
Influence of Hyperproteinemia on Insect Innate Immune Function of the Circulatory System in Bombyx mori

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Model
2.2. Fat Body Collection and Transcriptome Sequencing
2.3. Transcriptome Assembly, Annotation, and Differential Analysis
2.4. Real-Time Polymerase Chain Reaction Analysis
2.5. Determination of Phenoloxidase (PO) Activity
2.6. Melanization Measurement
2.7. Hemocyte Counting
2.8. Antibacterial Activity
2.9. Data Analysis
3. Results
3.1. High PPC Induces Hemocyte Phagocytosis
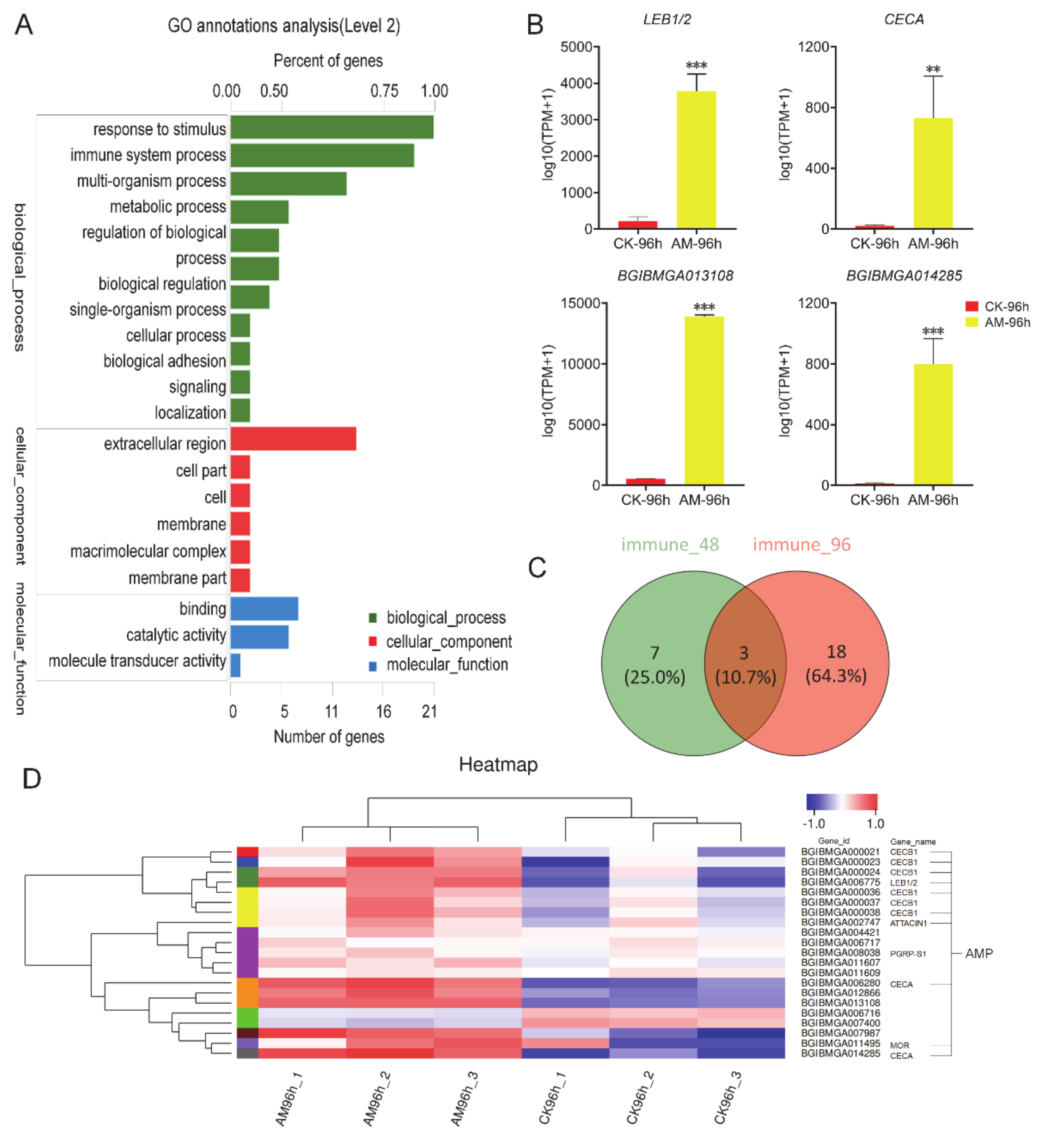
3.2. High PPC Affects the Expression of Innate Immune-Related Genes
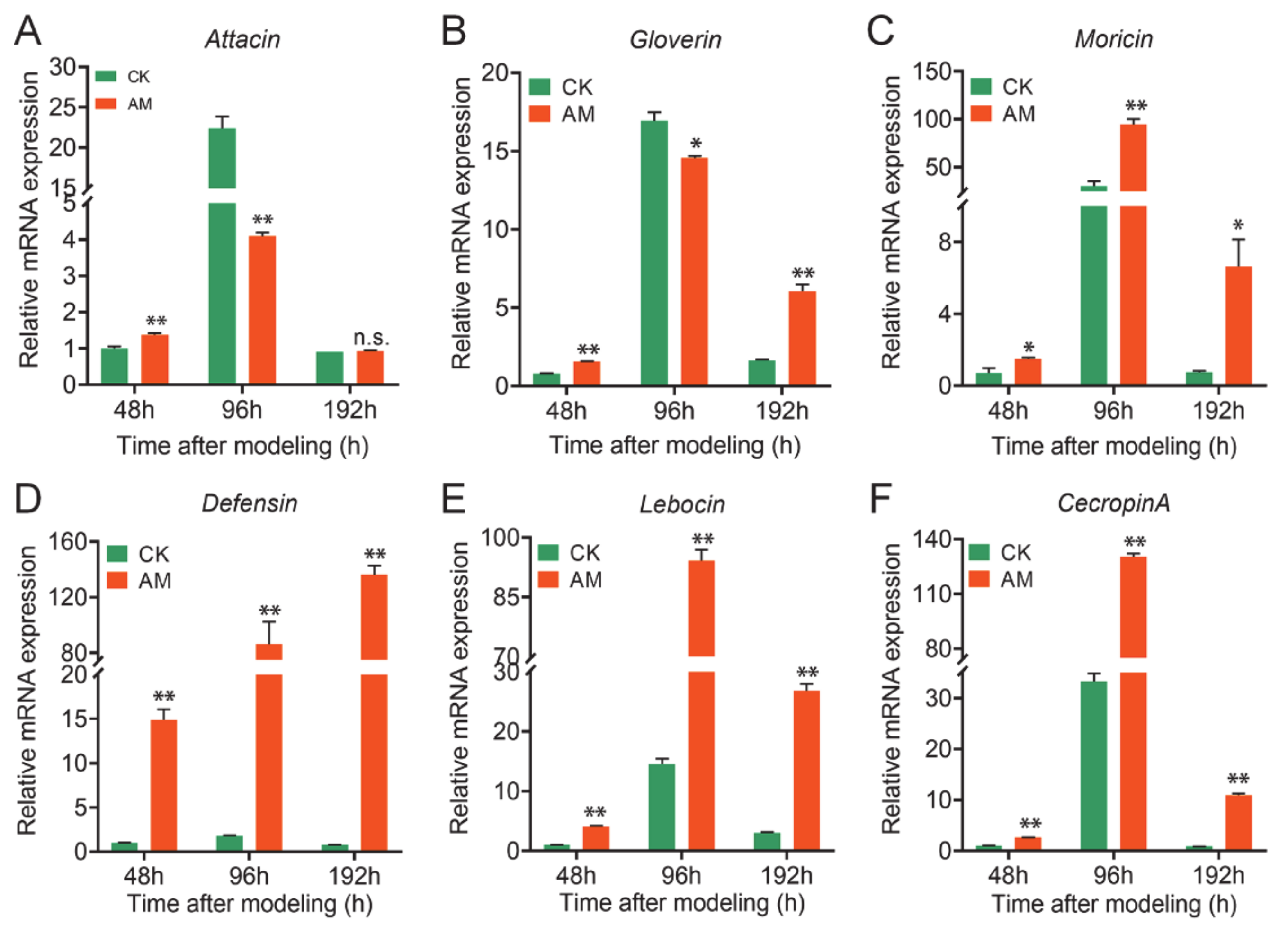
3.3. High PPC Affects Hemolymph Antibacterial Activity
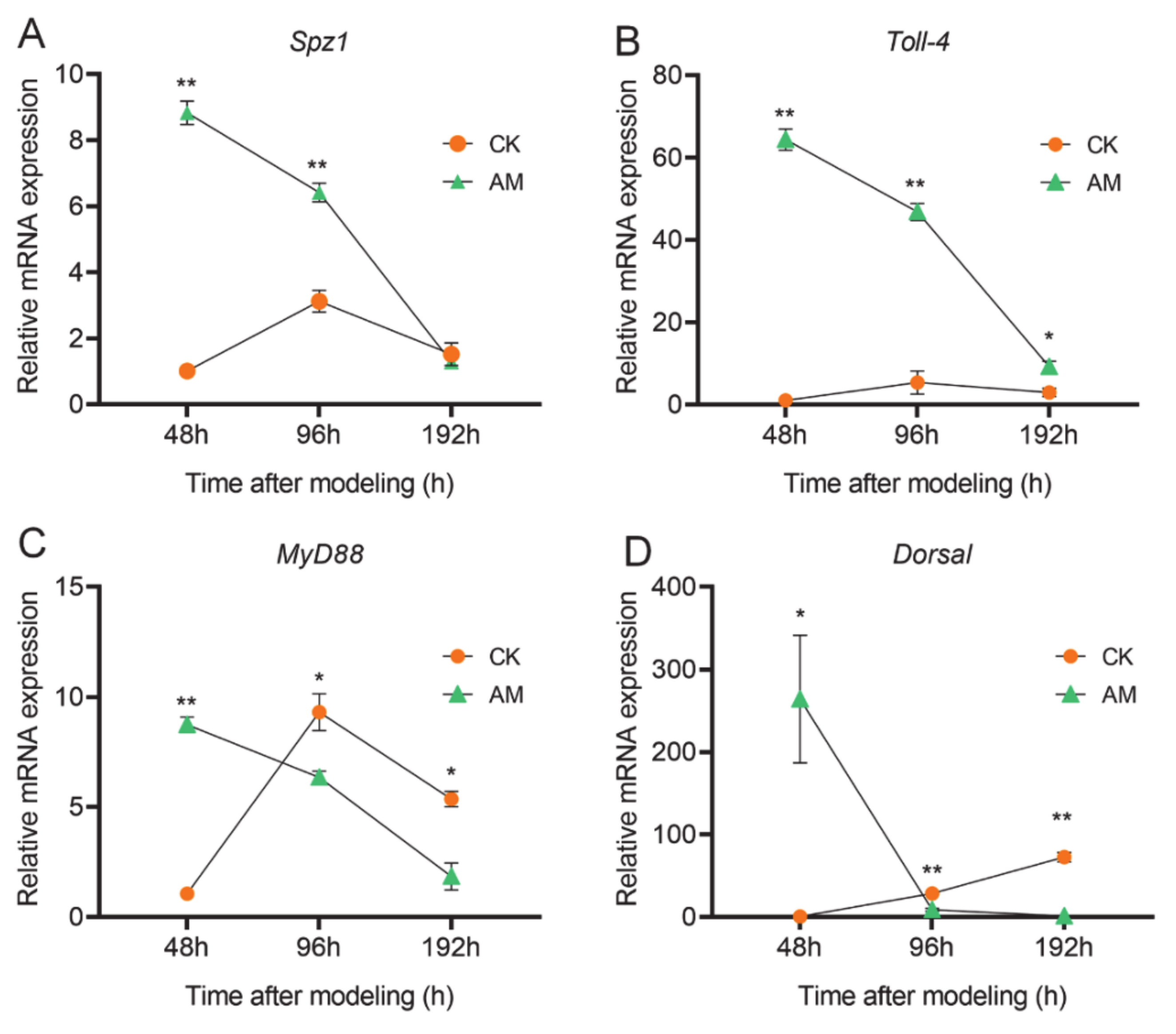
3.4. AMP Expression is Regulated by NF-κB Signaling
3.5. High PPC Affects Hemolymph Melanization
4. Discussion
4.1. High PPC Regulates Innate Immunity via the NF-κB Signaling Pathway
4.2. High PPC Inhibits Hemolymph Melanization in Silkworm
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singer, C.E.; Neagoe, D.; Coşoveanu, S.; Puiu, I.; Oancea, G. Severe Toxocariasis in Children-Diagnostic Difficulties. Curr. Health Sci. J. 2016, 42, 413–416. [Google Scholar] [PubMed]
- Meléndez-Lazo, A.; Ordeix, L.; Planellas, M.; Pastor, J.; Solano-Gallego, L. Clinicopathological findings in sick dogs naturally infected with Leishmania infantum: Comparison of five different clinical classification systems. Res. Veter Sci. 2018, 117, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Perondi, F.; Brovida, C.; Ceccherini, G.; Guidi, G.; Lippi, I. Double filtration plasmapheresis in the treatment of hyperproteinemia in dogs affected by Leishmania infantum. J. Veter Sci. 2018, 19, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Almenfi, H.F.; Almehdewi, A.M.; Hamza, M.S.; Bhat, M.S.; Vijayashankar, N.P. Laboratory Features of Newly Diagnosed Multiple Myeloma Patients. Cureus 2019, 11, e4716. [Google Scholar] [CrossRef] [PubMed]
- Kluck, G.E.G.; Wendt, C.H.C.; Imperio, G.E.D.; Araujo, M.F.C.; Atella, T.C.; Da Rocha, I.; Miranda, K.R.; Atella, G.C. Plasmodium Infection Induces Dyslipidemia and a Hepatic Lipogenic State in the Host through the Inhibition of the AMPK-ACC Pathway. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, Y.; Iizuka, H.; Kanemitsu, H.; Fujino, Y.; Nakashima, K.; Uchida, K.; Ohno, K.; Nakayama, H.; Tsujimoto, H. Myeloma-related disorder with leukaemic progression in a cat. J. Feline Med. Surg. 2010, 12, 982–987. [Google Scholar] [CrossRef]
