subcellular location, these proteases are found in various cellular compartments. Typical APs are mostly found in vacuoles, such as in the case of APs from barley [53], castor bean (Ricinus communis) [54], and Arabidopsis [55]. To a less extent, typical APs are also located in the extracellular space, such as in the case of tomato (Solanum lycopersicum) [16] and tobacco (Nicotiana tabacum) APs [56]. Atypical APs are widely distributed in the cell: Arabidopsis PCS1, ASPG1, and ASPR1 are located in the endoplasmic reticulum (ER) [29,30,57]; UNDEAD AP in mitochondria [58]; and CND41 and NANA in the chloroplast [27,40]. Rice OsAP65 is located in pre-vacuolar compartments [28]; nepenthesins and Arabidopsis AED1 and CDR1 are distributed in the extracellular space [33,59,60]. Arabidopsis A36 and A39 APs were found to be located in the plasma membrane as anchored proteins [61].
Plant APs are involved in many biological functions, particularly in developmental processes, such as chloroplast homeostasis and protein turnover [27,40], as well as in programmed cell death (PCD) and cell survival [62]. Developmentally controlled plant cell death is initiated through hormonal signaling, which in turn leads to the accumulation of reactive oxygen species (ROS) and transcriptional activation of PCD-related genes, such as proteases and nucleases. PCD can have different outcomes, such as senescence, the death of cells no longer required, or the creation of tissues that assume structural storage functions [63].
3. Aspartic Proteases Involved in Plant Defense Responses
Plants are sessile organisms, exposed to numerous biotic stresses and adverse environmental conditions [64]. Plant aspartic proteases were demonstrated to be involved in response to both abiotic [30,31,32] and biotic [33,34,35] environmental stressors.
3.1. Response to Abiotic Stress
Plant growth and productivity have been impaired due to abiotic stresses, such as drought, heat, cold, and excess of salt and metals in the soil [65]. Under drought stress, an aspartic protease from common bean (Phaseolus vulgaris), PvAP1, was shown to be up-regulated earlier in the leaves of a drought-susceptible cultivar than in the resistant cultivar (Table 1; Reference [66]). The Arabidopsis aspartic protease in guard cell 1 (ASPG1) was firstly shown to be involved in drought stress resistance, in addition to its role in the degradation of seed storage proteins [30]. Arabidopsis mutants overexpressing ASPG1 were shown to recover more efficiently from drought, as ASPG1 lead to a significant increase in abscisic acid (ABA) sensitivity by guard cells and antioxidant enzymes activation, preventing Arabidopsis plants from oxidative damage [30]. A gene homologous to ASPG1 from potato was shown to be down-regulated under drought and up-regulated upon re-watering, suggesting also a role in drought stress (Table 1; Reference [67]).

Table 1.
Aspartic proteases involved in abiotic stress responses.
Recently, an Arabidopsis aspartic protease, APA1, has been implicated also in drought tolerance. Plants overexpressing the apa1 gene (OE-APA1) were more tolerant to mild water deficit (MWD) than WT plants, while apa1 line was more susceptible (Table 1; Reference [31]). OE-APA1 lines exhibited more total leaf area, less chlorophyll content and shortened principal root length under MWD treatment. Analysis of stomatal behavior showed that OE-APA1 plants presented reduced stomatal pore aperture and reduced stomatal index [31]. Since ABA regulates stomatal closure, aperture pore size was determined upon ABA treatment. The stomata of OE-APA1 plants was already closed before ABA treatment. This work suggested that APA1 has a role in stomatal behavior via regulation of the ABA signaling pathway [31]. An aspartic protease gene from buckwheat (Fagopyrum esculentum), FeAP9, was found to be up-regulated in leaves in response to numerous abiotic stresses, including dark, drought, wounding, and UV-B light (Table 1; Reference [69]). Moreover, when pineapple fruit (Ananas comosus) is exposed to a chilling injury, it develops brown symptoms known as blackheart [70]. Gene expression analysis of the pineapple fruit under postharvest chilling treatment showed that aspartic protease AcAP1 was up-regulated in a variety resistant to blackheart and down-regulated on a susceptible one (Table 1; Reference [70]). Thus, it is expected that AcAP1 is involved in resistant mechanisms concerning chilling stress [70]. Transgenic Arabidopsis plants overexpressing the grapevine aspartic protease AP17 showed salt- and drought-tolerance as transgenic seeds had higher levels of germination and transgenic seedlings roots were longer under osmotic stress [32]. In addition, the plasma membranes of the transgenic seedlings suffered less damage, and the genes involved in ABA biosynthesis were up-regulated (Table 1; Reference [32]). These results suggest that AP17 is a key component for maintaining the integrity of the membrane and may be involved in ABA biosynthetic pathway [32].
Heavy metals, such as iron, copper, nickel, mercury, and cadmium, are one of the environmental pollutants affecting plant growth. Despite the fact that plants have the ability to tolerate certain concentrations of these metals, when a certain level is exceeded, it causes toxicity, leading to the generation of ROS [71]. Although plant extracts have been reported to prevent heavy-metal-induced stress [72], only cysteine proteases, such as caspases and vacuolar processing enzymes, have been shown to be involved in this process [73,74,75].
3.2. Aspartic Proteases Involvement in Plant–Pathogen Interaction
The first clue of the involvement of plant APs in biotic stress was found in tomato leaves, where pathogenesis-related (PR) proteins secreted upon pathogen challenge were degraded by an extracellular aspartic protease, preventing its over accumulation [16]. Three years later, the same function was proposed for an aspartic protease found in tobacco leaves [56]. The tomato aspartyl protease is thought to be responsible for the cleavage of PR-1b [76]. PR-1b cleavage releases a peptide, CAP-derived peptide 1 (CAPE1), that induces the expression of genes involved in stress and defense responses, innate immunity, and systemic acquired resistance (SAR) [77]. It was suggested that CAPE1 may act as a novel damage-associated molecular patterns (DAMP) linked to both jasmonic acid (JA) and salicylic acid (SA) pathways and SAR activation [77]. An aspartic protease gene detected in tomato leaves (LeAspP), in response to wounding and treatments with systemin and methyl jasmonate (MeJA), was also shown to be systemically induced, suggesting that this AP plays a role in defense against pathogens [78]. In potato tubers, the Solanum tuberosum aspartic protease 1 (StAP1) was identified in immunological analysis of intercellular washing fluids of potato tubers has presenting a higher accumulation in a resistant cultivar than in a susceptible one upon Phytophthora infestans infection [79]. Moreover, western blot analysis of an AP from potato leaves, Solanum tuberosum aspartic protease 3 (StAP3), showed a higher accumulation in potato resistant cultivar upon P. infestans infection [80]. StAsp, also isolated from leaves, presented a higher expression in the resistant cultivar after infection with P. infestans [49]. These results suggest that several StAPs may be involved in plant immunity response. In a recent study focusing on the soybean–Phytophthora sojae interaction, a secreted soybean AP (GmAP5) has been described to bind and degrade the pathogen effector PsXEG1, an apoplastic endoglucanase [81]. The cleavage of this effector severely affects P. sojae virulence (Figure 3a). Soybean has another layer of defense towards this P. sojae effector. The apoplastic inhibitor protein GmGIP1 bind to the PsXEG1 effector, reducing its enzymatic activity and, thus, the pathogen’s virulence. To counterattack this, P. sojae N-glycosilates this effector, protecting it from the proteolytic activity of GmAP5, and secretes a decoy effector, PsXLP1, that binds more tightly to GmGIP1 [81,82]. This intricate system shows the different layers of defense and offense in plant–pathogen interactions.
