
Fitness Analysis and Transcriptome Profiling Following Repeated Mild Heat Stress of Varying Frequency in Drosophila melanogaster Females

 ,
, 
 ,
,  ,
,
Simple Summary
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Drosophila melanogaster Strain and Heat Stress Modes
2.2. Fertility and Longevity Analysis
2.3. Body Mass and Triglyceride Content Measurements
2.4. Carbohydrates Metabolism Evaluation
2.5. Capillary Feeding Assay
2.6. Statistical Analysis of Fitness and Body Composition Data
2.7. RNA Isolation, cDNA Library Construction and RNA Sequencing
2.8. Mapping Reads onto the Genome and FPKM Evaluation
2.9. Transcriptome Data Analysis
2.10. Quantitative Real-Time Polymerase Chain Reaction
3. Results
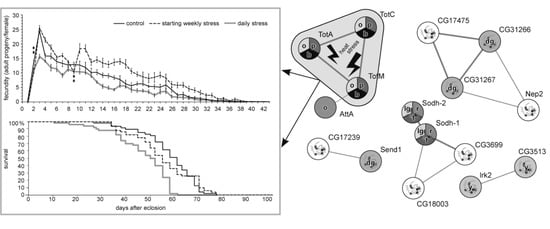
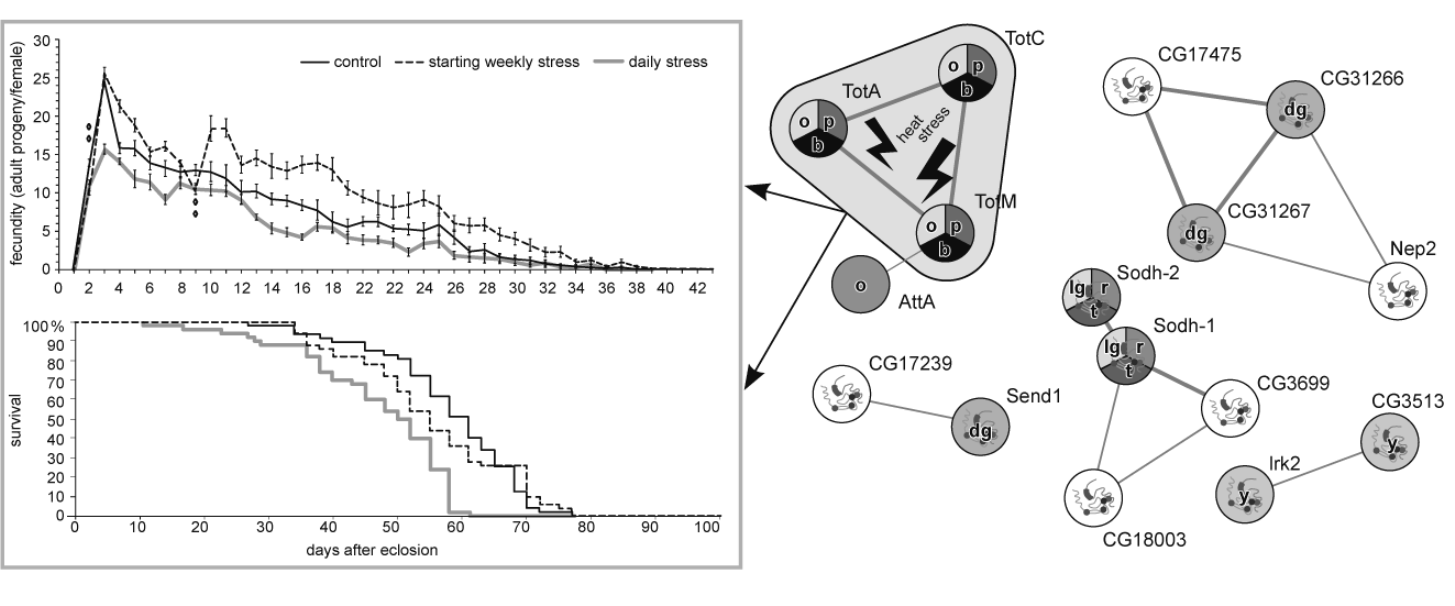
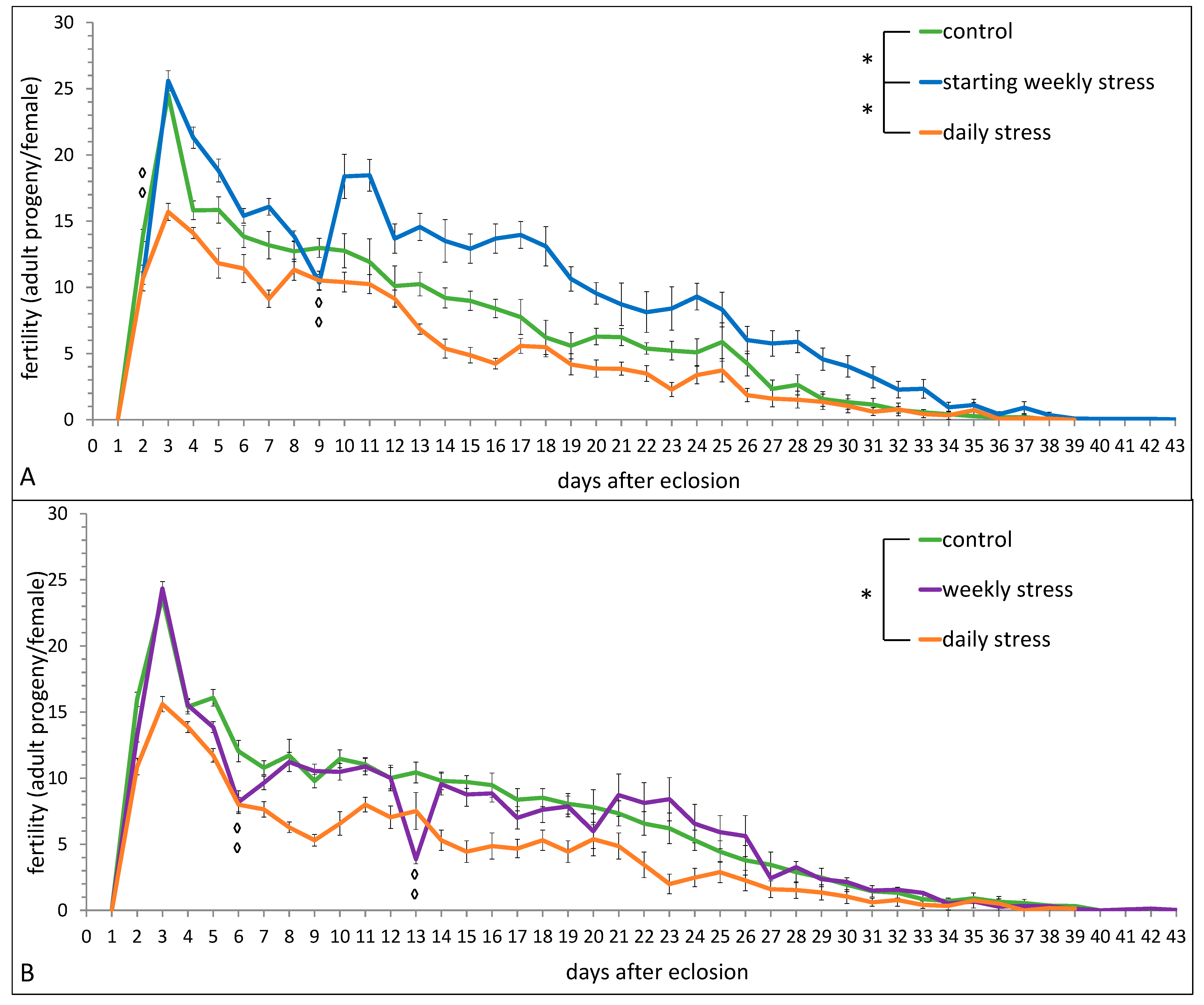
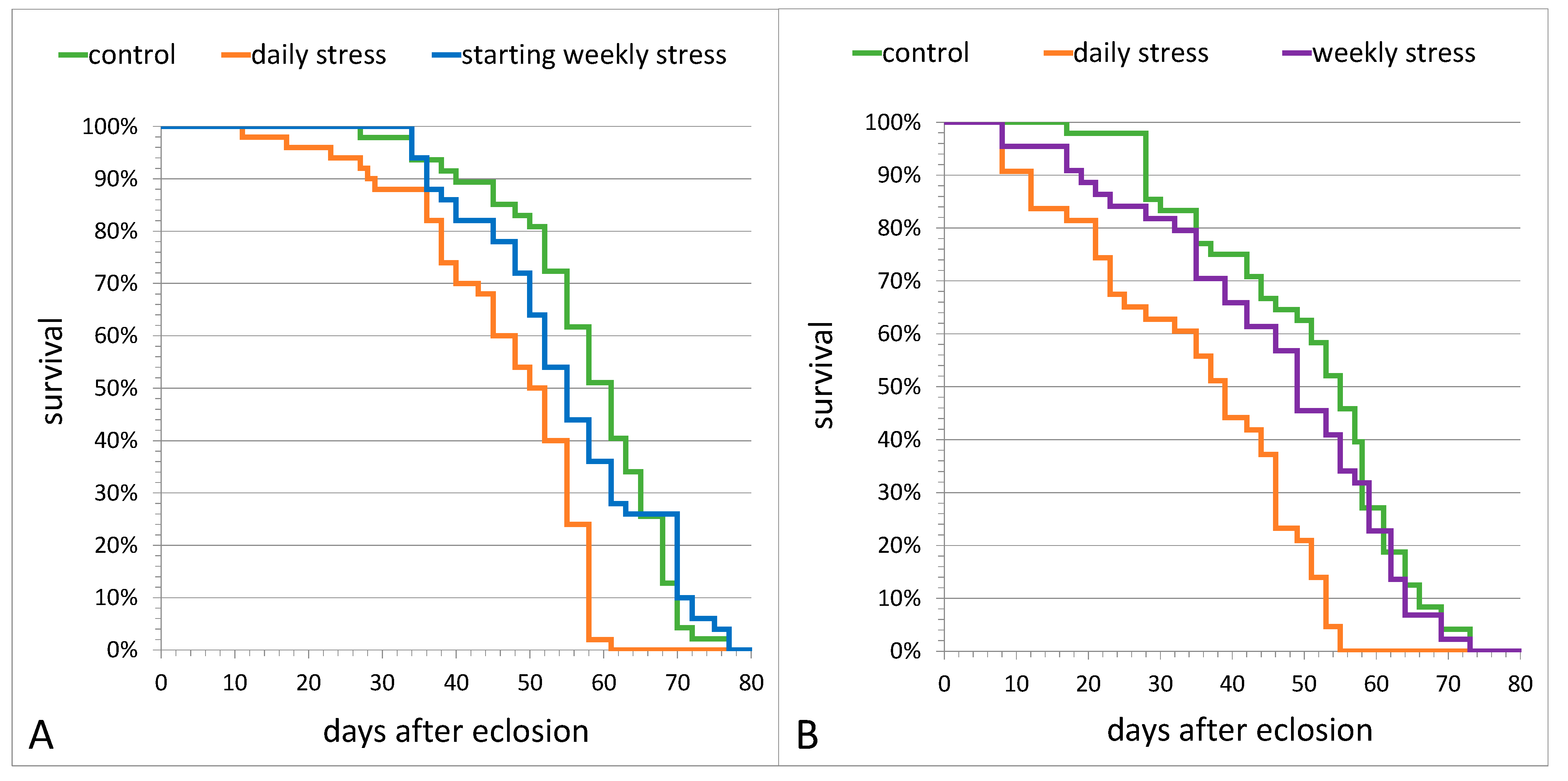
3.1. The Effects of Repeated Mild Stress Episodes of Varying Frequency on Fitness
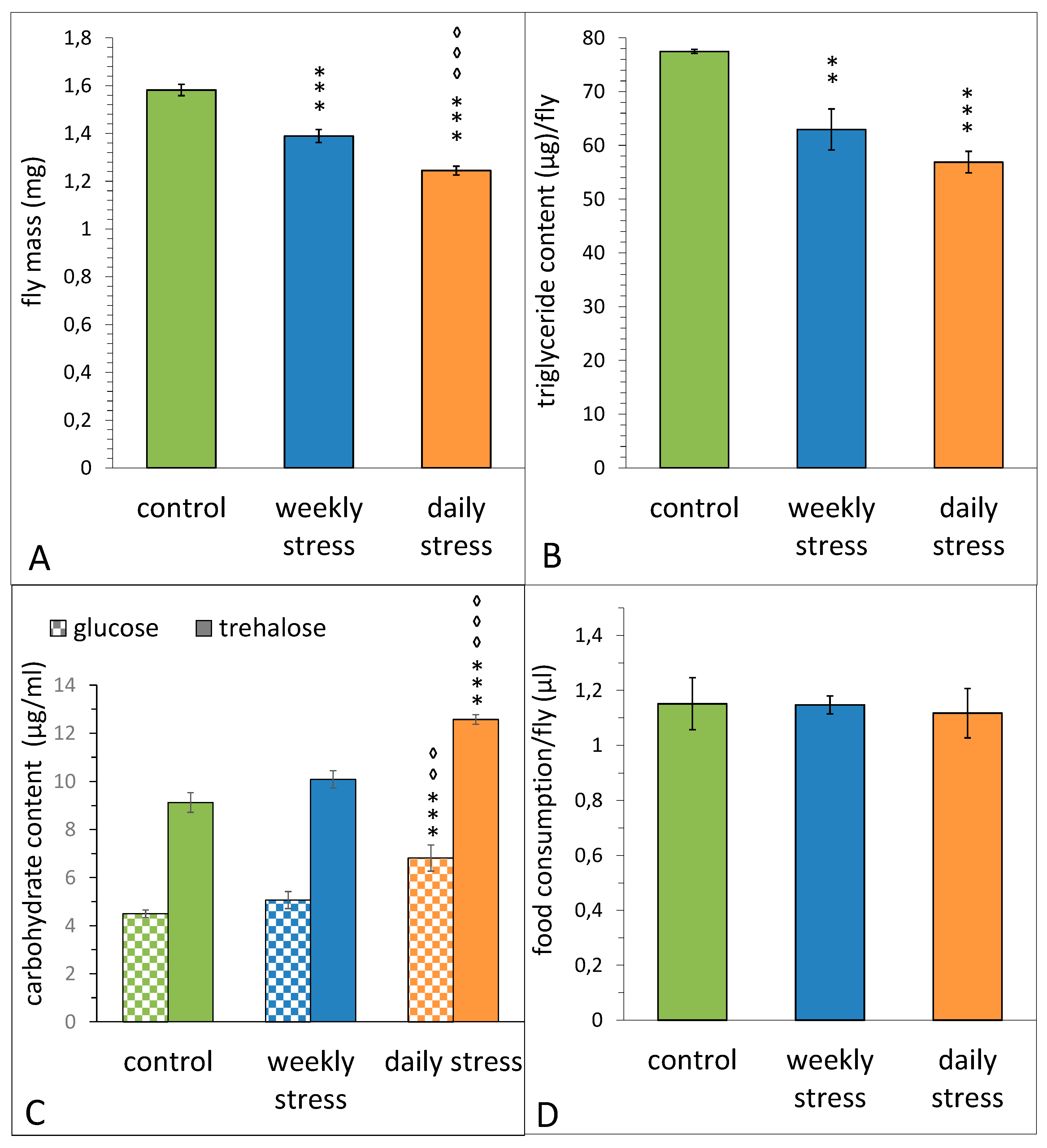
3.2. The Effects of Repeated Mild Stress Episodes of Varying Frequency on Appetite, Body Mass, Triglyceride and Carbohydrates Contents
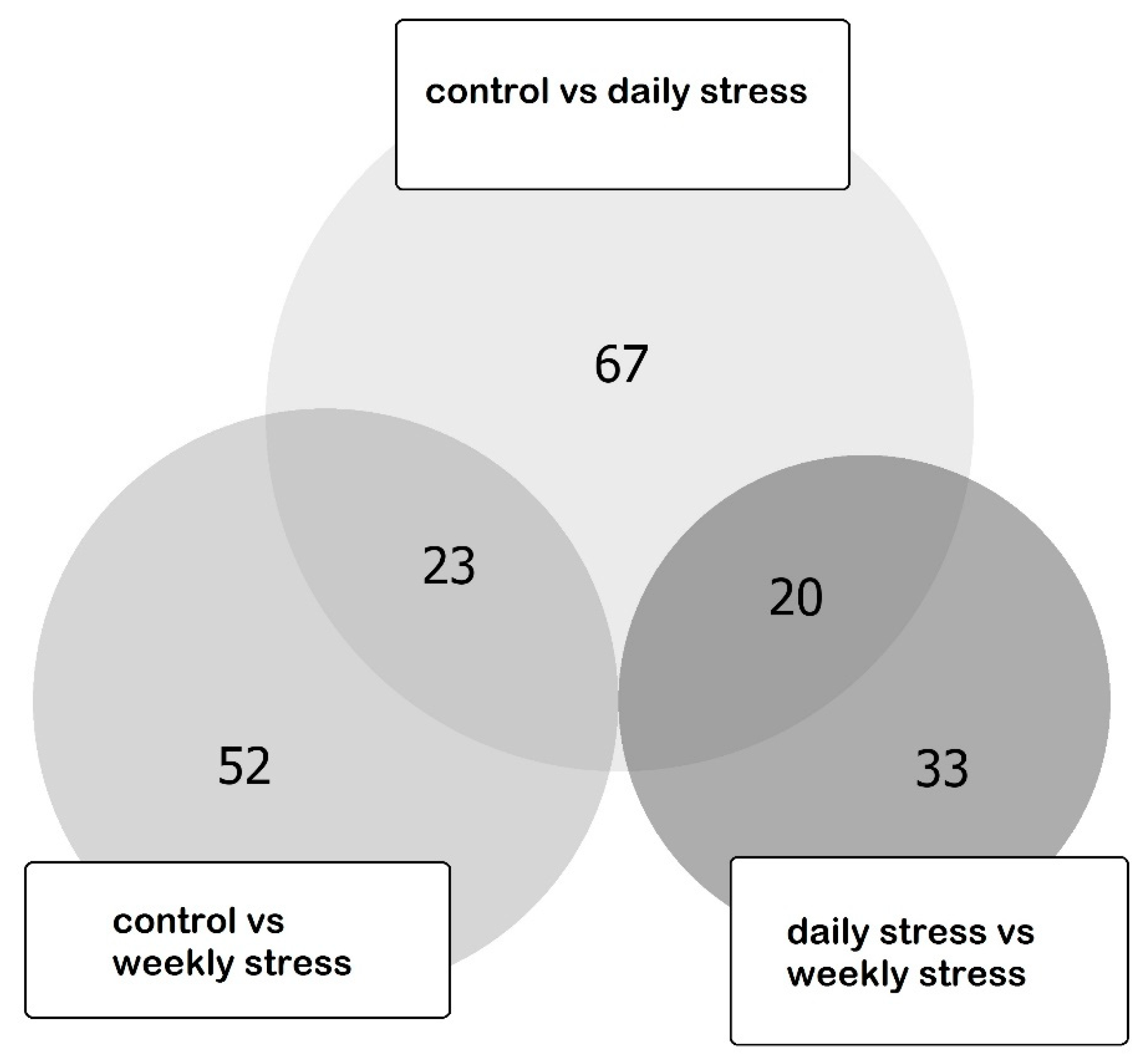
3.3. Differentially Expressed Genes (DEGs) Elucidated by RNA-Seq
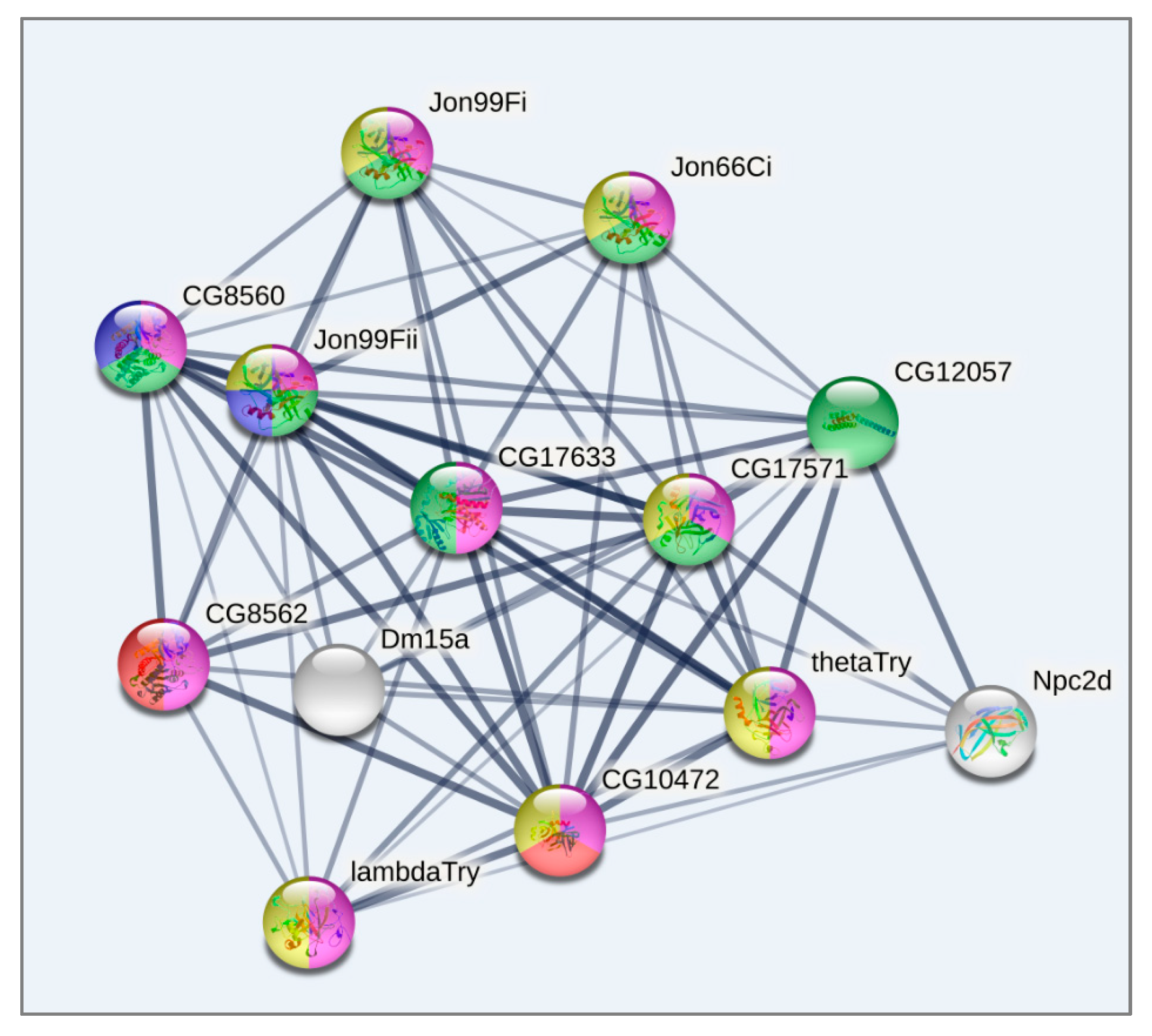
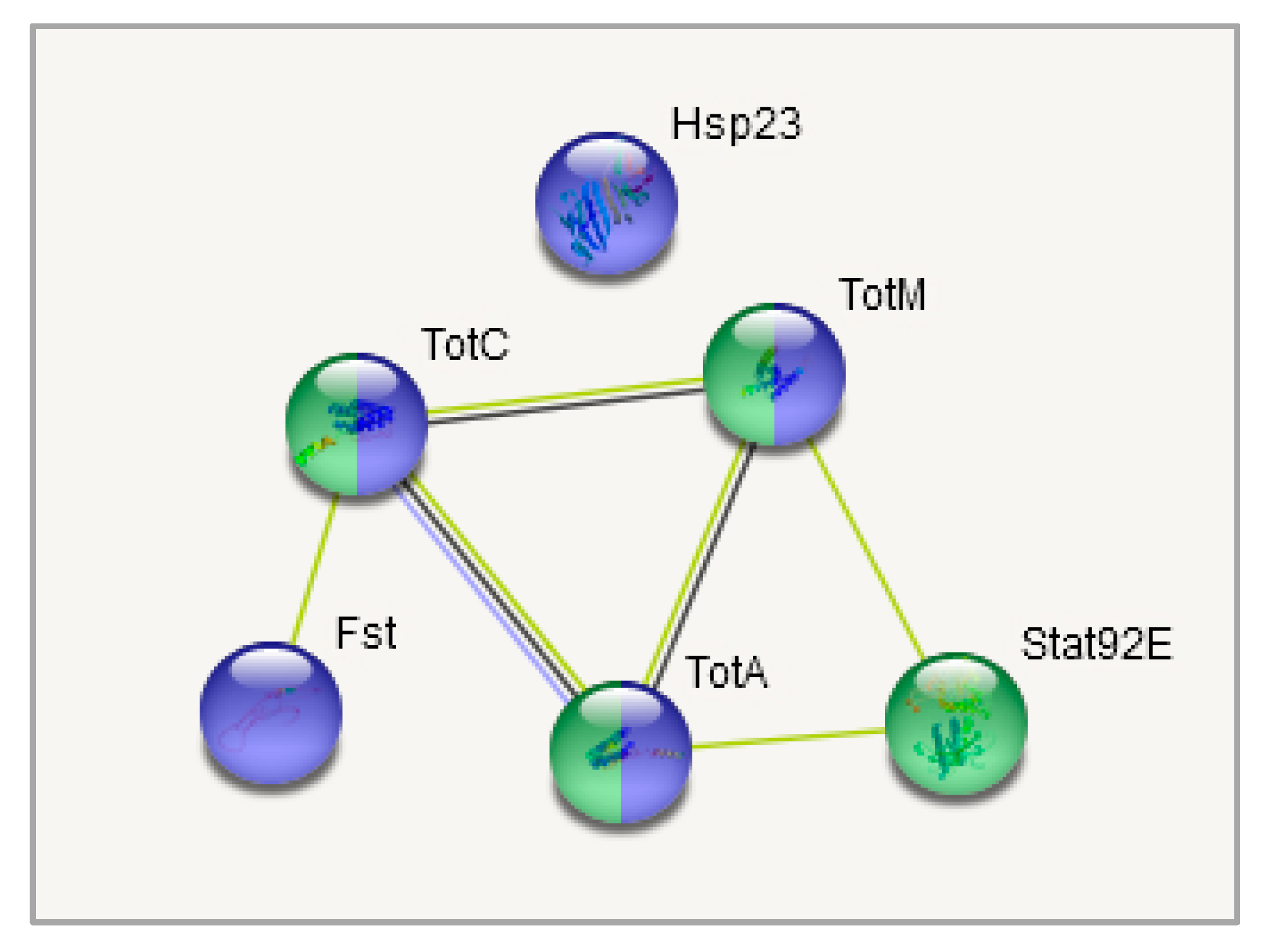
3.4. DEGs Clustering
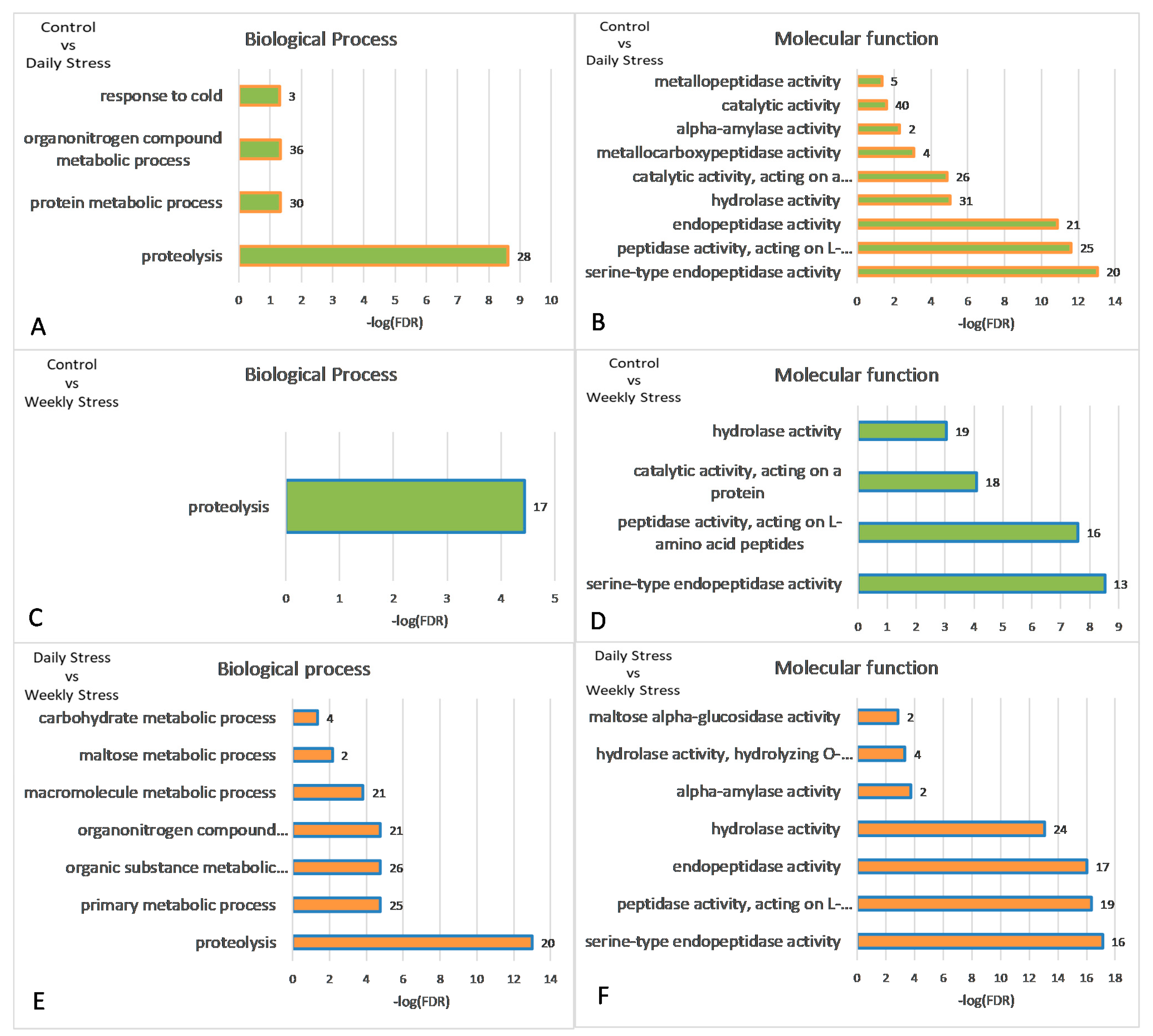
3.5. DEGs to GO
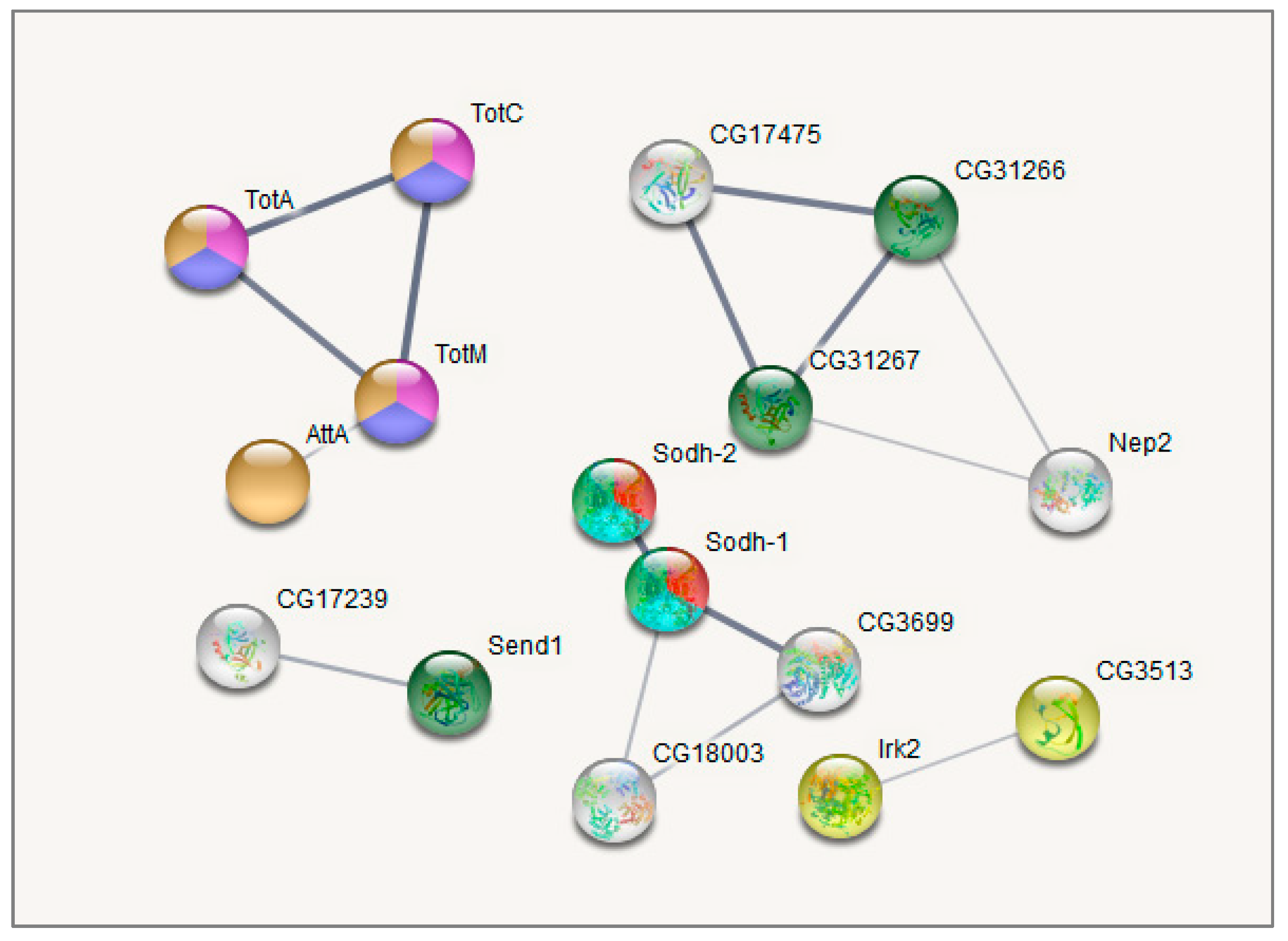
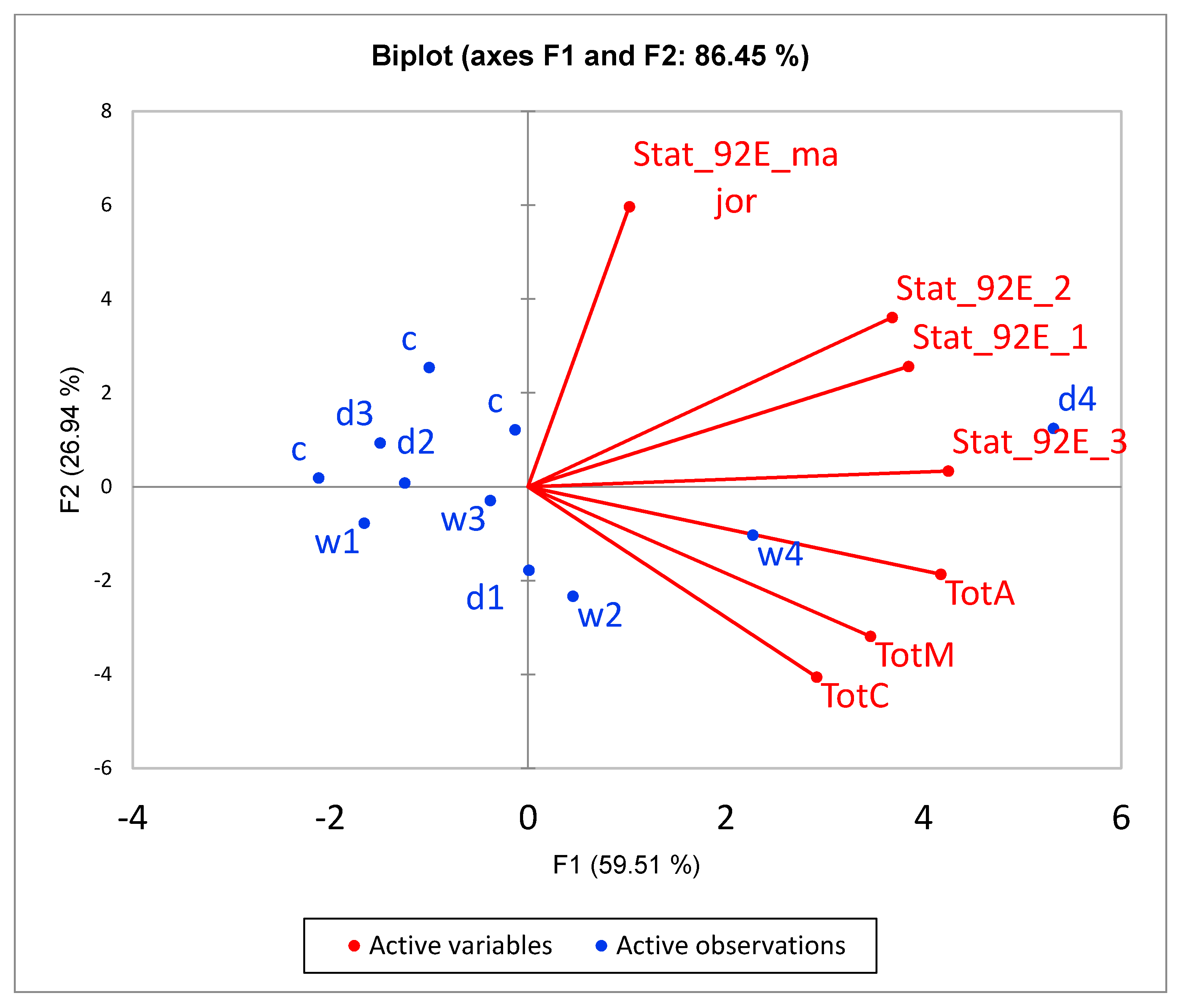
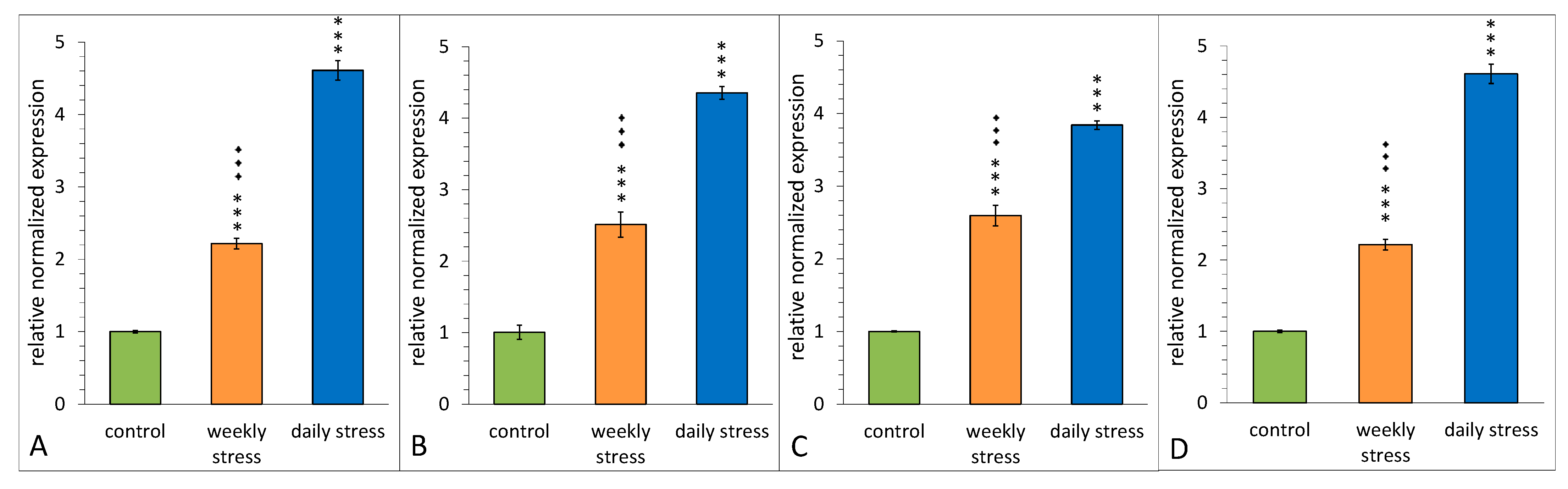
3.6. Confirmation of Altered Gene Expression by qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Evgen’ev, M.B.; Garbuz, D.G.; Zatsepina, O.G. Heat Shock Proteins and Whole Body Adaptation to Extreme Environments; Springer: Berlin, Germany; New York, NY, USA; London, UK, 2014; ISBN 9789401792356. [Google Scholar]
- Davis, R.J. Signal transduction by the JNK group of MAP kinases. Cell 2000, 103, 239–252. [Google Scholar] [CrossRef]
- Gonda, R.L.; Garlena, R.A.; Stronach, B. Drosophila Heat Shock Response Requires the JNK Pathway and Phosphorylation of Mixed Lineage Kinase at a Conserved Serine-Proline Motif. PLoS ONE 2012, 7, e42369. [Google Scholar] [CrossRef] [PubMed]
- Ekengren, S.; Hultmark, D. A family of Turandot-related genes in the humoral stress response of Drosophila. Biochem. Biophys. Res. Commun. 2001, 284, 998–1003. [Google Scholar] [CrossRef]