- Salvesen, H.A. Hyperproteinemia in a Case of Nephrosis.1. J. Intern. Med. 2010, 65, 152–159. [Google Scholar] [CrossRef]
- Kitazawa, A.; Koda, R.; Yoshino, A.; Ueda, Y.; Takeda, T. An IgA1-lambda-type monoclonal immunoglobulin deposition disease associated with membranous features in a patient with IgG4-related kidney disease: A case report. BMC Nephrol. 2018, 19, 1–7. [Google Scholar] [CrossRef]
- Matoušek, V.; Herold, I.; Holanová, L.; Balik, M. A Rare Case of Severe Metabolic Alkalosis with Unusual Hyperproteinemia Treated with Continuous Renal Replacement Therapy and Regional Citrate Anticoagulation. Case Rep. Nephrol. Dial. 2018, 8, 138–146. [Google Scholar] [CrossRef]
- Oliveira, V.D.C.; Junior, A.A.V.M.; Cavalcanti, M.C.D.H.; Madeira, M.D.F.; Ferreira, L.C.; Figueiredo, F.B.; De Campos, M.P.; Nadal, N.V.; Almosny, N.R.P.; Menezes, R. First description of parasite load and clinicopathological and anatomopathological changes in a dog naturally coinfected with Dioctophyme renale and Leishmania infantum in Brazil. Veter Parasitol. Reg. Stud. Rep. 2019, 18, 100351. [Google Scholar] [CrossRef]
- Bergstedt, J.; Lingen, C. Cirrhosis of the Liver in Two Children with Hyperproteinaemia. Acta Paediatr. 1957, 46, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, C.; Malbon, A.; Riond, B.; Schoster, A. κ Light-chain monoclonal gammopathy and cast nephropathy in a horse with multiple myeloma. J. Am. Veter Med. Assoc. 2018, 253, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Adeyeba, O.A. Hyperproteinaemia in dracunculiasis. Int. J. Zoonoses 1985, 12, 150–151. [Google Scholar] [PubMed]
- Wang, F.; Butler, T.; Rabbani, G.; Jones, P.K. The Acidosis of Cholera. New Engl. J. Med. 1986, 315, 1591–1595. [Google Scholar] [CrossRef]
- Ogborn, M.R.; Crocker, J.F.; Barnard, D.R. Plasma therapy for severe hemolytic-uremic syndrome in children in Atlantic Canada. CMAJ 1990, 143, 1323–1326. [Google Scholar] [CrossRef]
- Boyle, T.E.; Holowaychuk, M.K.; Adams, A.K.; Marks, S.L. Treatment of Three Cats with Hyperviscosity Syndrome and Congestive Heart Failure Using Plasmapheresis. J. Am. Anim. Hosp. Assoc. 2011, 47, 50–55. [Google Scholar] [CrossRef]