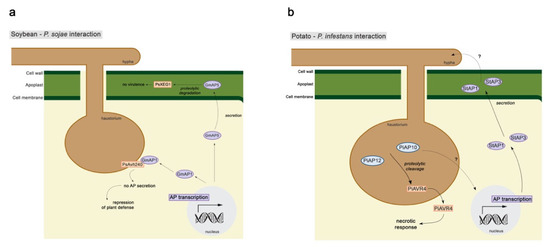
Figure 3.
Schematic representation of (a) soybean–P. sojae and (b) potato–P. infestans interactions. Purple ellipses, plant aspartic proteases; orange boxes, pathogen effectors; dashed arrows, unknown processes.
More aspartic proteases were described to induce systemic defense responses in plants. The Arabidopsis CDR1 gene was identified while studying a gain-of-function dominant mutation that presented a phenotype of enhanced resistance to the bacterial pathogen Pseudomonas syringae [33]. Arabidopsis CDR1 mutants exhibited a phenotype that mimics constitutive activation of SAR, including the accumulation of high levels of SA; SAR transcripts marker genes, such as PR1 and PR2; and oxidative bursts resulting from hypersensitive response (HR). It was hypothesized that CDR1 released a peptide elicitor that may function as a mobile SAR signal [33]. Ectopic expression of the rice ortholog CDR1 (OsCDR1), in both Arabidopsis and rice, conferred enhanced resistance to Pseudomonas syringae, Hyaloperonospora arabidopsidis, Xanthomonas oryzae, and Magnaporthe oryzae, which is correlated with the enhanced PR gene expression [83]. Infiltration of Arabidopsis leaves with purified OsCDR1–GST fusion protein induced PR2 expression. Interestingly, the expression of this pathogen related protein was also identified in non-inoculated neighbor leaves, demonstrating that OsCDR1 induces systemic defense [52]. Conversely, apoplastic enhanced disease susceptibility 1 aspartic protease (AED1) has been described as having a role in SAR repression [59]. In enhanced disease susceptibility 1 (eds1) mutant plants, which are SAR defected, AED1 was found in a proteome profiling analysis of the extracellular fluid in response to the P. syringae effector AvrRpm1 [59]. The AED1 transcript accumulation was found to be induced by this pathogen both locally and systemically in WT and eds1 mutants, despite AED1 content was much lower in the eds1 background [59]. These results suggest that systemic accumulation of AED1 in response to P. syringae depends on EDS1 protein and, thus, is correlated with SAR [59]. Additionally, overexpression of AED1 led to the repression of both SAR- and SA-induced resistance without affecting P. syringae growth in healthy plants. These findings support the hypothesis that AED1 might be part of a homeostatic mechanism to limit SAR signaling and to reallocate resources from defense to plant growth [59]. Another recent report has highlighted the importance of aspartic proteases in plant defense by demonstrating its antibacterial function. Wang and colleagues showed that A. thaliana secreted aspartic protease 1 and 2 (SAP1 and SAP2) are able to cleave a highly conserved bacterial protein, MucD. The cleavage of MucD inhibits the growth of P. syringae in planta and in vitro, showing the importance of antibacterial mechanisms in plant defense [84].
The activity of the rice aspartic protease 77 gene (OsAP77) in rice transgenic lines was induced upon infection by M. oryzae, X. oryzae, and cucumber mosaic virus (CMV) in vascular tissues [34]. In addition, rice transgenic plants treated with SA, isonicotinic acid, hydrogen peroxide, and ABA showed an increased level of the reporter gene GUS. These results suggest that OsAP77 has a positive role in pathogens defense [34]. Moreover, two APs were shown to be induced in the rice apoplast upon M. oryzae infection, suggesting that these proteins may act as signal transductors apart from their hydrolytic activity [85]. The Bcl-2 associated athanogene protein 6 (BAG6) participates in limiting pathogen colonization and spread of the necrotrophic fungus Botritis cinerea by inducing autophagy [86]. For autophagy to occur, full-length BAG6 needs to be activated by protease processing. Recently, Arabidopsis aspartyl protease cleaving BAG (APCB1) was demonstrated to be essential in BAG6 proteolytic processing [87]. The apcb1 mutants exhibited enhanced susceptibility similar to the bag6 mutants. Mutation of the catalytic site of APCB1 led to the absence of BAG6 cleavage. This aspartic protease appears to be BAG6 specific, once the two unrelated Arabidopsis aspartyl proteases were unable to cleave BAG6 in apcb1 mutant line, causing loss of resistance [87].
Previous studies have also shown that the grapevine aspartic protease 13 (AP13) gene was up-regulated in Chinese wild Vitis quinquangularis cv. “Shang-24” following Erysiphe necator infection. It was also up-regulated in V. labrusca × V. vinifera cv. “Kyoho” following a treatment with SA, suggesting that this gene may confer resistance to biotrophic pathogens [25]. Further studies analyzed the expression levels of AP13 upon inoculation with B. cinerea and treatments with hormones involved in plant defense. AP13 from V. quinquangularis cv. “Shang-24” was shown to be up-regulated after both SA and ethylene (ET) treatment and was down-regulated upon JA and MeJA treatment and B. cinerea infection. These results suggest that AP13 promotes the SA dependent signal transduction pathway and suppresses the JA signal transduction pathway. The ectopic expression of AP13 in Arabidopsis improved the resistance of transgenic plants to E. necator and the bacterial pathogen P. syringae but reduced the resistance to B. cinerea [35].
The perception of microbial or damage signals by plants’ receptors initiates a response that leads to the production of peptides and small molecules that enhance immunity responses [88]. Several studies have highlighted the antimicrobial activity of plant proteases. Potato-isolated APs were shown to inhibit both Phytophthora infestans and Fusarium solani growth; salpicornin, isolated from Salpichroa origanifolia fruits, was shown to inhibit Fusarium solani growth; and cirsin, isolated from thistle plants also presented antifungal activity against Lewia infectoria, Alternaria alternata, and Drechslera biseptata [79,80,89,90].
