
Structural Aspects of Potential Endocrine-Disrupting Activity of Stereoisomers for a Common Pesticide Permethrin against Androgen Receptor


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Protein Preparation
2.2. Ligand Preparation
2.3. Induced Fit Docking
2.4. Binding Affinity Calculations
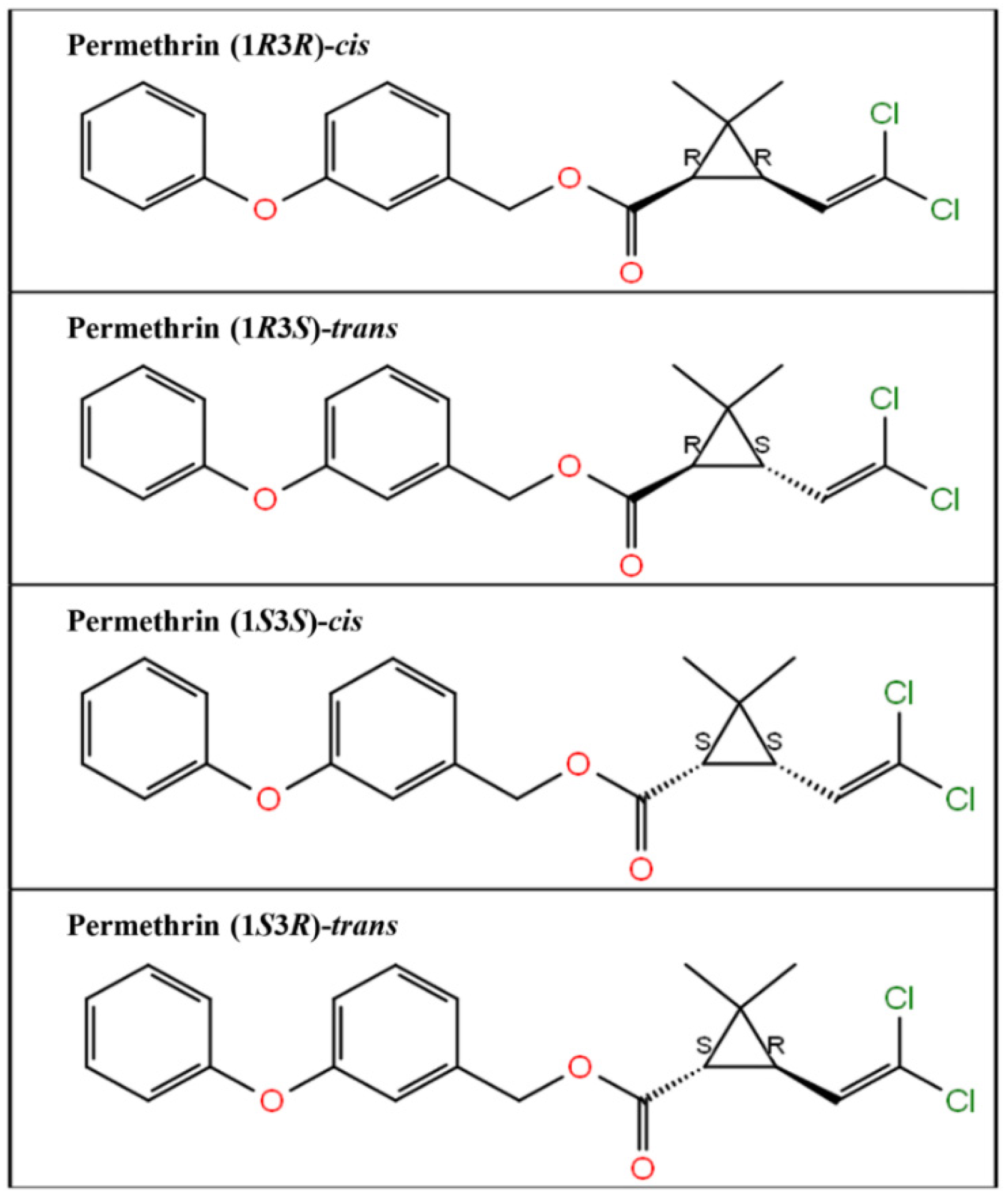
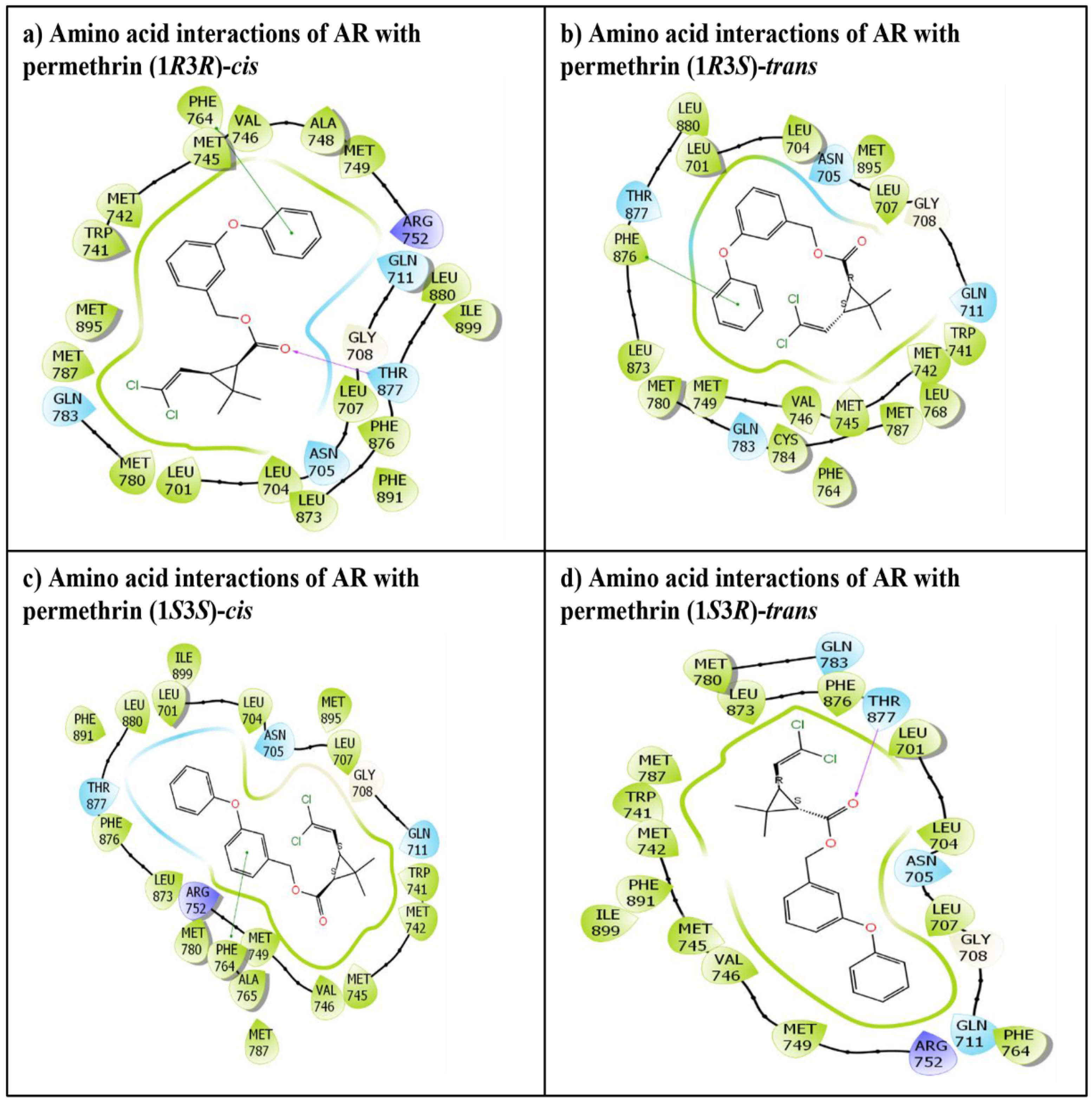
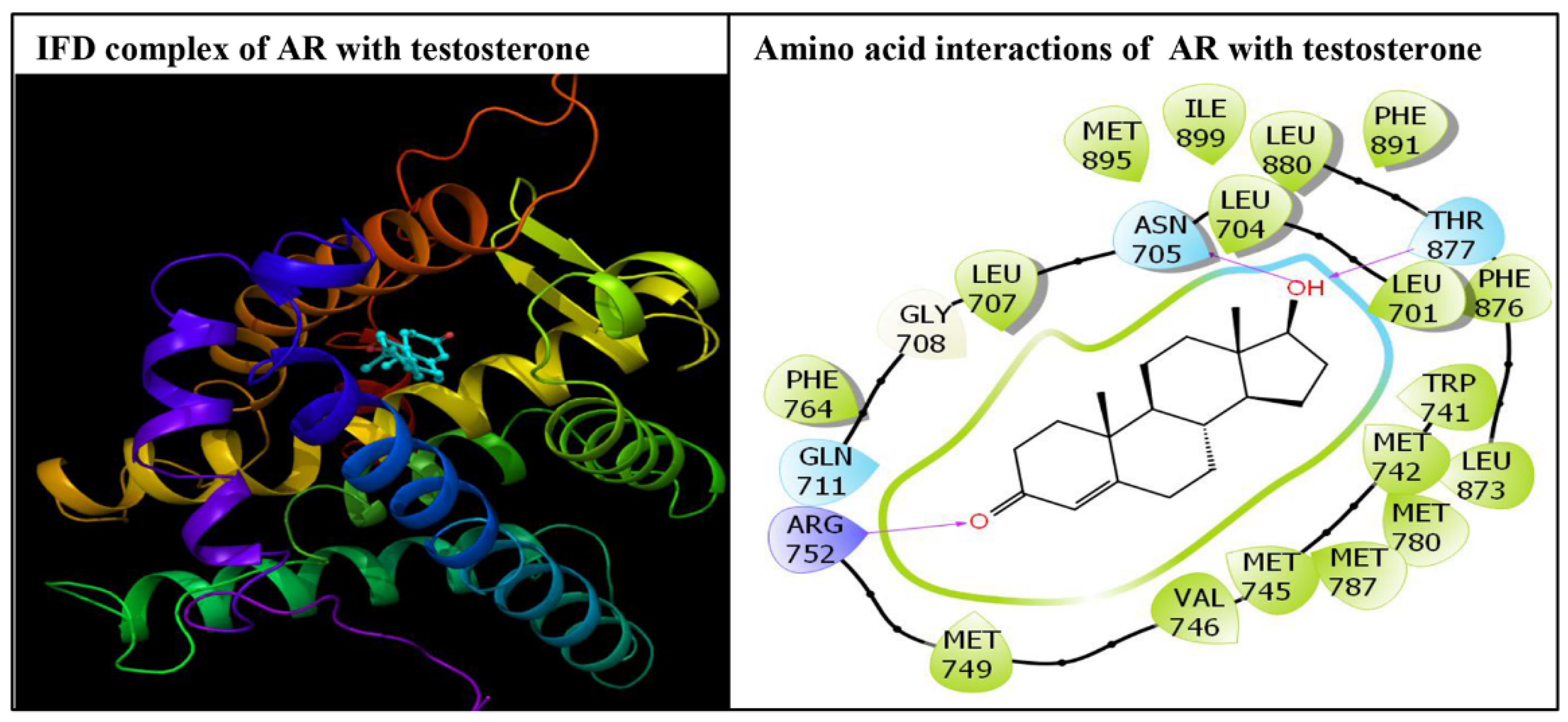
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Annamalai, J.; Namasivayam, V. Endocrine disrupting chemicals in the atmosphere: Their effects on humans and wildlife. Environ. Int. 2015, 76, 78–97. [Google Scholar] [CrossRef] [PubMed]
- Lauretta, R.; Sansone, A.; Sansone, M.; Romanelli, F.; Appetecchia, M. Endocrine Disrupting Chemicals: Effects on Endocrine Glands. Front. Endocrinol. 2019, 10, 178. [Google Scholar] [CrossRef]
- Ye, X.; Liu, J. Effects of pyrethroid insecticides on hypothalamic-pituitary-gonadal axis: A reproductive health perspective. Environ. Pollut. 2019, 245, 90–599. [Google Scholar] [CrossRef]
- Zhu, Q.; Yang, Y.; Zhong, Y.; Lao, Z.; O’Neill, P.; Hong, D.; Zhang, K.; Zhao, S.; Suqing, Z. Synthesis, insecticidal activity, resistance, photodegradation and toxicity of pyrethroids (A review). Chemosphere 2020, 254, 126779. [Google Scholar] [CrossRef]
- Bian, Q.; Xu, L.C.; Wang, S.L.; Xia, Y.K.; Tan, L.F.; Chen, J.F.; Song, L.; Chang, H.C.; Wang, X.R. Study on the relation between occupational fenvalerate exposure and spermatozoa DNA damage of pesticide factory workers. Occup. Environ. Med. 2004, 61, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Pine, M.D.; Hiney, J.K.; Lee, B.; Dees, W.L. The pyrethroid pesticide esfenvalerate suppresses the afternoon rise of luteinizing hormone and delays puberty in female rats. Environ. Health Perspect 2008, 116, 1243–1247. [Google Scholar] [CrossRef]
- Palmquist, K.; Salatas, J.; Fairbrother, A. Pyrethroid insecticides: Use, environmental fate, and ecotoxicology. In Insecticides—Advances in Integrated Pest Management; Perveen, F., Ed.; InTech Open: London, UK, 2012; pp. 251–278. [Google Scholar]