- Thielen, L.A.; Chen, J.; Jing, G.; Moukha-Chafiq, O.; Xu, G.; Jo, S.; Grayson, T.B.; Lu, B.; Li, P.; Augelli-Szafran, C.E.; et al. Identification of an Anti-diabetic, Orally Available Small Molecule that Regulates TXNIP Expression and Glucagon Action. Cell. Metab. 2020, 32, 353–365. [Google Scholar] [CrossRef]
- Manzoni, A.G.; Passos, D.F.; Doleski, P.H.; Leitemperger, J.W.; Loro, V.L.; Leal, D.B.R. Purine Metabolism in Platelets and Heart Cells of Hyperlipidemic Rats. Cardiovasc. Drugs Ther. 2020, 34, 813–821. [Google Scholar] [CrossRef]
- McCrindle, B.W.; Gidding, S.S. What Should Be the Screening Strategy for Familial Hypercholesterolemia? N. Engl. J. Med. 2016, 375, 1685–1686. [Google Scholar] [CrossRef]
- Cicero, A.F.G.; Fogacci, F.; Hernandez, A.V.; Banach, M.; on behalf of the Lipid and Blood Pressure Meta-Analysis Collaboration (LBPMC) Group and the International Lipid Expert Panel (ILEP). Efficacy and safety of bempedoic acid for the treatment of hypercholesterolemia: A systematic review and meta-analysis. PLoS Med. 2020, 17, e1003121. [Google Scholar] [CrossRef]
- Ponticelli, C.; Podestà, M.A.; Moroni, G. Hyperuricemia as a trigger of immune response in hypertension and chronic kidney disease. Kidney. Int. 2020, 98, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.W.; Lau, S.F.; Chin, C.S.L.; Ahmad, N.I.; Rahman, N.-A.; Kuppusamy, K.; Omar, S.; Radzi, R. Clinicopathological and radiographic features in 40 cats diagnosed with pulmonary and cutaneous Rhodococcus equi infection (2012–2018). J. Feline Med. Surg. 2019, 22, 774–790. [Google Scholar] [CrossRef] [PubMed]
- Weidmann, H.; Heikaus, L.; Long, A.T.; Naudin, C.; Schlüter, H.; Renné, T. The plasma contact system, a protease cascade at the nexus of inflammation, coagulation and immunity. Biochim. Biophys. Acta Bioenerg. 2017, 1864, 2118–2127. [Google Scholar] [CrossRef] [PubMed]
- McDonald, B.; Dunbar, M. Platelets and Intravascular Immunity: Guardians of the Vascular Space during Bloodstream Infections and Sepsis. Front. Immunol. 2019, 10, 2400. [Google Scholar] [CrossRef] [PubMed]
- Wójcik, P.; Biernacki, M.; Wroński, A.; Łuczaj, W.; Waeg, G.; Zarković, N.; Skrzydlewska, E. Altered Lipid Metabolism in Blood Mononuclear Cells of Psoriatic Patients Indicates Differential Changes in Psoriasis Vulgaris and Psoriatic Arthritis. Int. J. Mol. Sci. 2019, 20, 4249. [Google Scholar] [CrossRef] [PubMed]