- Perić-Mataruga, V.; Nenadović, V.; Ivanović, J. Neurohormones in insect stress: A review. Arch. Biol. Sci. 2006, 58, 1–12. [Google Scholar] [CrossRef]
- Gruntenko, N.E.; Rauschenbach, I.Y. Interplay of juvenile hormone, 20-hydroxyecdisone and biogenic amines under normal and stress conditions and its effect on reproduction. J. Insect Physiol. 2008, 54, 902–908. [Google Scholar] [CrossRef]
- Gruntenko, N.E.; Rauschenbach, I.Y. The role of insulin signalling in the endocrine stress response in Drosophila melanogaster: A mini-review. Gen. Comp. Endocrinol. 2018, 258, 134–139. [Google Scholar] [CrossRef]
- Johnson, E.C.; White, M.P. Stressed-out insects: Hormonal actions and behavioral modifications. In Hormones, Brain and Behavior; Pfaff, D.W., Arnold, A.P., Etgen, A.M., Fahrbach, S.E., Rubin, R.T., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 1069–1096. ISBN 9780080887838. [Google Scholar]
- Miyashita, A.; Adamo, S.A. Stayin’ Alive: Endocrinological Stress Responses in Insects. In Advances in Invertebrate Endocrinology, 1st ed.; Apple Academic Press: Toronto, ON, Canada, 2020; pp. 283–324. ISBN 9781003029861. [Google Scholar]
- Gruntenko, N.E.; Bownes, M.; Terashima, J.; Sukhanova, M.Z.; Rauschenbach, I.Y. Heat stress affects oogenesis differently in wild type Drosophila virilis and a mutant with altered juvenile hormone and 20-hydroxyecdysone levels. Insect Mol. Biol. 2003, 12, 393–404. [Google Scholar] [CrossRef]
- Heimann, M.; Reichstein, M. Terrestrial ecosystem carbon dynamics and climate feedbacks. Nature 2008, 451, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Le Bourg, É. Using Drosophila melanogaster to study the positive effects of mild stress on aging. Exp. Gerontol. 2011, 46, 345–348. [Google Scholar] [CrossRef]
- Kuether, K.; Arking, R. Drosophila selected for extended longevity are more sensitive to heat shock. Age 1999, 22, 175–180. [Google Scholar] [CrossRef][Green Version]
- Minois, N.; Vaynberg, S. Fecundity and life span in transgenic Drosophila melanogaster overexpressing hsp70. Biogerontology 2002, 3, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Le Bourg, É.; Valenti, P.; Lucchetta, P.; Payre, F. Effects of mild heat shocks at young age on aging and longevity in Drosophila melanogaster. Biogerontology 2001, 2, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Khazaeli, A.A.; Tatar, M.; Pletcher, S.D.; Curtsinger, J.W. Heat-induced longevity extension in Drosophila. I. Heat treatment, mortality, and thermotolerance. J. Gerontol. Biol. Sci. 1997, 52, B48–B52. [Google Scholar] [CrossRef] [PubMed]