- Fenner, K.; Canonica, S.; Wackett, L.P.; Elsner, M. Evaluating pesticide degradation in the environment: Blind spots and emerging opportunities. Science 2013, 341, 752–758. [Google Scholar] [CrossRef] [PubMed]
- IMARC Group. Pyrethroids Market: Global Industry Trends, Share, Size, Growth, Opportunity and Forecast 2020–2025. 2020. Available online: https://www.imarcgroup.com/pyrethroids-market (accessed on 20 December 2020).
- Marketresearch. Global Pyrethroid Insecticide Market- Growth, Trends and Forecasts (2017–2022). 2020. Available online: www.marketresearch.com/Mordor-Intelligence-LLP-v4018/Global-Pyrethroid-Insecticide-Growth-Trends-11110002/ (accessed on 20 December 2020).
- ReportLinker. New York, Sept. 25, 2020 (GLOBE NEWSWIRE). Global Pyrethroids Industry. 2020. Available online: https://www.reportlinker.com/p05959828/?utm_source=GNW (accessed on 20 December 2020).
- UH. Pesticide Properties Database; University of Hertfordshire. 2011. Available online: http://sitem.herts.ac.uk/aeru/footprint/index2.htm (accessed on 20 December 2020).
- Corcellas, C.; Andreu, A.; Manez, M.; Sergio, F.; Hiraldo, F.; Eljarrat, E.; Barcelo, D. Pyrethroid insecticides in wild bird eggs from a World Heritage Listed Park: A case study in Donana National Park (Spain). Environ. Pollut. 2017, 228, 321–330. [Google Scholar] [CrossRef]
- Barr, D.B.; Olsson, A.O.; Wong, L.Y.; Udunka, S.; Baker, S.E.; Whitehead, R.D.; Magsumbol, M.S.; Williams, B.L.; Needham, L.L. Urinary concentrations of metabolites of pyrethroid insecticides in the general U.S. Population: National health and nutrition examination survey 1999–2002. Environ. Health Perspect 2010, 118, 742–748. [Google Scholar] [CrossRef]
- Naeher, L.P.; Tulve, N.S.; Egeghy, P.P.; Barr, D.B.; Adetona, O.; Fortmann, R.C.; Needham, L.L.; Bozeman, E.; Hilliard, A.; Sheldon, L.S. Organophosphorus and pyrethroid insecticide urinary metabolite concentrations in young children living in a southeastern United States city. Sci. Total Environ. 2010, 408, 1145–1153. [Google Scholar] [CrossRef]