Monomeric aspartic proteases from S. tuberosum were reported to have bifunctional activity, both proteolytic and antimicrobial [91]. The plant-specific insert of S. tuberosum aspartic protease 1 (StAsp-PSI) is able to interact with pathogens’ spore surface, inducing damage in its plasma membranes and causing death in a dose-dependent manner [91]. In vitro, StAsp-PSI was able to kill spores of P. infestans and Fusarium solani by direct interaction of the protein with the pathogens’ cell membrane, leading to an increased permeability and lysis [91]. A recent study has suggested that the PSI from S. tuberosum facilitates membrane fusion at acidic pH, while the mature AP degrades pathogenic proteins in the extracellular space [92]. Arabidopsis transgenic lines expressing the potato PSI increased plant resistance to B. cinerea infection through direct cytotoxic activity and induction of gene expression associated to the regulation of JA and SA pathways, such as PDF1.2 and PR-1 [50]. Moreover, cytotoxic analysis of salpichroin and circin activity suggested it was not related to proteolysis but to membrane permeabilization of pathogen conidia. Altogether, these studies suggest that the PSI domain may be involved in membrane permeabilization [89]. Therefore, the constitutive expression of these APs’ PSI could potentially be used as a strategy to cope with plant pathogens [50].
4. Pathogen Aspartic Proteases
Proteases from pathogens can also play an important role in the mechanisms of virulence during infection, by participating in the degradation of the host’s physical barriers and combating the host’s defense mechanisms [93]. However, aspartic proteases’ role in virulence of plant pathogens is still poorly studied [93,94,95].
Botrytis cinerea, a necrotrophic fungus that causes the grey mold disease in many plant species, has a significant content of secreted APs upon successful infection of host [96]. In 1990, APs were first established as possible B. cinerea virulence factors when the supplementation of inoculum with pepstain A (AP inhibitor) limited infection [97]. BcAp5, BcAP8, BcAP9 and BcAP14 were observed to be up-regulated in the first hours of infection upon grape berry infection [98]. A recent report has also shown that, in the first 24 hours of infection, there is an increase in the transcription of phytotoxins and cell wall degrading enzymes [99]. Taken together, these studies show that APs are a crucial tool for the B. cinerea infection strategy.
A genome analysis of three Phytophthora species (P. infestans, P. sojae, and P. ramorum) reported, in 2011, that these pathogens present 5 clans and 12 families of APs and that they are all predicted to be membrane-bound [100]. APs have also been identified in the secretome of four Phytophthora species (P. infestans [101] and P. pseudosyringae—forest pathogen; P. chamydospora and P. gonapodyides—frequent in aquatic habitats; Reference [102]). In a recent study, P. infestans (Pi) transformants, overexpressor, and silenced lines for APs, were characterized to determine if P. infestans aspartic proteases (PiAPs) play a role in virulence. PiAP10 and PiAP12 silenced lines showed a reduction in mycelial growth and sporangia production and low infection efficiency on inoculated potato leaves. Activity assays suggest that both lines were capable to cleave the P. infestans Arginine-x-Leucine-Arginine effector AVR4. PiAP11 silenced transformants did not show any reduction. These results suggest that PiAP10 and PiAP12 play a role in virulence (Figure 3b) [103]. An effector of P. sojae (PsAvh240) has been recently described to interact with an AP (GmAP1) from a resistant soybean cultivar (Figure 3a). This interaction suppresses GmAP1 secretion to the apoplast, limiting soybean apoplastic immunity and, thus, plant defense [104].
The infection mechanism of Fusarium proliferatum, a pathogen that causes fungal keratitis in several crops [105], has been shown to be pH-dependent [106]. At an alkaline pH, the infection of bananas by F. proliferatum is hampered. At the same time, in these conditions, a secreted AP from this pathogen is down-regulated. Although further studies are needed, the authors suggest that this AP may be crucial for the infection process of F. proliferatum [106]. A wheat fungal pathogen, Zymoseptoria tritici, has been described to up regulate extracellular APs during its asymptomatic biotrophic phase [107]. Further studies could confirm that these APs may be effector genes that suppress wheat apoplastic immunity. During infection of sunflower cotyledons by Sclerotinia sclerotiorum, a necrotrophic fungus, several acid proteases were shown to be secreted and the aspS gene, encoding for an aspartic protease, was expressed at the early stages of infection [108]. Studies have shown that an acid AP from Fusarium culmorum presented a role in the pathogen infection due to its capacity to degrade plant inhibitor proteins such as bean polygalacturonase inhibitor and soybean trypsin inhibitor [109]. Overexpression of endothiapepsin (Epn), an AP secreted by the Cryphonectria parasitica fungus responsible for chestnut blight, leads to enhanced necrosis on chestnut bark and wood tissues, suggesting its involvement in pathogenicity, as well [110]. The extracellular aspartic proteases Eap1 from Sporisorium reilianum (that causes maize and sorghum head smut) and APSm1 from Stenocarpella maydis (that causes diplodia ear and stalk rot in maize) were purified and characterized [111,112]. Strong evidences highlight their involvement as key components of the biological and infection cycles of the pathogen [111,112]. Ralstonia solenacearum, a pathogenic bacterium causing bacterial wilt on many solenaceous crops, secretes an AP (Rsa1) that is able to elicit HR response in potato and has an important role in the virulence of this pathogen [113]. The proteases of the biotrophic pathogen Cladosporium fulvum, which causes tomato leaf mold disease, were analyzed in a recent study, by transcriptome and proteome analysis. In total, 14 out of the 59 predicted protease genes were expressed in vitro and in planta, and one of proteins (CfPro7) was predicted to be secreted [114]. During the infection of apples by Penicillium expansum, an AP (PEX2_009280) was reported to be up-regulated, suggesting its possible role in this pathogen infection mechanism [115]. Another apple fruit pathogen, Colletotricum acutatum, has been shown to secrete an AP during infection. A protease inhibitor extracted from apple fruits, that showed a similar activity to a commercial AP inhibitor, inhibited C. acucatum growth in vivo and in vitro, showing the importance of this AP in the virulence of this pathogen [116].
The studies reviewed here emphasize the diversity of pathogens that rely on the proteolytic activity of aspartic proteases for the success of their infection process. However, further studies are needed to deepen our knowledge on the role of proteases, as well as protease inhibitors, as virulence factors.
5. Conclusions and Future Perspectives
In the past years, several studies regarding the role of plant aspartic proteases in different cell functions have been reported. Although there is a lot of information about plant typical APs, there is still a need for a better understanding of the structure of atypical and nucellin-like APs, as well as their substrates, interacting proteins, and proteolytic activity. Since the end of the 19th century, the study of the involvement of APs in plant development has increased. Although many of the functions given to APs remain hypothetical, studies using reverse engineering tools and biochemical studies are essential to understand the detail biological function. The knowledge about plant–pathogen interaction has risen, as well as the involvement of aspartic proteases in this interaction. It is clear that APs have an important role in plant defense against a wide range of pathogens. On the other hand, aspartic proteases from pathogens are still poorly studied. Research within a variety of molecular, genetic, and biochemical approaches will contribute to fully address these questions and finally understand the regulation mechanisms regarding plant aspartic proteases role, particularly in plant development and pathogen interaction.