- Nagy, T.; Fisi, V.; Frank, D.; Kátai, E.; Nagy, Z.; Miseta, A. Hyperglycemia-Induced Aberrant Cell Proliferation; A Metabolic Challenge Mediated by Protein O-GlcNAc Modification. Cells 2019, 8, 999. [Google Scholar] [CrossRef] [PubMed]
- Duan, Q.; Li, H.; Gao, C.; Zhao, H.; Wu, S.; Wu, H.; Wang, C.; Shen, Q.; Yin, T. High glucose promotes pancreatic cancer cells to escape from immune surveillance via AMPK-Bmi1-GATA2-MICA/B pathway. J. Exp. Clin. Cancer Res. 2019, 38, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Zimman, A.; Gao, D.; Byzova, T.V.; Podrez, E.A. TLR2 Plays a Key Role in Platelet Hyperreactivity and Accelerated Thrombosis Associated With Hyperlipidemia. Circ. Res. 2017, 121, 951–962. [Google Scholar] [CrossRef]
- Proto, J.D.; Doran, A.C.; Subramanian, M.; Wang, H.; Zhang, M.; Sozen, E.; Rymond, C.C.; Kuriakose, G.; D’Agati, V.; Winchester, R.; et al. Hypercholesterolemia induces T cell expansion in humanized immune mice. J. Clin. Investig. 2018, 128, 2370–2375. [Google Scholar] [CrossRef]
- Mailer, R.K.; Gisterå, A.; Polyzos, K.A.; Ketelhuth, D.F.; Hansson, G.K. Hypercholesterolemia Induces Differentiation of Regulatory T Cells in the Liver. Circ. Res. 2017, 120, 1740–1753. [Google Scholar] [CrossRef]
- Tall, A.R.; Yvan-Charvet, L. Cholesterol, inflammation and innate immunity. Nat. Rev. Immunol. 2015, 15, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Tzou, P. How Drosophila combats microbial infection: A model to study innate immunity and host–pathogen interactions. Curr. Opin. Microbiol. 2002, 5, 102–110. [Google Scholar] [CrossRef]
- Strand, M.R. The insect cellular immune response. Insect Sci. 2008, 15, 1–14. [Google Scholar] [CrossRef]
- Lu, A.; Zhang, Q.; Zhang, J.; Yang, B.; Wu, K.; Xie, W.; Luan, Y.-X.; Ling, E. Insect prophenoloxidase: The view beyond immunity. Front. Physiol. 2014, 5, 252. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Lu, Z. Immune responses to bacterial and fungal infections in the silkworm, Bombyx mori. Dev. Comp. Immunol. 2018, 83, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Cerenius, L.; Lee, B.L.; Söderhäll, K. The proPO-system: Pros and cons for its role in invertebrate immunity. Trends Immunol. 2008, 29, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Binggeli, O.; Neyen, C.; Poidevin, M.; Lemaitre, B. Prophenoloxidase Activation Is Required for Survival to Microbial Infections in Drosophila. PLoS Pathog. 2014, 10, e1004067. [Google Scholar] [CrossRef]