- Trunnelle, K.J.; Bennett, D.H.; Tulve, N.S.; Clifton, M.S.; Davis, M.D.; Calafat, A.M.; Moran, R.; Tancredi, D.J.; Hertz-Picciotto, I. Urinary pyrethroid and chlorpyrifos metabolite concentrations in northern California families and their relationship to indoor residential insecticide levels, part of the study of use of products and exposure related behavior (superb). Environ. Sci. Technol. 2014, 48, 1931–1939. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Pan, W.; Zhao, S.; Zhao, Y.; Zhu, Y.; Liu, J.; Liu, W. Relationships of pyrethroid exposure with gonadotropin levels and pubertal development in Chinese boys. Environ. Sci. Technol. 2017, 51, 6379–6386. [Google Scholar] [CrossRef]
- Sun, H.; Xu, X.L.; Xu, L.C.; Song, L.; Hong, X.; Chen, J.F.; Cui, L.B.; Wang, X.R. Antiandrogenic activity of pyrethroid pesticides and their metabolite in reporter gene assay. Chemosphere 2007, 66, 474–479. [Google Scholar] [CrossRef]
- Li, Y.F.; Pan, C.; Hu, J.X.; Li, J.; Xu, L.C. Effects of cypermethrin on male reproductive system in adult rats. Biomed. Environ. Sci. 2013, 26, 201–208. [Google Scholar]
- Pascotto, V.M.; Guerra, M.T.; Franci, J.A.; de Camargo, J.L.; Kempinas, W.G.; Franchi, C.A. Effects of a mixture of pesticides on the adult female reproductive system of sprague-dawley, wistar, and lewis rats. J. Toxicol. Environ. Health Part A 2015, 78, 602–616. [Google Scholar] [CrossRef]
- Ye, X.; Li, F.; Zhang, J.; Ma, H.; Ji, D.; Huang, X.; Curry, T.E., Jr.; Liu, W.; Liu, J. Pyrethroid insecticide cypermethrin accelerates pubertal onset in male mice via disrupting hypothalamic-pituitary-gonadal axis. Environ. Sci. Technol. 2017, 51, 10212–10221. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Y.; Ito, Y.; Yamanoshita, O.; Yanagiba, Y.; Kobayashi, M.; Taya, K.; Li, C.M.; Okamura, A.; Miyata, M.; Ueyama, J.; et al. Permethrin may disrupt testosterone biosynthesis via mitochondrial membrane damage of leydig cells in adult male mouse. Endocrinology 2007, 148, 3941–3949. [Google Scholar] [CrossRef]