Funding
The present work was supported by Portuguese Foundation for Science and Technology (FCT/MCTES/PIDDAC, Portugal), BioISI (UIDB/04046/2020 and UIDP/04046/2020), PTDC/BIA-BQM/28539/2017 and IF/00819/2015.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
We thank to Joana Figueiredo and Ana Rita Cavaco for the critical reading and suggestions on this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schwann, T. Ueber das Wesen des Verdauungsprocesses. Ann. Phys. Chem. 1836, 114, 358–364. [Google Scholar] [CrossRef]
- Darwin, C. Insectivorous Plants; Murray, J., Ed.; John Murray: London, UK, 1875. [Google Scholar]
- Northrop, J.H. Crystalline pepsin: I. Isolation and tests of purity. J. Gen. Physiol. 1930, 13, 739–766. [Google Scholar] [CrossRef] [PubMed]
- Northrop, J.H. Crystalline Enzymes. The Chemistry of Pepsin, Trypsin, and Bacteriophage; Columbia University Press: New York, NY, USA, 1939. [Google Scholar]
- Herriott, R.M. Pepsinogen and pepsin. J. Gen. Physiol. 1962, 45, 57–76. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, H. Enzyme Inhibitors of Microbial Origin; University of Tokyo Press: Tokyo, Japan, 1972. [Google Scholar]
- Mutlu, A.; Gal, S. Plant aspartic proteinases: Enzymes on the way to a function. Physiol. Plant. 1999, 105, 569–576. [Google Scholar] [CrossRef]
- Fruton, J.S. A history of pepsin and related enzymes. Q. Rev. Biol. 2002, 77, 127–147. [Google Scholar] [CrossRef]
- Tang, J.; Sepulveda, P.; Marciniszyn, J.; Chen, K.C.; Huang, W.Y.; Tao, N.; Liu, D.; Lanier, J.P. Amino acid sequence of porcine pepsin. Proc. Natl. Acad. Sci. USA 1973, 70, 3437–3439. [Google Scholar] [CrossRef]
- Fruton, J.S. Aspartyl proteinases. In Hydrolytic Enzymes; Brocklehurst, K., Neuberger, A., Eds.; Elsevier: Amsterdam, The Netherlands, 1987; p. 423. ISBN 9780080860756. [Google Scholar]
- Szecsi, P.B. The aspartic proteases. Scand. J. Clin. Lab. Investig. 1992, 52, 2–22. [Google Scholar] [CrossRef]
- Dol, E.; Shibata, D.; Matoba, T.; Yoneza, W.A.D. Characterization of pepstatin-sensitive acid protease in resting rice seeds. Agric. Biol. Chem. 1980, 44, 741–747. [Google Scholar] [CrossRef]
- Polanowski, A.; Wilusz, T.; Kolaczkowska, M.K.; Wieczorek, M.; Wilimowska-Pelc, A. Purification and characterization of aspartic proteinases from Cucumis sativus and Cucurbita maxima seeds. In Aspartic Proteinases and Their Inhibitors; Kostka, V., Ed.; Springer: Boston, MA, USA, 1985; pp. 49–52. [Google Scholar]
- Dunaevsky, Y.E.; Sarbakanova, S.T.; Belozersky, M.A. Wheat seed carboxypeptidase and joint action on gliadin of proteases from dry and germinating seeds. J. Exp. Bot. 1989, 40, 1323–1329. [Google Scholar] [CrossRef]
- Sarkkinen, P.; Kalkkinen, N.; Tilgmann, C.; Siuro, J.; Kervinen, J.; Mikola, L. Aspartic proteinase from barley grains is related to mammalian lysosomal cathepsin D. Planta 1992, 186, 317–323. [Google Scholar] [CrossRef]
- Rodrigo, I.; Vera, P.; Conejero, V. Degradation of tomato pathogenesis-related proteins by an endogenous 37-kDa aspartyl endoproteinase. Eur. J. Biochem. 1989, 184, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Runeberg-Roos, P.; Tormakangas, K.; Ostman, A. Primary structure of a barley-grain aspartic proteinase. A plant aspartic proteinase resembling mammalian cathepsin D. Eur. J. Biochem. 1991, 202, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, N.D.; Barrett, A.J. MEROPS: The peptidase database. Nucleic Acids Res. 1999, 27, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res. 2018, 46, D624–D632. [Google Scholar] [CrossRef]
- Simões, I.; Faro, C. Structure and function of plant aspartic proteinases. Eur. J. Biochem. 2004, 271, 2067–2075. [Google Scholar] [CrossRef]
- Beers, E.P.; Jones, A.M.; Dickerman, A.W. The S8 serine, C1A cysteine and A1 aspartic protease families in Arabidopsis. Phytochemistry 2004, 65, 43–58. [Google Scholar] [CrossRef]
- Faro, C.; Gal, S. Aspartic Proteinase Content of the Arabidopsis Genome. Curr. Protein Pept. Sci. 2005, 6, 493–500. [Google Scholar] [CrossRef]
- Dunn, B.M. Structure and mechanism of the pepsin-like family of aspartic peptidases. Chem. Rev. 2002, 102, 4431–4458. [Google Scholar] [CrossRef]
- Chen, J.; Ouyang, Y.; Wang, L.; Xie, W.; Zhang, Q. Aspartic proteases gene family in rice: Gene structure and expression, predicted protein features and phylogenetic relation. Gene 2009, 442, 108–118. [Google Scholar] [CrossRef]
- Guo, R.; Xu, X.; Carole, B.; Li, X.; Gao, M.; Zheng, Y.; Wang, X. Genome-wide identification, evolutionary and expression analysis of the aspartic protease gene superfamily in grape. BMC Genom. 2013, 14, 554. [Google Scholar] [CrossRef]
- Cao, S.; Guo, M.; Wang, C.; Xu, W.; Shi, T.; Tong, G.; Zhen, C.; Cheng, H.; Yang, C.; Elsheery, N.I.; et al. Genome-wide characterization of aspartic protease (AP) gene family in Populus trichocarpa and identification of the potential PtAPs involved in wood formation. BMC Plant Biol. 2019, 19, 276. [Google Scholar] [CrossRef] [PubMed]
- Paparelli, E.; Gonzali, S.; Parlanti, S.; Novi, G.; Giorgi, F.M.; Licausi, F.; Kosmacz, M.; Feil, R.; Lunn, J.E.; Brust, H.; et al. Misexpression of a chloroplast aspartyl protease leads to severe growth defects and alters carbohydrate metabolism in Arabidopsis. Plant Physiol. 2012, 160, 1237–1250. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhao, X.; Cheng, K.; Jiang, Y.; Ouyang, Y.; Xu, C.; Li, X.; Xiao, J.; Zhang, Q. OsAP65, a rice aspartic protease, is essential for male fertility and plays a role in pollen germination and pollen tube growth. J. Exp. Bot. 2013, 64, 3351–3360. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.; Niedermaier, S.; Faro, R.; Loos, A.; Manadas, B.; Faro, C.; Huesgen, P.F.; Cheung, A.Y.; Simões, I. An atypical aspartic protease modulates lateral root development in Arabidopsis thaliana. J. Exp. Bot. 2019, 70, 2157–2171. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Xiong, W.; Ye, T.; Wu, Y. Overexpression of the aspartic protease ASPG1 gene confers drought avoidance in Arabidopsis. J. Exp. Bot. 2012, 63, 2579–2593. [Google Scholar] [CrossRef] [PubMed]