- Hercus, M.J.; Loeschcke, V.; Rattan, S.I.S. Lifespan extension of Drosophila melanogaster through hormesis by repeated mild heat stress. Biogerontology 2003, 4, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Burdina, E.V.; Adonyeva, N.V.; Karpova, E.K.; Rauschenbach, I.Y.; Menshanov, P.N.; Gruntenko, N.E. The effect of mild heat stress of different frequencies on the adaptability of Drosophila melanogaster females. Arch. Insect Biochem. 2019, 102, e21619. [Google Scholar] [CrossRef]
- Bogomolova, E.V.; Rauschenbach, I.Y.; Adonyeva, N.V.; Alekseev, A.A.; Faddeeva, N.V.; Gruntenko, N.E. Dopamine down-regulates activity of alkaline phosphatase in Drosophila: The role of D2-Like receptors. J. Insect Physiol. 2010, 56, 1155–1159. [Google Scholar] [CrossRef]
- Pendleton, R.G.; Rasheed, A.; Sardina, T.; Tully, T.; Hillman, R. Effects of tyrosine hydroxylase mutants on locomotor activity in Drosophila: A study in functional genomics. Behav. Genet. 2002, 32, 89–94. [Google Scholar] [CrossRef]
- Socha, R.; Kodrik, D.; Zemek, R. Stimulatory effects of bioamines norepinephrine and dopamine on locomotion of Pyrrhocoris apterus (L.): Is the adipokinetic hormone involved. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 151, 305–310. [Google Scholar] [CrossRef]
- Plaçais, P.; de Tredern, É.; Scheunemann, L.; Trannoy, S.; Goguel, V.; Han, K.; Isabel, G.; Preat, T. Upregulated energy metabolism in the Drosophila mushroom body is the trigger for long-term memory. Nat. Commun. 2017, 8, 15510. [Google Scholar] [CrossRef]
- Friedman, D.A.; Pilko, A.; Skowronska-Krawczyk, D.; Krasinska, K.; Parker, J.W.; Hirsh, J.; Gordon, D.M. The Role of Dopamine in the Collective Regulation of Foraging in Harvester Ants. iScience 2018, 8, 283–294. [Google Scholar] [CrossRef]
- Wroblewski, V.J.; Harshman, L.G.; Hanzlik, T.N.; Hammock, B.D. Regulation of juvenile hormone esterase gene expression in the tobacco budworm (Heliothis virescens). Arch. Biochem. Biophys. 1990, 278, 461–466. [Google Scholar] [CrossRef]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef]