- Wolansky, M.J.; Harrill, J.A. Neurobehavioral toxicology of pyrethroid insecticides in adult animals: A critical review. Neurotoxicol. Teratol. 2008, 30, 55–78. [Google Scholar] [CrossRef]
- Khaki, A.; Khaki, A.A.; Rajabzadeh, A. The effects of Permethrin and antioxidant properties of Allium cepa (onion) on testicles parameters of male rats. Toxin. Rev. 2017, 36, 1–6. [Google Scholar] [CrossRef]
- Ujihara, K. The history of extensive structural modifications of pyrethroids. J. Pestic Sci. 2019, 44, 215–224. [Google Scholar] [CrossRef]
- Diao, J.L.; Xu, P.; Liu, D.H.; Zhou, Z.Q. Enantiomer-specific toxicity and bioaccumulation of alpha-cypermethrin to earthworm Eisenia fetida. J. Hazard. Mater. 2011, 192, 1072–1078. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Gan, J.; Lee, S.; Werner, I. Isomer selectivity in aquatic toxicity and biodegradation of bifenthrin and permethrin. Environ. Toxicol. Chem. 2005, 24, 1861–1866. [Google Scholar] [CrossRef] [PubMed]
- Nillos, M.G.; Chajkowski, S.; Rimoldi, J.M.; Gan, J.; Lavado, R.; Schlenk, D. Stereoselective biotransformation of permethrin to estrogenic metabolites in fish. Chem. Res. Toxicol. 2010, 23, 1568–1575. [Google Scholar] [CrossRef] [PubMed]
- Dornetshuber, J.; Bicker, W.; Lämmerhofer, M.; Lindner, W.; Karwan, A.; Bursch, W. Impact of stereochemistry on biological effects of permethrin: Induction of apoptosis in human hepatoma cells (HCC-1.2) and primary rat hepatocyte cultures. BMC Pharmacol. 2007, 7, A65. [Google Scholar] [CrossRef]
- Leicht, W.; Fuchs, R.; Londershausen, M. Stability and biological activity of cyfluthrin isomers. Pestic. Sci. 1996, 48, 325–332. [Google Scholar] [CrossRef]
- Naumann, K. Synthetic Pyrethroids Insecticides: Structures and Properties; Springer: Berlin, Germany, 1990. [Google Scholar]