- Banerjee, U.; Girard, J.R.; Goins, L.M.; Spratford, C.M. Drosophila as a genetic model for hematopoiesis. Genetics 2019, 211, 367–417. [Google Scholar] [CrossRef]
- De Gregorio, E.; Spellman, P.T.; Tzou, P.; Rubin, G.M.; Lemaitre, B. The Toll and Imd pathways are the major regulators of the immune response in Drosophila. EMBO J. 2002, 21, 2568–2579. [Google Scholar] [CrossRef]
- Hoffmann, J.A.; Reichhart, J.-M. Drosophila innate immunity: An evolutionary perspective. Nat. Immunol. 2002, 3, 121–126. [Google Scholar] [CrossRef]
- Chowdhury, M.; Li, C.-F.; He, Z.; Lu, Y.; Liu, X.-S.; Wang, Y.-F.; Ip, Y.T.; Strand, M.R.; Yu, X.-Q. Toll family members bind multiple Spätzle proteins and activate antimicrobial peptide gene expression in Drosophila. J. Biol. Chem. 2019, 294, 10172–10181. [Google Scholar] [CrossRef]
- Valanne, S.; Wang, J.-H.; Rämet, M. The Drosophila Toll Signaling Pathway. J. Immunol. 2011, 186, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Myllymäki, H.; Valanne, S.; Rämet, M. The Drosophila Imd Signaling Pathway. J. Immunol. 2014, 192, 3455–3462. [Google Scholar] [CrossRef] [PubMed]
- Hillyer, J.F. Insect immunology and hematopoiesis. Dev. Comp. Immunol. 2016, 58, 102–118. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-D.; Wang, Y.-F.; Wang, Y.-L.; Li, Q.-Y.; Ma, H.-Y.; Wang, L.; Sima, Y.-H.; Xu, S.-Q. Induced Hyperproteinemia and Its Effects on the Remodeling of Fat Bodies in Silkworm, Bombyx mori. Front. Physiol. 2018, 9, 302. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-F.; Chen, X.-D.; Wang, G.; Li, Q.-Y.; Liang, X.-Y.; Sima, Y.-H.; Xu, S.-Q. Influence of hyperproteinemia on reproductive development in an invertebrate model. Int. J. Biol. Sci. 2019, 15, 2170–2181. [Google Scholar] [CrossRef]
- Magid, J.H. Chronic Hyperproteinemia Associated with a Probable Abdominal Abscess in an Appaloosa Stallion. Veter- Clin. N. Am. Equine Pr. 2006, 22, 53–60. [Google Scholar] [CrossRef]
- Riemer, F.; Kuehner, K.A.; Ritz, S.; Sauter-Louis, C.; Hartmann, K. Clinical and laboratory features of cats with feline infectious peritonitis—A retrospective study of 231 confirmed cases (2000–2010). J. Feline Med. Surg. 2016, 18, 348–356. [Google Scholar] [CrossRef]