- Sebastián, D.I.; Fernando, F.D.; Raúl, D.G.; Gabriela, G.M. Overexpression of Arabidopsis aspartic protease APA1 gene confers drought tolerance. Plant Sci. 2020, 292, 110406. [Google Scholar] [CrossRef]
- Guo, R.; Zhao, J.; Wang, X.X.; Guo, C.; Li, Z.; Wang, Y.; Wang, X.X. Constitutive expression of a grape aspartic protease gene in transgenic Arabidopsis confers osmotic stress tolerance. Plant Cell Tissue Organ Cult. 2015, 121, 275–287. [Google Scholar] [CrossRef]
- Xia, Y.; Suzuki, H.; Borevitz, J.; Blount, J.; Guo, Z.; Patel, K.; Dixon, R.A.; Lamb, C. An extracellular aspartic protease functions in Arabidopsis disease resistance signaling. EMBO J. 2004, 23, 980–988. [Google Scholar] [CrossRef]
- Alam, M.M.; Nakamura, H.; Ichikawa, H.; Miyao, A.; Hirochika, H.; Kobayashi, K.; Yamaoka, N.; Nishiguchi, M. Response of an aspartic protease gene OsAP77 to fungal, bacterial and viral infections in rice. Rice 2014, 7, 9. [Google Scholar] [CrossRef]
- Guo, R.; Tu, M.; Wang, X.; Zhao, J.; Wan, R.; Li, Z.; Wang, Y.; Wang, X. Ectopic expression of a grape aspartic protease gene, AP13, in Arabidopsis thaliana improves resistance to powdery mildew but increases susceptibility to Botrytis cinerea. Plant Sci. 2016, 248, 17–27. [Google Scholar] [CrossRef]
- Faro, C.; Ramalho-Santos, M.; Veríssimo, P.; Pissarra, J.; Frazão, C.; Costa, J.; Lin, X.-L.; Tang, J.; Pires, E. Structural and functional aspects of cardosins. In Advances in Experimental Medicine and Biology; Springer LLC: New York, NY, USA, 1998; Volume 436, pp. 423–433. [Google Scholar]
- Kervinen, J.; Tobin, G.J.; Costa, J.; Waugh, D.S.; Wlodawer, A.; Zdanov, A. Crystal structure of plant aspartic proteinase prophytepsin: Inactivation and vacuolar targeting. EMBO J. 1999, 18, 3947–3955. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.; Ribeiro Carlton, S.M.; Simões, I. Atypical and nucellin-like aspartic proteases: Emerging players in plant developmental processes and stress responses. J. Exp. Bot. 2019, 70, 2059–2076. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Foolad, M.R. Molecular organization of a gene in barley which encodes a protein similar to aspartic protease and its specific expression in nucellar cells during degeneration. Plant Mol. Biol. 1997, 35, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Murakami, S.; Shoji, T.; Yoshida, S.; Yamada, Y.; Sato, F. A novel protein with DNA binding activity from tobacco chloroplast nucleoids. Plant Cell 1997, 9, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Cheung, L.K.Y.; Dupuis, J.H.; Dee, D.R.; Bryksa, B.C.; Yada, R.Y. Roles of Plant-Specific Inserts in Plant Defense. Trends Plant Sci. 2020. [Google Scholar] [CrossRef]
- Park, H.; Kusakabe, I.; Sakakibara, Y.; Kobayashi, H. Autoproteolytic processing of aspartic proteinase from sunflower seeds. Biosci. Biotechnol. Biochem. 2001, 65, 702–705. [Google Scholar] [CrossRef]
- Castanheira, P.; Samyn, B.; Sergeant, K.; Clemente, J.C.; Dunn, B.M.; Pires, E.; Van Beeumen, J.; Faro, C. Activation, proteolytic processing, and peptide specificity of recombinant cardosin A. J. Biol. Chem. 2005, 280, 13047–13054. [Google Scholar] [CrossRef]
- Feijoo-Siota, L.; Rama, J.L.R.; Sánchez-Pérez, A.; Villa, T.G. Expression, activation and processing of a novel plant milk-clotting aspartic protease in Pichia pastoris. J. Biotechnol. 2018, 268, 28–39. [Google Scholar] [CrossRef]
- Ramalho-Santos, M.; Verissimo, P.; Cortes, L.; Samyn, B.; Van Beeumen, J.; Pires, E.; Faro, C. Identification and proteolytic processing of procardosin A. Eur. J. Biochem. 1998, 255, 133–138. [Google Scholar] [CrossRef]
- Chen, H.-J.; Huang, Y.-H.; Huang, G.-J.; Huang, S.-S.; Chow, T.-J.; Lin, Y.-H. Sweet potato SPAP1 is a typical aspartic protease and participates in ethephon-mediated leaf senescence. J. Plant Physiol. 2015, 180, 1–17. [Google Scholar] [CrossRef]
- Guevara, M.G.; Oliva, C.R.; Machinandiarena, M.; Daleo, G.R. Purification and properties of an aspartic protease from potato tuber that is inhibited by a basic chitinase. Physiol. Plant. 1999, 106, 164–169. [Google Scholar] [CrossRef]
- Guevara, M.G.; Daleo, G.R.; Oliva, C.R. Purification and characterization of an aspartic protease from potato leaves. Physiol. Plant. 2001, 112, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Guevara, M.G.; Almeida, C.; Mendieta, J.R.; Faro, C.J.; Veríssimo, P.; Pires, E.V.; Daleo, G.R. Molecular cloning of a potato leaf cDNA encoding an aspartic protease (StAsp) and its expression after P. infestans infection. Plant Physiol. Biochem. 2005, 43, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Frey, M.E.; D’Ippolito, S.; Pepe, A.; Daleo, G.R.; Guevara, M.G. Transgenic expression of plant-specific insert of potato aspartic proteases (StAP-PSI) confers enhanced resistance to Botrytis cinerea in Arabidopsis thaliana. Phytochemistry 2018, 149, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Simões, I.; Faro, R.; Bur, D.; Faro, C. Characterization of recombinant CDR1, an Arabidopsis aspartic proteinase involved in disease resistance. J. Biol. Chem. 2007, 282, 31358–31365. [Google Scholar] [CrossRef]