- Pihlajamaa, P.; Sahu, B.; Janne, O.A. Determinants of receptor- and tissue-specific actions in androgen signaling. Endocr. Rev. 2015, 36, 357–384. [Google Scholar] [CrossRef]
- Beg, M.A.; Sheikh, I.A. Endocrine disruption: Structural interactions of androgen receptor against Di(2-ethylhexyl) phthalate and its metabolites. Toxics 2020, 8, 115. [Google Scholar] [CrossRef]
- Sheikh, I.A.; Beg, M.A. Structural binding interactions of tetrabromobisphenol A with sex steroid nuclear receptors and sex hormone-binding globulin. J. Appl. Toxicol. 2020, 40, 832–842. [Google Scholar] [CrossRef]
- Sheikh, I.A. Endocrine-disrupting potential of polybrominated diphenyl ethers (PBDEs) on androgen receptor signaling: A structural insight. Struct. Chem. 2020. [Google Scholar] [CrossRef]
- Sheikh, I.A. Stereoselectivity and potential endocrine disrupting activity of Bis-(2-ethylhexyl) phthalate (DEHP) against human progesterone receptor: A computational perspective. J. Appl. Toxicol. 2016, 36, 741–747. [Google Scholar] [CrossRef]
- Romero, A.; Ares, I.; Ramos, E.; Castellano, V.; Martinez, M.; Martinez- Larranaga, M.R.; Anadon, A.; Martinez, M.A. Evidence for dose-additive effects of a type II pyrethroid mixture. In vitro assessment. Environ. Res. 2015, 138, 58–66. [Google Scholar] [CrossRef]
- Koureas, M.; Tsakalof, A.; Tsatsakis, A.; Hadjichristodoulou, C. Systematic review of biomonitoring studies to determine the association between exposure to organophosphorus and pyrethroid insecticides and human health outcomes. Toxicol. Lett. 2012, 210, 155–168. [Google Scholar] [CrossRef]
- Morgan, M.K. Children’s exposures to pyrethroid insecticides at home: A review of data collected in published exposure measurement studies conducted in the United States. Int. J. Environ. Res. Publ. Health 2012, 9, 2964–2985. [Google Scholar] [CrossRef] [PubMed]