- He, Y.; Xu, X.; Qiu, J.; Yin, W.; Sima, Y.; Xu, S.-Q. Bombyx mori used as a fast detection model of liver melanization after a clinical drug—Acetaminophen exposure. J. Asia Pacific Éntomol. 2020, 23, 177–185. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, W.; Xu, J.; Yang, W.; Li, Q.; Zhong, Y.; Cao, Y.; Yu, X.-Q.; Deng, X. Regulation of antimicrobial peptide genes via insulin-like signaling pathway in the silkworm Bombyx mori. Insect Biochem. Mol. Biol. 2018, 103, 12–21. [Google Scholar] [CrossRef]
- Barger, S.R.; Gauthier, N.C.; Krendel, M. Squeezing in a Meal: Myosin Functions in Phagocytosis. Trends. Cell. Biol. 2020, 30, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Flannagan, R.S.; Harrison, R.E.; Yip, C.M.; Jaqaman, K.; Grinstein, S. Dynamic macrophage “probing” is required for the efficient capture of phagocytic targets. J. Cell Biol. 2010, 191, 1205–1218. [Google Scholar] [CrossRef] [PubMed]
- Jafar, N.; Edriss, H.; Nugent, K. The Effect of Short-Term Hyperglycemia on the Innate Immune System. Am. J. Med. Sci. 2016, 351, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Kritikou, E.; Van Der Heijden, T.; Swart, M.; Van Duijn, J.; Slütter, B.; Wezel, A.; Smeets, H.J.; Maffia, P.; Kuiper, J.; Bot, I. Hypercholesterolemia Induces a Mast Cell–CD4+ T Cell Interaction in Atherosclerosis. J. Immunol. 2019, 202, 1531–1539. [Google Scholar] [CrossRef]
- Wu, Q.; Patočka, J.; Kuča, K. Insect antimicrobial peptides, a mini review. Toxins 2018, 10, 461. [Google Scholar] [CrossRef]
- Lu, Y.; Su, F.; Li, Q.; Zhang, J.; Li, Y.; Tang, T.; Hu, Q.; Yu, X.-Q. Pattern recognition receptors in Drosophila immune responses. Dev. Comp. Immunol. 2020, 102, 103468. [Google Scholar] [CrossRef]
- Yi, H.-Y.; Chowdhury, M.; Huang, Y.-D.; Yu, X.-Q. Insect antimicrobial peptides and their applications. Appl. Microbiol. Biotechnol. 2014, 98, 5807–5822. [Google Scholar] [CrossRef]
- Hetru, C.; Hoffmann, J.A. NF- B in the Immune Response of Drosophila. Cold Spring Harb. Perspect. Biol. 2009, 1, a000232. [Google Scholar] [CrossRef]
- Zaidman-Rémy, A.; Poidevin, M.; Hervé, M.; Welchman, D.P.; Paredes, J.C.; Fahlander, C.; Steiner, H.; Mengin-Lecreulx, D.; Lemaitre, B. Drosophila Immunity: Analysis of PGRP-SB1 Expression, Enzymatic Activity and Function. PLoS ONE 2011, 6, e17231. [Google Scholar] [CrossRef]
- Zhao, P.; Xia, F.; Jiang, L.; Guo, H.; Xu, G.; Sun, Q.; Wang, B.; Wang, Y.; Lu, Z.; Xia, Q. Enhanced antiviral immunity against Bombyx mori cytoplasmic polyhedrosis virus via overexpression of peptidoglycan recognition protein S2 in transgenic silkworms. Dev. Comp. Immunol. 2018, 87, 84–89. [Google Scholar] [CrossRef]