- Prasad, B.D.; Creissen, G.; Lamb, C.; Chattoo, B.B. Heterologous expression and characterization of recombinant OsCDR1, a rice aspartic proteinase involved in disease resistance. Protein Expr. Purif. 2010, 72, 169–174. [Google Scholar] [CrossRef]
- Runeberg-Roos, P.; Kervinen, J.; Kovaleva, V.; Raikhel, N.V.; Gal, S. The aspartic proteinase of barley is a vacuolar enzyme that processes probarley lectin in vitro. Plant Physiol. 1994, 105, 321–329. [Google Scholar] [CrossRef][Green Version]
- Hiraiwa, N.; Kondo, M.; Nishimura, M.; Hara-Nishimura, I. An aspartic endopeptidase is involved in the breakdown of propeptides of storage proteins in protein-storage vacuoles of plants. Eur. J. Biochem. 1997, 246, 133–141. [Google Scholar] [CrossRef]
- Mutlu, A.; Chen, X.; Reddy, S.M.; Gal, S. The aspartic proteinase is expressed in Arabidopsis thaliana seeds and localized in the protein bodies. Seed Sci. Res. 1999, 9, 75–84. [Google Scholar] [CrossRef]
- Rodrigo, I.; Vera, P.; Van Loon, L.C.; Conejero, V. Degradation of tobacco pathogenesis-related proteins: Evidence for conserved mechanisms of degradation of pathogenesis-related proteins in plants. Plant Physiol. 1991, 95, 616–622. [Google Scholar] [CrossRef]
- Ge, X.; Dietrich, C.; Matsuno, M.; Li, G.; Berg, H.; Xia, Y. An Arabidopsis aspartic protease functions as an anti-cell-death component in reproduction and embryogenesis. EMBO Rep. 2005, 6, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Phan, H.A.; Iacuone, S.; Li, S.F.; Parish, R.W. The MYB80 transcription factor is required for pollen development and the regulation of tapetal programmed cell death in Arabidopsis thaliana. Plant Cell 2011, 23, 2209–2224. [Google Scholar] [CrossRef] [PubMed]
- Breitenbach, H.H.; Wenig, M.; Wittek, F.; Jordá, L.; Maldonado-Alconada, A.M.; Sarioglu, H.; Colby, T.; Knappe, C.; Bichlmeier, M.; Pabst, E.; et al. Contrasting roles of the apoplastic aspartyl protease Apoplastic, Enhanced Disease Susceptibility1-Dependent1 and Legume Lectin-Like Protein1 in Arabidopsis systemic acquired resistance. Plant Physiol. 2014, 165, 791–809. [Google Scholar] [CrossRef] [PubMed]
- Athauda, S.B.P.; Matsumoto, K.; Rajapakshe, S.; Kuribayashi, M.; Kojima, M.; Kubomura-Yoshida, N.; Iwamatsu, A.; Shibata, C.; Inoue, H.; Takahashi, K. Enzymic and structural characterization of nepenthesin, a unique member of a novel subfamily of aspartic proteinases. Biochem. J. 2004, 381, 295–306. [Google Scholar] [CrossRef]
- Gao, H.; Li, R.; Guo, Y. Arabidopsis aspartic proteases A36 and A39 play roles in plant reproduction. Plant Signal. Behav. 2017, 12, e1304343. [Google Scholar] [CrossRef][Green Version]
- Stael, S.; Van Breusegem, F.; Gevaert, K.; Nowack, M.K. Plant Proteases and Programmed Cell Death; Oxford University Press: Oxford, UK, 2019; Volume 70, pp. 1991–1995. [Google Scholar]
- Huysmans, M.; Lema, A.S.; Coll, N.S.; Nowack, M.K. Dying two deaths—Programmed cell death regulation in development and disease. Curr. Opin. Plant Biol. 2017. [Google Scholar] [CrossRef]
- Andersen, E.; Ali, S.; Byamukama, E.; Yen, Y.; Nepal, M. Disease resistance mechanisms in plants. Genes 2018, 9, 339. [Google Scholar] [CrossRef]
- Zhu, J.-K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Contour-Ansel, D.; Torres-Franklin, M.L.; Zuily-Fodil, Y.; de Carvalho, M.H.C. An aspartic acid protease from common bean is expressed “on call” during water stress and early recovery. J. Plant Physiol. 2010, 167, 1606–1612. [Google Scholar] [CrossRef]
- Gong, L.; Zhang, H.; Gan, X.; Zhang, L.; Chen, Y.; Nie, F.; Shi, L.; Li, M.; Guo, Z.; Zhang, G.; et al. Transcriptome profiling of the potato (Solanum tuberosum L.) plant under drought stress and water-stimulus conditions. PLoS ONE 2015, 10, e0128041. [Google Scholar] [CrossRef]
- Gong, P.; Riemann, M.; Dong, D.; Stoeffler, N.; Gross, B.; Markel, A.; Nick, P. Two grapevine metacaspase genes mediate ETI-like cell death in grapevine defence against infection of Plasmopara viticola. Protoplasma 2019, 256, 951–969. [Google Scholar] [CrossRef] [PubMed]
- Timotijević, G.S.; Milisavljević, M.D.; Radović, S.R.; Konstantinović, M.M.; Maksimović, V.R. Ubiquitous aspartic proteinase as an actor in the stress response in buckwheat. J. Plant Physiol. 2010, 167, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Raimbault, A.K.; Zuily-Fodil, Y.; Soler, A.; Cruz de Carvalho, M.H. A novel aspartic acid protease gene from pineapple fruit (Ananas comosus): Cloning, characterization and relation to postharvest chilling stress resistance. J. Plant Physiol. 2013, 170, 1536–1540. [Google Scholar] [CrossRef]
- Yanık, F.; Çetinbaş-Genç, A.; Vardar, F. Abiotic stress–induced programmed cell death in plants. In Plant Life Under Changing Environment; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1–24. [Google Scholar]
- Tortora, F.; Notariale, R.; Maresca, V.; Good, K.V.; Sorbo, S.; Basile, A.; Piscopo, M.; Manna, C. Phenol-rich Feijoa sellowiana (Pineapple guava) extracts protect human red blood cells from mercury-induced cellular toxicity. Antioxidants 2019, 8, 220. [Google Scholar] [CrossRef]
- Sychta, K.; Dubas, E.; Yamada, K.; Słomka, A.; Krzewska, M.; Kuta, E. Papain-like cysteine proteases are involved in programmed cell death in plants under heavy metal stress. Environ. Exp. Bot. 2020, 174, 104041. [Google Scholar] [CrossRef]