- Swan, S.H.; Elkin, E.P.; Fenster, L. The question of declining sperm density revisited: An analysis of 101 studies published 1934–1996. Environ. Health Perspect 2000, 108, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Travison, T.G.; Araujo, A.B.; O’Donnell, A.B.; Kupelian, V.; McKinlay, J.B. A population-level decline in serum testosterone levels in American men. J. Clin. Endocrinol. Metab. 2007, 92, 196–202. [Google Scholar] [CrossRef]
- Meeker, J.D.; Barr, D.B.; Hauser, R. Pyrethroid insecticide metabolites are associated with serum hormone levels in adult men. Reprod. Toxicol. 2009, 27, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Burns, C.J.; Pastoor, T.P. Pyrethroid epidemiology: A quality-based review. Crit. Rev. Toxicol. 2018, 48, 297–311. [Google Scholar] [CrossRef]
- Tang, W.; Wang, D.; Wang, J.; Wu, Z.; Li, L.; Huang, M.; Xu, S.; Yan, D. Pyrethroid pesticide residues in the global environment: An overview. Chemosphere 2018, 191, 990–1007. [Google Scholar] [CrossRef] [PubMed]
- Toynton, K.; Luukinen, B.; Buhl, K.; Stone, D. Permethrin Technical Fact Sheet; National Pesticide Information Center, Oregon State University Extension Services: Baker City, OR, USA, 2009. [Google Scholar]
- Osaka, A.; Ueyama, J.; Kondo, T.; Nomura, H.; Sugiura, Y.; Saito, I.; Nakane, K.; Takaishi, A.; Ogi, H.; Wakusawa, S.; et al. Exposure characterization of three major insecticide lines in urine of young children in Japan-neonicotinoids, organophosphates, and pyrethroids. Environ Res. 2016, 147, 89–96. [Google Scholar] [CrossRef]
- Glorennec, P.; Serrano, T.; Fravallo, M.; Warembourg, C.; Monfort, C.; Cordier, S.; Viel, J.; Le Gléau, F.; Le Bot, B.; Chevrier, C. Determinants of children’s exposure to pyrethroid insecticides in western France. Environ. Int. 2017, 104, 76–82. [Google Scholar] [CrossRef] [PubMed]
- CDC. Fourth National Report on Human Exposure to Environmental Chemicals Updated Tables; United States Department of Health and Human Services; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2019; Volume One and Two, pp. 223–232. Available online: https://www.cdc.gov/exposurereport/pdf/FourthReport_UpdatedTables_Volume1_Jan2019-508.pdf.gov (accessed on 20 December 2020).