- Ma, X.; Li, X.; Dong, S.; Xia, Q.; Wang, F. A Fas associated factor negatively regulates anti-bacterial immunity by promoting Relish degradation in Bombyx mori. Insect Biochem. Mol. Biol. 2015, 63, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Yamamoto, M.; Moriyama, Y.; Yamao, M.; Furukawa, S.; Sagisaka, A.; Nakazawa, H.; Mori, H.; Yamakawa, M. A novel Rel protein and shortened isoform that differentially regulate antibacterial peptide genes in the silkworm Bombyx mori. Biochim. Biophys. Acta. 2005, 1730, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Yamakawa, M. Regulation of the innate immune responses in the silkworm, Bombyx mori. ISJ-Invert. Surviv. J. 2011, 8, 59–69. [Google Scholar]
- Furukawa, S.; Tanaka, H.; Nakazawa, H.; Ishibashi, J.; Shono, T.; Yamakawa, M. Inducible gene expression of moricin, a unique antibacterial peptide from the silkworm (Bombyx mori). Biochem. J. 1999, 340. [Google Scholar] [CrossRef]
- Tanaka, H.; Sagisaka, A.; Fujita, K.; Furukawa, S.; Ishibashi, J.; Yamakawa, M. BmEts upregulates promoter activity of lebocin in Bombyx mori. Insect Biochem. Mol. Biol. 2012, 42, 474–481. [Google Scholar] [CrossRef]
- Shakeel, M.; Xu, X.; De Mandal, S.; Jin, F. Role of serine protease inhibitors in insect-host-pathogen interactions. Arch. Insect Biochem. Physiol. 2019, 102, e21556. [Google Scholar] [CrossRef]
- Waterhouse, R.M.; Kriventseva, E.V.; Meister, S.; Xi, Z.; Alvarez, K.S.; Bartholomay, L.C.; Barillas-Mury, C.; Bian, G.; Blandin, S.; Christensen, B.M.; et al. Evolutionary Dynamics of Immune-Related Genes and Pathways in Disease-Vector Mosquitoes. Science 2007, 316, 1738–1743. [Google Scholar] [CrossRef]
- Cooper, D.; Wuebbolt, C.; Heryanto, C.; Eleftherianos, I. The prophenoloxidase system in Drosophila participates in the anti-nematode immune response. Mol. Immunol. 2019, 109, 88–98. [Google Scholar] [CrossRef]
- Li, J.; Ma, L.; Lin, Z.; Zou, Z.; Lu, Z. Serpin-5 regulates prophenoloxidase activation and antimicrobial peptide pathways in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2016, 73, 27–37. [Google Scholar] [CrossRef]
- Yuan, C.; Xing, L.; Wang, M.; Wang, X.; Yin, M.; Wang, Q.; Hu, Z.; Zou, Z. Inhibition of melanization by serpin-5 and serpin-9 promotes baculovirus infection in cotton bollworm Helicoverpa armigera. PLoS. Pathog. 2017, 13, e1006645. [Google Scholar] [CrossRef]