- Yanık, F.; Aytürk, Ö.; Vardar, F. Programmed cell death evidence in wheat (Triticum aestivum L.) roots induced by aluminum oxide (Al2O3) nanoparticles. Caryologia 2017, 70, 112–119. [Google Scholar] [CrossRef]
- Yao, S.; Huang, W.; Pan, C.; Zhan, J.; He, L.F. Caspase-like proteases regulate aluminum-induced programmed cell death in peanut. Plant Cell. Tissue Organ Cult. 2016, 127, 691–703. [Google Scholar] [CrossRef]
- Hou, S.; Jamieson, P.; He, P. The cloak, dagger, and shield: Proteases in plant–pathogen interactions. Biochem. J. 2018, 475, 2491–2509. [Google Scholar] [CrossRef]
- Chen, Y.L.; Lee, C.Y.; Cheng, K.T.; Chang, W.H.; Huang, R.N.; Nam, H.G.; Chen, Y.R. Quantitative peptidomics study reveals that a wound-induced peptide from PR-1 regulates immune signaling in tomato. Plant Cell 2014, 26, 4135–4148. [Google Scholar] [CrossRef]
- Schaller, A.; Ryan, C.A. Molecular cloning of a tomato leaf cDNA encoding an aspartic protease, a systemic wound response protein. Plant Mol. Biol. 1996, 31, 1073–1077. [Google Scholar] [CrossRef]
- Guevara, M.G.; Oliva, C.R.; Huarte, M.; Daleo, G.R. An aspartic protease with antimicrobial activity is induced after infection and wounding in intercellular fluids of potato tubers. Eur. J. Plant Pathol. 2002, 108, 131–137. [Google Scholar] [CrossRef]
- Guevara, M.G.; Veríssimo, P.; Pires, E.; Faro, C.; Daleo, G.R.; Gabriela Guevara, M.; Veríssimo, P. Potato aspartic proteases: Induction, antimicrobial activity and substrate specificity. J. Plant Pathol. 2004, 233–238. [Google Scholar] [CrossRef]
- Xia, Y.; Ma, Z.; Qiu, M.; Guo, B.; Zhang, Q.; Jiang, H.; Zhang, B.; Lin, Y.; Xuan, M.; Sun, L.; et al. N-glycosylation shields Phytophthora sojae apoplastic effector PsXEG1 from a specific host aspartic protease. Proc. Natl. Acad. Sci. USA 2020, 117, 27685–27693. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Zhu, L.; Song, T.; Wang, Y.; Zhang, Q.; Xia, Y.; Qiu, M.; Lin, Y.; Li, H.; Kong, L.; et al. A paralogous decoy protects Phytophthora sojae apoplastic effector PsXEG1 from a host inhibitor. Science 2017, 355, 710–714. [Google Scholar] [CrossRef]
- Prasad, B.D.; Creissen, G.; Lamb, C.; Chattoo, B.B. Overexpression of rice (Oryza sativa L.) OsCDRI leads to constitutive activation of defense responses in rice and Arabidopsis. Mol. Plant Microbe Interact. 2009, 22, 1635–1644. [Google Scholar] [CrossRef]
- Wang, Y.; Garrido-Oter, R.; Wu, J.; Winkelmüller, T.M.; Agler, M.; Colby, T.; Nobori, T.; Kemen, E.; Tsuda, K. Site-specific cleavage of bacterial MucD by secreted proteases mediates antibacterial resistance in Arabidopsis. Nat. Commun. 2019, 10, 2853. [Google Scholar] [CrossRef]
- Kim, S.G.; Wang, Y.; Lee, K.H.; Park, Z.-Y.; Park, J.; Wu, J.; Kwon, S.J.; Lee, Y.-H.; Agrawal, G.K.; Rakwal, R.; et al. In-depth insight into in vivo apoplastic secretome of rice-Magnaporthe oryzae interaction. J. Proteom. 2013, 78, 58–71. [Google Scholar] [CrossRef]
- Doukhanina, E.V.; Chen, S.; Van Der Zalm, E.; Godzik, A.; Reed, J.; Dickman, M.B. Identification and functional characterization of the BAG protein family in Arabidopsis thaliana. J. Biol. Chem. 2006, 281, 18793–18801. [Google Scholar] [CrossRef]
- Li, Y.; Kabbage, M.; Liu, W.; Dickman, M.B. Aspartyl protease-mediated cleavage of BAG6 is necessary for autophagy and fungal resistance in plants. Plant Cell 2016, 28, 233–247. [Google Scholar] [CrossRef]
- Mott, G.A.; Middleton, M.A.; Desveaux, D.; Guttman, D.S. Peptides and small molecules of the plant-pathogen apoplastic arena. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef]
- Díaz, M.E.; Rocha, G.F.; Kise, F.; Rosso, A.M.; Guevara, M.G.; Parisi, M.G. Antimicrobial activity of an aspartic protease from Salpichroa origanifolia fruits. Lett. Appl. Microbiol. 2018, 67, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Curto, P.; Lufrano, D.; Pinto, C.; Custódio, V.; Gomes, A.C.; Trejo, S.A.; Bakás, L.; Vairo-Cavalli, S.; Faro, C.; Simões, I. Establishing the yeast Kluyveromyces lactis as an expression host for production of the saposin-like domain of the aspartic protease cirsin. Appl. Environ. Microbiol. 2014, 80, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, F.F.; Mendieta, J.R.; Pagano, M.R.; Paggi, R.A.; Daleo, G.R.; Guevara, M.G. The swaposin-like domain of potato aspartic protease (StAsp-PSI) exerts antimicrobial activity on plant and human pathogens. Peptides 2010, 31, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Tian, J. (Jingxin); Yu, H.; Bryksa, B.C.; Dupuis, J.H.; Ou, X.; Qian, Z.; Song, C.; Wang, S.; Yada, R.Y. Insights into the mechanism of membrane fusion induced by the plant defense element, plant-specific insert. J. Biol. Chem. 2020, 295, 14548–14562. [Google Scholar] [CrossRef]
- Figaj, D.; Ambroziak, P.; Przepiora, T.; Skorko-Glonek, J. The Role of Proteases in the Virulence of Plant Pathogenic Bacteria. Int. J. Mol. Sci. 2019, 20, 672. [Google Scholar] [CrossRef]
- Mandujano-González, V.; Villa-Tanaca, L.; Anducho-Reyes, M.A.; Mercado-Flores, Y. Secreted fungal aspartic proteases: A review. Rev. Iberoam. Micol. 2016, 33, 76–82. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Thangavelu, B.; Chun, S.C.; Sathiyabama, M. Proteases from phytopathogenic fungi and their importance in phytopathogenicity. J. Gen. Plant Pathol. 2016, 82, 233–239. [Google Scholar] [CrossRef]
- Espino, J.J.; Gutiérrez-Sánchez, G.; Brito, N.; Shah, P.; Orlando, R.; González, C. The Botrytis cinerea early secretome. Proteomics 2010, 10, 3020–3034. [Google Scholar] [CrossRef]