- ATSDR. Toxicological Profile for Pyrethrins and Pyrethroids; Agency for Toxic Substances and Disease Registry, Division of Toxicology: Atlanta, GA, USA, 2003. Available online: https://www.atsdr.cdc.gov/ToxProfiles/tp155-c1-b.pdf (accessed on 20 December 2020).
- EPA. Prevention, Pesticides and Toxic Substances: Permethrin Facts; EPA 738-F-09-001; The United States Environmental Protection Agency: Washington, DC, USA, 2009. Available online: https://www3.epa.gov/pesticides/chem_search/reg_actions/reregistration/fs_PC-109701_1-Aug-09.pdf (accessed on 20 December 2020).
- Tu, W.; Xu, C.; Jin, Y.; Lu, B.; Lin, C.; Wu, Y.; Liu, W. Permethrin is a potential thyroid-disrupting chemical: In vivo and in silico envidence. Aquat. Toxicol. 2016, 175, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Eil, C.; Nisula, B.C. The binding properties of pyrethroids to human skin fibroblast androgen receptors and to sex hormone binding globulin. J. Steroid. Biochem. 1990, 35, 409–414. [Google Scholar] [CrossRef]
- Xu, L.C.; Liu, L.; Ren, X.M.; Zhang, M.R.; Cong, N.; Xu, A.Q.; Shao, J.H. Evaluation of androgen receptor transcriptional activities of some pesticides in vitro. Toxicology 2007, 243, 59–65. [Google Scholar] [CrossRef]
- Han, Y.; Xia, Y.K.; Han, J.Y.; Zhou, J.P.; Wang, S.L.; Zhu, P.F.; Zhao, R.C.; Jin, N.Z.; Song, L.; Wang, X.R. The relationship of 3-PBA pyrethroids metabolite and male reproductive hormones among non-occupational exposure males. Chemosphere 2008, 72, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Green, M.P.; Harvey, A.J.; Finger, B.J.; Tarulli, G.A. Endocrine disrupting chemicals: Impacts on human fertility and fecundity during the peri-conception period. Environ. Res. 2020, 194, 110694. [Google Scholar] [CrossRef]
- Mostafa, H.E.; Abd El-Baset, S.A.; Kattaia, A.A.; Zidan, R.A.; Al Sadek, M.M. Efficacy of naringenin against perme-thrin-induced testicular toxicity in rats. Int. J. Exp. Pathol. 2016, 97, 37–49. [Google Scholar] [CrossRef]
- Dohlman, T.M.; Jahnke, M.M.; West, J.K.; Phillips, P.E.; Gunn, P.J. Effects of label-dose permethrin administration in yearling beef cattle: I. Reproductive function and embryo quality of superovulated heifers. Theriogenology 2016, 85, 1528–1533. [Google Scholar] [CrossRef]
- Issam, C.; Zohra, H.; Monia, Z.; Hassen, B.C. Effects of dermal sub-chronic exposure of pubescent male rats to perme-thrin (prmt) on the histological structures of genital tract, testosterone and lipoperoxidation. Exp. Toxicol. Pathol. 2011, 63, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Liu, J.; Wang, L.; Chen, R.; Zhou, C.; Yang, Y.; Liu, W.; Fu, Z. Permethrin exposure during puberty has the poten-tial to enantioselectively induce reproductive toxicity in mice. Environ. Int. 2012, 42, 144–151. [Google Scholar] [CrossRef]