- De Gregorio, E.; Han, S.-J.; Lee, W.-J.; Baek, M.-J.; Osaki, T.; Kawabata, S.-I.; Lee, B.-L.; Iwanaga, S.; Lemaitre, B.; Brey, P.T. An Immune-Responsive Serpin Regulates the Melanization Cascade in Drosophila. Dev. Cell. 2002, 3, 581–592. [Google Scholar] [CrossRef]
- Kondo, Y.; Yoda, S.; Mizoguchi, T.; Ando, T.; Yamaguchi, J.; Yamamoto, K.; Banno, Y.; Fujiwara, H. Toll ligand Spätzle3 controls melanization in the stripe pattern formation in caterpillars. Proc. Natl. Acad. Sci. USA 2017, 114, 8336–8341. [Google Scholar] [CrossRef]
- Senftleben, U.; Cao, Y.; Xiao, G.; Greten, F.R.; Krähn, G.; Bonizzi, G.; Chen, Y.; Hu, Y.; Fong, A.; Sun, S.-C.; et al. Activation by IKKalpha of a Second, Evolutionary Conserved, NF-kappa B Signaling Pathway. Science 2001, 293, 1495–1499. [Google Scholar] [CrossRef] [PubMed]
- Van Uden, P.; Kenneth, N.S.; Webster, R.; Müller, H.A.; Mudie, S.; Rocha, S. Evolutionary Conserved Regulation of HIF-1β by NF-κB. PLoS Genet. 2011, 7, e1001285. [Google Scholar] [CrossRef] [PubMed]
- Brennan, J.J.; Gilmore, T.D. Evolutionary Origins of Toll-like Receptor Signaling. Mol. Biol. Evol. 2018, 35, 1576–1587. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-F.; Wang, G.; Li, J.-L.; Qu, Y.-X.; Liang, X.-Y.; Chen, X.-D.; Sima, Y.-H.; Xu, S.-Q. Influence of Hyperproteinemia on Insect Innate Immune Function of the Circulatory System in Bombyx mori. Biology 2021, 10, 112. https://doi.org/10.3390/biology10020112
Wang Y-F, Wang G, Li J-L, Qu Y-X, Liang X-Y, Chen X-D, Sima Y-H, Xu S-Q. Influence of Hyperproteinemia on Insect Innate Immune Function of the Circulatory System in Bombyx mori. Biology. 2021; 10(2):112. https://doi.org/10.3390/biology10020112
Chicago/Turabian StyleWang, Yong-Feng, Guang Wang, Jiang-Lan Li, Ya-Xin Qu, Xin-Yin Liang, Xue-Dong Chen, Yang-Hu Sima, and Shi-Qing Xu. 2021. "Influence of Hyperproteinemia on Insect Innate Immune Function of the Circulatory System in Bombyx mori" Biology 10, no. 2: 112. https://doi.org/10.3390/biology10020112
APA StyleWang, Y.-F., Wang, G., Li, J.-L., Qu, Y.-X., Liang, X.-Y., Chen, X.-D., Sima, Y.-H., & Xu, S.-Q. (2021). Influence of Hyperproteinemia on Insect Innate Immune Function of the Circulatory System in Bombyx mori. Biology, 10(2), 112. https://doi.org/10.3390/biology10020112