- Liu, Z.; Huang, X. Lipid metabolism in Drosophila: Development and disease. Acta Biochim. Biophys. Sin. 2013, 45, 44–50. [Google Scholar] [CrossRef]
- Rivera Pérez, C.; Clifton, M.E.; Noriega, F.G.; Jindra, M. Juvenile Hormone Regulation and Action. In Advances in Invertebrate Endocrinology, 1st ed.; Apple Academic Press: Toronto, ON, Canada, 2020; pp. 1–76. ISBN 9781003029861. [Google Scholar]
- Rauschenbach, I.Y.; Gruntenko, N.E.; Khlebodarova, T.M.; Mazurov, M.M.; Grenback, L.G.; Sukhanova, M.J.; Shumnaja, L.V.; Zakharov, I.K.; Hammock, B.D. The role of the degradation system of the juvenile hormone in the reproduction of Drosophila under stress J. Insect Physiol. 1996, 42, 735–742. [Google Scholar] [CrossRef]
- Jørgensen, K.T.; Sørensen, J.G.; Bundgaard, J. Heat tolerance and the effect of mild heat stress on reproductive characters in Drosophila buzzatii males J. Therm. Biol. 2006, 31, 280–286. [Google Scholar] [CrossRef]
- Karpova, E.K.; Komyshev, E.G.; Genaev, M.A.; Adonyeva, N.V.; Afonnikov, D.A.; Eremina, M.A.; Gruntenko, N.E. Quantifying Drosophila adults with the use of a smartphone. Biol. Open 2020, 9, bio054452. [Google Scholar] [CrossRef]
- Mukherjee, S.; Mishra, M. Biochemical Estimation to Detect the Metabolic Pathways of Drosophila. In Fundamental Approaches to Screen Abnormalities in Drosophila Springer Protocols Handbooks; Mishra, M., Ed.; Springer: New York, NY, USA, 2020; pp. 135–149. ISBN 978-1-4939-9755-8. [Google Scholar]
- Karpova, E.K.; Eremina, M.A.; Pirozhkova, D.S.; Gruntenko, N.E. Stress-related hormones affect carbohydrate metabolism in Drosophila melanogaster females. Arch. Insect Biochem. Physiol. 2019, 101, e21540. [Google Scholar] [CrossRef]
- Ja, W.W.; Carvalho, G.B.; Mak, E.M.; De la Rosa, N.N.; Fang, A.Y.; Liong, J.C.; Brummel, T.; Benzer, S. Prandiology of Drosophila and the CAFE assay. Proc. Natl. Acad. Sci. USA 2007, 104, 8253–8256. [Google Scholar] [CrossRef]
- Williams, M.J.; Wang, Y.; Klockars, A.; Lind, P.M.; Fredriksson, R.; Schiöth, H.B. Exposure to Bisphenol A Affects Lipid Metabolism in Drosophila melanogaster. Basic Clin. Pharmacol. Toxicol. 2014, 114, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Han, D.; Hou, Y.; Chen, H.; Chen, Z. Statistical Inference Methods for Two Crossing Survival Curves: A Comparison of Methods. PLoS ONE 2015, 10, e0116774. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Dobin, A.; Gingeras, T.R. Mapping RNA-seq Reads with STAR. Curr. Protoc. Bioinform. 2015, 51, 11.14.1–11.14.19. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Jensen, L.J.; Kuhn, M.; Stark, M.; Chaffron, S.; Creevey, C.; Muller, J.; Doerks, T.; Julien, P.; Roth, A.; Simonovic, M.; et al. STRING 8—A global view on proteins and their functional interactions in 630 organisms. Nucleic Acids Res. 2009, 37, D412–D416. [Google Scholar] [CrossRef] [PubMed]
- Erkosar, B.; Kolly, S.; van der Meer, J.R.; Kawecki, T.J. Adaptation to Chronic Nutritional Stress Leads to Reduced Dependence on Microbiota in Drosophila melanogaster. mBio 2017, 8, e01496-17. [Google Scholar] [CrossRef]
- Zhong, W.; McClure, C.D.; Evans, C.R.; Mlynski, D.T.; Immonen, E.; Ritchie, M.G.; Priest, N.K. Immune anticipation of mating in Drosophila: Turandot M promotes immunity against sexually transmitted fungal infections. Proc. R. Soc. B 2013, 280, 20132018. [Google Scholar] [CrossRef] [PubMed]
- Kučerová, L.; Kubrak, O.I.; Bengtsson, J.M.; Strnad, H.; Nylin, S.; Theopold, U.; Nässel, D.R. Slowed aging during reproductive dormancy is reflected in genome-wide transcriptome changes in Drosophila melanogaster. BMC Genet. 2016, 17, 50. [Google Scholar] [CrossRef]
- Moskalev, A.; Zhikrivetskaya, S.; Krasnov, G.; Mikhail, S.; Ekaterina, P.; Dmitry, B.; Anton, D.; Darya, P.; Irina, S.; Eugenia, D.; et al. A comparison of the transcriptome of Drosophila melanogaster in response to entomopathogenic fungus, ionizing radiation, starvation and cold shock. BMC Genet. 2015, 16, S8. [Google Scholar] [CrossRef]
- Colinet, H.; Siaussat, D.; Bozzolan, F.; Bowler, K. Rapid decline of cold tolerance at young age is associated with expression of stress genes in Drosophila melanogaster. J. Exp. Biol. 2013, 216, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Manning, L.; Sheth, J.; Bridges, S.; Saadin, A.; Odinammadu, K.; Andrew, D.; Spencer, S.; Montell, D.; Starz-Gaiano, M. A hormonal cue promotes timely follicle cell migration by modulating transcription profiles. Mech. Dev. 2017, 148, 56–68. [Google Scholar] [CrossRef]
- Sørensen, J.G.; Nielsen, M.M.; Kruhøffer, M.; Justesen, J.; Loeschcke, V. Full genome gene expression analysis of the heat stress response in Drosophila melanogaster. Cell Stress Chaperones 2005, 10, 312–328. [Google Scholar] [CrossRef] [PubMed]