- Movahedi, S.; Heale, J.B. The roles of aspartic proteinase and endo-pectin lyase enzymes in the primary stages of infection and pathogenesis of various host tissues by different isolates of Botrytis cinerea Pers ex. Pers. Physiol. Mol. Plant Pathol. 1990, 36, 303–324. [Google Scholar] [CrossRef]
- Kelloniemi, J.; Trouvelot, S.; Héloir, M.-C.; Simon, A.; Dalmais, B.; Frettinger, P.; Cimerman, A.; Fermaud, M.; Roudet, J.; Baulande, S.; et al. Analysis of the Molecular Dialogue Between Gray Mold (Botrytis cinerea) and Grapevine (Vitis vinifera) Reveals a Clear Shift in Defense Mechanisms During Berry Ripening. Mol. Plant Microbe Interact. 2015, 28, 1167–1180. [Google Scholar] [CrossRef]
- Haile, Z.M.; Pilati, S.; Sonego, P.; Malacarne, G.; Vrhovsek, U.; Engelen, K.; Tudzynski, P.; Zottini, M.; Baraldi, E.; Moser, C. Molecular analysis of the early interaction between the grapevine flower and Botrytis cinerea reveals that prompt activation of specific host pathways leads to fungus quiescence. Plant. Cell Environ. 2017, 40, 1409–1428. [Google Scholar] [CrossRef] [PubMed]
- Kay, J.; Meijer, H.J.G.; ten Have, A.; van Kan, J.A.L. The aspartic proteinase family of three Phytophthora species. BMC Genom. 2011, 12, 254. [Google Scholar] [CrossRef] [PubMed]
- Meijer, H.J.G.; Mancuso, F.M.; Espadas, G.; Seidl, M.F.; Chiva, C.; Govers, F.; Sabidó, E. Profiling the Secretome and Extracellular Proteome of the Potato Late Blight Pathogen Phytophthora infestans. Mol. Cell. Proteom. 2014, 13, 2101–2113. [Google Scholar] [CrossRef] [PubMed]
- McGowan, J.; O’Hanlon, R.; Owens, R.A.; Fitzpatrick, D.A. Comparative Genomic and Proteomic Analyses of Three Widespread Phytophthora Species: Phytophthora chlamydospora, Phytophthora gonapodyides and Phytophthora pseudosyringae. Microorganisms 2020, 8, 653. [Google Scholar] [CrossRef] [PubMed]
- Schoina, C.; Verbeek-de Kruif, N.; Govers, F.; Bouwmeester, K. Clade 5 aspartic proteases of Phytophthora infestans are virulence factors implied in RXLR effector cleavage. Eur. J. Plant Pathol. 2019, 154, 17–29. [Google Scholar] [CrossRef]
- Guo, B.; Wang, H.; Yang, B.; Jiang, W.; Jing, M.; Li, H.; Xia, Y.; Xu, Y.; Hu, Q.; Wang, F.; et al. Phytophthora sojae Effector PsAvh240 Inhibits Host Aspartic Protease Secretion to Promote Infection. Mol. Plant 2019, 12, 552–564. [Google Scholar] [CrossRef]
- Sun, S.; Lui, Q.; Han, L.; Ma, Q.; He, S.; Li, X.; Zhang, H.; Zhang, J.; Liu, X.; Wang, L. Identification and Characterization of Fusarium proliferatum, a New Species of Fungi that Cause Fungal Keratitis. Sci. Rep. 2018, 8, 4859. [Google Scholar] [CrossRef]
- Li, T.; Wu, Q.; Wang, Y.; John, A.; Qu, H.; Gong, L.; Duan, X.; Zhu, H.; Yun, Z.; Jiang, Y. Application of Proteomics for the Investigation of the Effect of Initial pH on Pathogenic Mechanisms of Fusarium proliferatum on Banana Fruit. Front. Microbiol. 2017, 8, 2327. [Google Scholar] [CrossRef]
- Krishnan, P.; Ma, X.; McDonald, B.A.; Brunner, P.C. Widespread signatures of selection for secreted peptidases in a fungal plant pathogen. BMC Evol. Biol. 2018, 18, 7. [Google Scholar] [CrossRef]
- Poussereau, N.; Gente, S.; Rascle, C.; Billon-Grand, G.; Fèvre, M. aspS encoding an unusual aspartyl protease from Sclerotinia sclerotiorum is expressed during phytopathogenesis. FEMS Microbiol. Lett. 2001, 194, 27–32. [Google Scholar] [CrossRef]
- Urbanek, H.; Yirdaw, G. Hydrolytic ability of acid protease of Fusarium culmorum and its possible role in phytopathogenesis. Acta Microbiol. Pol. 1984, 33, 131–136. [Google Scholar] [PubMed]
- Choi, G.H.; Pawlyk, D.M.; Rae, B.; Shapira, R.; Nuss, D.L. Molecular analysis and overexpression of the gene encoding endothiapepsin, an aspartic protease from Cryphonectria parasitica. Gene 1993, 125, 135–141. [Google Scholar] [CrossRef]
- Mandujano-González, V.; Arana-Cuenca, A.; Anducho-Reyes, M.Á.; Téllez-Jurado, A.; González-Becerra, A.E.; Mercado-Flores, Y. Biochemical study of the extracellular aspartyl protease Eap1 from the phytopathogen fungus Sporisorium reilianum. Protein Expr. Purif. 2013, 92, 214–222. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mandujano-González, V.; Téllez-Jurado, A.; Anducho-Reyes, M.A.; Arana-Cuenca, A.; Mercado-Flores, Y. Purification and characterization of the extracellular aspartyl protease APSm1 from the phytopathogen fungus Stenocarpella maydis. Protein Expr. Purif. 2016, 117, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.; Cheong, H.; Choi, O.; Kim, J.K.; Kang, Y.; Kim, J.; Lee, S.; Koh, S.; Moon, J.S.; Hwang, I. An HrpB-dependent but type III-independent extracellular aspartic protease is a virulence factor of Ralstonia solanacearum. Mol. Plant Pathol. 2011, 12, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Jashni, M.K.; van der Burgt, A.; Battaglia, E.; Mehrabi, R.; Collemare, J.; de Wit, P.J.G.M. Transcriptome and proteome analyses of proteases in biotroph fungal pathogen Cladosporium fulvum. J. Plant Pathol. 2020, 102, 377–386. [Google Scholar] [CrossRef]
- Levin, E.; Kishore, A.; Ballester, A.R.; Raphael, G.; Feigenberg, O.; Liu, Y.; Norelli, J.; Gonzalez-Candelas, L.; Wisniewski, M.; Droby, S. Identification of pathogenicity-related genes and the role of a subtilisin-related peptidase S8 (PePRT) in authophagy and virulence of Penicillium expansum on apples. Postharvest Biol. Technol. 2019, 149, 209–220. [Google Scholar] [CrossRef]
- Gregori, R.; Guidarelli, M.; Mari, M. Preliminary studies on partial reduction of Colletotrichum acutatum infection by proteinase inhibitors extracted from apple skin. Physiol. Mol. Plant Pathol. 2010, 74, 303–308. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).