- Hu, F.; Li, L.; Wang, C.; Zhang, Q.; Zhang, X.; Zhao, M. Enantioselective induction of oxidative stress by permethrin in rat adrenal pheochromocytoma (PC12) cells. Environ. Toxicol. Chem. 2010, 29, 683–690. [Google Scholar] [CrossRef]
- Willemin, M.E.; Desmots, S.; Le Grand, R.; Lestremau, F.; Zeman, F.A.; Leclerc, E.; Moesch, C.; Brochot, C. PBPK modeling of the cis- and trans-permethrin isomers and their major urinary metabolites in rats. Toxicol. Appl. Pharmacol. 2016, 294, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Lavado, R.; Li, J.; Rimoldi, J.M.; Schlenk, D. Evaluation of stereoselective biotransformation of permethrin in human liver microsomes: Contributions of cytochrome P450 monooxygenase to the formation of estrogenic metabolites. Toxicol. Lett. 2014, 226, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Chen, R.; Sun, L.; Wang, W.; Zhou, L.; Liu, W.; Fu, Z. Enantioselective induction of estrogen-responsive gene ex-pression by permethrin enanatiomers in embryo-larval zebrafish. Chemosphere 2009, 74, 1238–1244. [Google Scholar] [CrossRef] [PubMed]
- Chrustek, A.; Hołyńska-Iwan, I.; Dziembowska, I.; Bogusiewicz, J.; Wróblewski, M.; Cwynar, A.; Olszewska-Słonina, D. Current Research on the Safety of Pyrethroids Used as Insecticides. Medicina 2018, 54, 61. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Ligand | Number of Interacting AR Residues | Percentage of Interacting Residues Common with Native Ligand (%) | IFD Score | Docking Score (Kcal/mol) | Glide Score (Kcal/mol) | MMGB-SA (Kcal/mol) |
|---|---|---|---|---|---|---|
| Permethrin (1R3R)-cis | 24 | 100 | −575.06 | −10.87 | −10.87 | −145.62 |
| Permethrin (1R3S)-trans | 22 | 86 | −575.58 | −10.57 | −10.57 | −144.54 |
| Permethrin (1S3S)-cis | 23 | 100 | −574.66 | −10.37 | −10.37 | −134.02 |
| Permethrin (1S3R)-trans | 21 | 86 | −575.70 | −10.42 | −10.42 | −144.76 |
| Testosterone | 22 | 100 | −577.54 | −12.85 | −12.87 | −152.82 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheikh, I.A.; Beg, M.A. Structural Aspects of Potential Endocrine-Disrupting Activity of Stereoisomers for a Common Pesticide Permethrin against Androgen Receptor. Biology 2021, 10, 143. https://doi.org/10.3390/biology10020143
Sheikh IA, Beg MA. Structural Aspects of Potential Endocrine-Disrupting Activity of Stereoisomers for a Common Pesticide Permethrin against Androgen Receptor. Biology. 2021; 10(2):143. https://doi.org/10.3390/biology10020143
Chicago/Turabian StyleSheikh, Ishfaq Ahmad, and Mohd Amin Beg. 2021. "Structural Aspects of Potential Endocrine-Disrupting Activity of Stereoisomers for a Common Pesticide Permethrin against Androgen Receptor" Biology 10, no. 2: 143. https://doi.org/10.3390/biology10020143
APA StyleSheikh, I. A., & Beg, M. A. (2021). Structural Aspects of Potential Endocrine-Disrupting Activity of Stereoisomers for a Common Pesticide Permethrin against Androgen Receptor. Biology, 10(2), 143. https://doi.org/10.3390/biology10020143