- Sarup, P.; Sørensen, P.; Loeschcke, V. The long-term effects of a life-prolonging heat treatment on the Drosophila melanogaster transcriptome suggest that heat shock proteins extend lifespan. Exp. Gerontol. 2014, 50, 34–39. [Google Scholar] [CrossRef] [PubMed]
- De Luca, M.; Roshina, N.V.; Geiger-Thornsberry, G.L.; Lyman, R.F.; Pasyukova, E.G.; Mackay, T.F. Dopa decarboxylase (Ddc) affects variation in Drosophila longevity. Nat. Genet. 2003, 34, 429–433. [Google Scholar] [CrossRef]
- Karpova, E.K.; Rauschenbach, I.Y.; Gruntenko, N.E. Comparative analysis of the fitness of Drosophila virilis lines contrasting in response to stress. Vavilovskii Zhurnal Genet. Sel. = Vavilov J. Genet. Breed. 2018, 22, 1090–1096. [Google Scholar] [CrossRef]
- Klepsatel, P.; Gáliková, M.; Xu, Y.; Kühnlein, R.P. Thermal stress depletes energy reserves in Drosophila. Sci. Rep. 2016, 6, 33667. [Google Scholar] [CrossRef]
- Eremina, M.A.; Menshanov, P.N.; Shishkina, O.D.; Gruntenko, N.E. Transcription factor dFOXO controls insulin pathway genes expression and lipids content under heat stress in Drosophila melanogaster. Vavilovskii Zhurnal Genet. Sel. = Vavilov J. Genet. Breed. 2021, 25, 465–471. [Google Scholar] [CrossRef]
- Overgaard, J.; Malmendal, A.; Sørensen, J.G.; Bundy, J.G.; Loeschcke, V.; Nielsen, N.C.; Holmstrup, M. Metabolomic profiling of rapid cold hardening and cold shock in Drosophila melanogaster. J. Insect Physiol. 2007, 53, 1218–1232. [Google Scholar] [CrossRef]
- Wolfson, J.L.; Murdock, L.L. Diversity in digestive proteinase activity among insects. J. Chem. Ecol. 1990, 16, 1089–1102. [Google Scholar] [CrossRef]
- Zatsepina, O.G.; Przhiboro, A.A.; Yushenova, I.A.; Shilova, V.; Zelentsova, E.S.; Shostak, N.G.; Evgen’ev, M.B.; Garbuz, D.G. A Drosophila heat shock response represents an exception rather than a rule amongst Diptera species. Insect Mol. Biol. 2016, 25, 431–449. [Google Scholar] [CrossRef]
- Marin, R.; Valet, J.P.; Tanguay, R.M. Hsp23 and hsp26 exhibit distinct spatial and temporal patterns of constitutive expression in Drosophila adults. Dev. Genet. 1993, 14, 69–77. [Google Scholar] [CrossRef]
- Kurapati, R.; Passananti, H.B.; Rose, M.R.; Tower, J. Increased hsp22 RNA levels in Drosophila lines genetically selected for increased longevity. J. Gerontol. A Biol. Sci. Med. Sci. 2000, 55, B552–B559. [Google Scholar] [CrossRef] [PubMed]
- Boardman, L.; Mitchell, K.A.; Terblanche, J.S.; Sørensen, J.G. A transcriptomics assessment of oxygen-temperature interactions reveals novel candidate genes underlying variation in thermal tolerance and survival. J. Insect Physiol. 2018, 106, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Lockwood, B.L.; Julick, C.R.; Montooth, K.L. Maternal loading of a small heat shock protein increases embryo thermal tolerance in Drosophila melanogaster. J. Exp. Biol. 2017, 220, 4492–4501. [Google Scholar] [CrossRef]
- Kawasaki, F.; Koonce, N.L.; Guo, L.; Fatima, S.; Qiu, C.; Moon, M.T.; Zheng, Y.; Ordway, R.W. Small heat shock proteins mediate cell-autonomous and -nonautonomous protection in a Drosophila model for environmental-stress-induced degeneration. Dis. Model. Mech. 2016, 9, 953–964. [Google Scholar] [CrossRef]
- Goto, S.G. A novel gene that is up-regulated during recovery from cold shock in Drosophila melanogaster. Gene 2001, 270, 259–264. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Gibbs, A.G.; Roberts, S.P. Gene transcription during exposure to, and recovery from, cold and desiccation stress in Drosophila melanogaster. Insect Mol. Biol. 2007, 16, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Udaka, H.; Ueda, C.; Goto, S.G. Survival rate and expression of Heat-shock protein 70 and Frost genes after temperature stress in Drosophila melanogaster lines that are selected for recovery time from temperature coma. J. Insect Physiol. 2010, 56, 1889–1894. [Google Scholar] [CrossRef]
- Colinet, H.; Lee, S.F.; Hoffmann, A. Functional characterization of the Frost gene in Drosophila melanogaster: Importance for recovery from chill coma. PLoS ONE 2010, 5, e10925. [Google Scholar] [CrossRef] [PubMed]
- Ekengren, S.; Tryselius, Y.; Dushay, M.S.; Liu, G.; Steiner, H.; Hultmark, D. A humoral stress response in Drosophila. Curr. Biol. 2001, 11, 714–718. [Google Scholar] [CrossRef]
- Agaisse, H.; Petersen, U.M.; Boutros, M.; Mathey-Prevot, B.; Perrimon, N. Signaling role of hemocytes in Drosophila JAK/STAT-dependent response to septic injury. Dev. Cell 2003, 5, 441–450. [Google Scholar] [CrossRef]
- Brun, S.; Vidal, S.; Spellman, P.; Takahashi, K.; Tricoire, H.; Lemaitre, B. The MAPKKK Mekk1 regulates the expression of Turandot stress genes in response to septic injury in Drosophila. Genes Cells 2006, 11, 397–407. [Google Scholar] [CrossRef]
- Salehipour-shirazi, G.; Ferguson, L.V.; Sinclair, B.J. Does cold activate the Drosophila melanogaster immune system? J. Insect Physiol. 2017, 96, 29–34. [Google Scholar] [CrossRef]
- Chow, J.; Kagan, J.C. The Fly Way of Antiviral Resistance and Disease Tolerance. Adv. Immunol. 2018, 140, 59–93. [Google Scholar] [CrossRef] [PubMed]
- Marshall, K.E.; Sinclair, B.J. Repeated stress exposure results in a survival-reproduction trade-off in Drosophila melanogaster. Proc. Biol. Sci. 2010, 277, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Diangelo, J.R.; Hughes, M.E.; Hogenesch, J.B.; Sehgal, A. The circadian clock interacts with metabolic physiology to influence reproductive fitness. Cell Metab. 2011, 13, 639–654. [Google Scholar] [CrossRef] [PubMed]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gruntenko, N.E.; Karpova, E.K.; Babenko, V.N.; Vasiliev, G.V.; Andreenkova, O.V.; Bobrovskikh, M.A.; Menshanov, P.N.; Babenko, R.O.; Rauschenbach, I.Y. Fitness Analysis and Transcriptome Profiling Following Repeated Mild Heat Stress of Varying Frequency in Drosophila melanogaster Females. Biology 2021, 10, 1323. https://doi.org/10.3390/biology10121323
Gruntenko NE, Karpova EK, Babenko VN, Vasiliev GV, Andreenkova OV, Bobrovskikh MA, Menshanov PN, Babenko RO, Rauschenbach IY. Fitness Analysis and Transcriptome Profiling Following Repeated Mild Heat Stress of Varying Frequency in Drosophila melanogaster Females. Biology. 2021; 10(12):1323. https://doi.org/10.3390/biology10121323
Chicago/Turabian StyleGruntenko, Nataly E., Evgenia K. Karpova, Vladimir N. Babenko, Gennady V. Vasiliev, Olga V. Andreenkova, Margarita A. Bobrovskikh, Petr N. Menshanov, Roman O. Babenko, and Inga Yu. Rauschenbach. 2021. "Fitness Analysis and Transcriptome Profiling Following Repeated Mild Heat Stress of Varying Frequency in Drosophila melanogaster Females" Biology 10, no. 12: 1323. https://doi.org/10.3390/biology10121323
APA StyleGruntenko, N. E., Karpova, E. K., Babenko, V. N., Vasiliev, G. V., Andreenkova, O. V., Bobrovskikh, M. A., Menshanov, P. N., Babenko, R. O., & Rauschenbach, I. Y. (2021). Fitness Analysis and Transcriptome Profiling Following Repeated Mild Heat Stress of Varying Frequency in Drosophila melanogaster Females. Biology, 10(12), 1323. https://doi.org/10.3390/biology10121323