
The Mediterranean Diet Positively Affects Resting Metabolic Rate and Salivary Microbiota in Human Subjects: A Comparison with the Vegan Regimen

 ,
, 
 ,
,  ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Indirect Calorimetry
2.2. Bacterial DNA Extraction and RT-PCR
2.3. Quantification of Absolute and Relative Bacterial DNA
2.4. Statistical Analysis
3. Results
3.1. Descriptive Statistics
3.2. Basal Metabolic Rate Evaluation
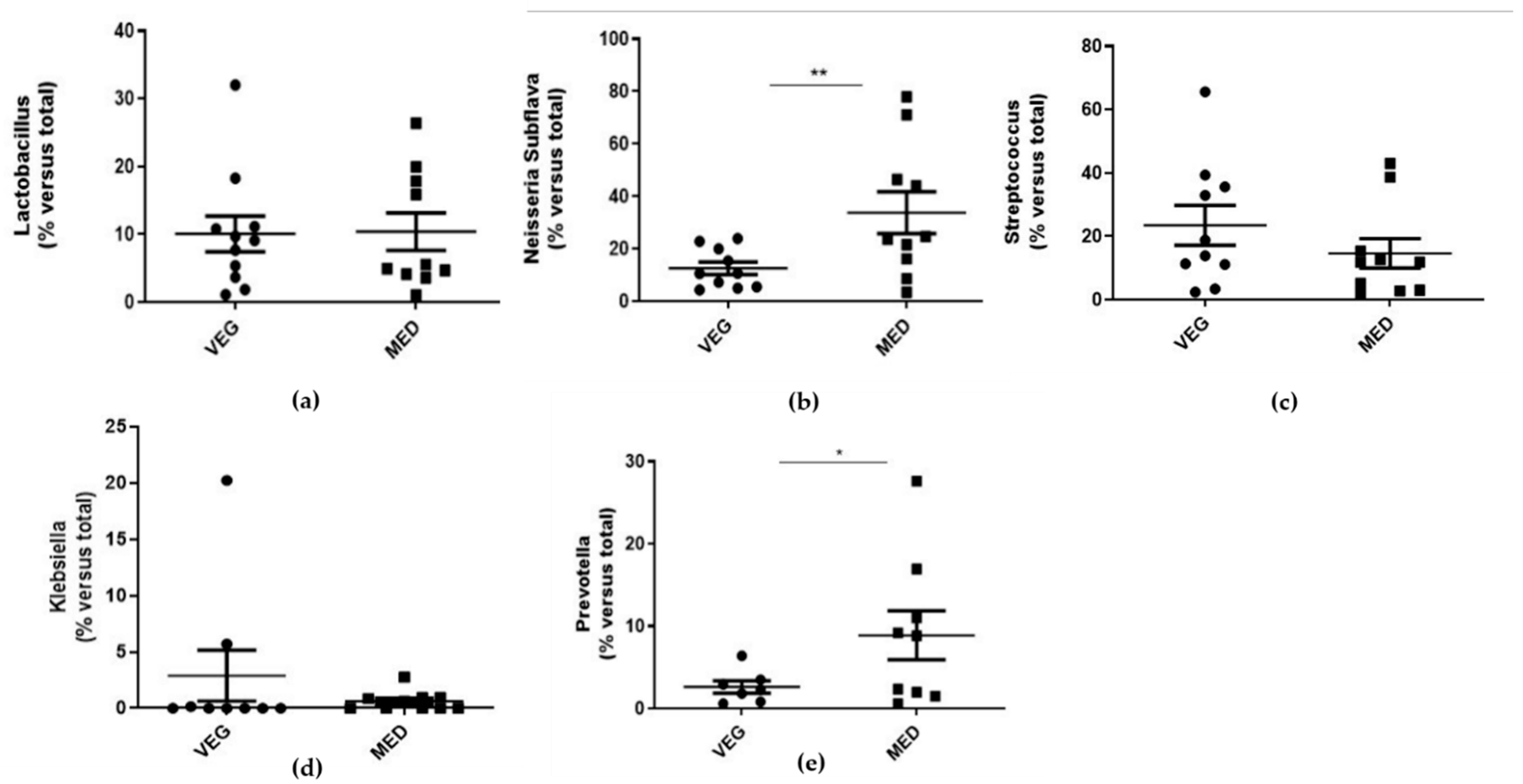
3.3. Oral Microbiota Analysis
3.4. Correlation between Metabolic and Microbiota Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kodukula, K.; Faller, D.V.; Harpp, D.N.; Kanara, I.; Pernokas, J.; Pernokas, M.; Powers, W.R.; Soukos, N.S.; Steliou, K.; Moos, W.H. Gut Microbiota and Salivary Diagnostics: The Mouth Is Salivating to Tell Us Something. BioResearch Open Access 2017, 6, 123–132. [Google Scholar] [CrossRef]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Bibbò, S.; Ianiro, G.; Giorgio, V.; Scaldaferri, F.; Masucci, L.; Gasbarrini, A.; Cammarota, G. The role of diet on gut microbiota composition. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4742–4749. [Google Scholar]
- Tomasello, G.; Mazzola, M.; Leone, A.; Sinagra, E.; Zummo, G.; Farina, F.; Damiani, P.; Cappello, F.; Geagea, A.G.; Jurjus, A.; et al. Nutrition, oxidative stress and intestinal dysbiosis: Influence of diet on gut microbiota in inflammatory bowel diseases. Biomed. Pap. 2016, 160, 461–466. [Google Scholar] [CrossRef] [Green Version]
- Sakkas, H.; Bozidis, P.; Touzios, C.; Kolios, D.; Athanasiou, G.; Athanasopoulou, E.; Gerou, I.; Gartzonika, C. Nutritional Status and the Influence of the Vegan Diet on the Gut Microbiota and Human Health. Medicina 2020, 56, 88. [Google Scholar] [CrossRef] [Green Version]
- Tomova, A.; Bukovsky, I.; Rembert, E.; Yonas, W.; Alwarith, J.; Barnard, N.D.; Kahleova, H. The Effects of Vegetarian and Vegan Diets on Gut Microbiota. Front. Nutr. 2019, 6, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belstrøm, D. The salivary microbiota in health and disease. J. Oral Microbiol. 2020, 12, 1723975. [Google Scholar] [CrossRef] [Green Version]
- Marsh, P.D.; Do, T.; Beighton, D.; Devine, D.A. Influence of saliva on the oral microbiota. Periodontol. 2000 2016, 70, 80–92. [Google Scholar] [CrossRef]
- Oba, P.; Holscher, H.; Mathai, R.; Kim, J.; Swanson, K. Diet Influences the Oral Microbiota of Infants during the First Six Months of Life. Nutrients 2020, 12, 3400. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.H.; Kern, T.; Bak, E.G.; Kashani, A.; Allin, K.; Nielsen, T.; Hansen, T.; Pedersen, O. Impact of a vegan diet on the human salivary microbiota. Sci. Rep. 2018, 8, 5847. [Google Scholar] [CrossRef]
- De Filippis, F.; Vannini, L.; La Storia, A.; Laghi, L.; Piombino, P.; Stellato, G.; Serrazanetti, D.I.; Gozzi, G.; Turroni, S.; Ferrocino, I.; et al. The Same Microbiota and a Potentially Discriminant Metabolome in the Saliva of Omnivore, Ovo-Lacto-Vegetarian and Vegan Individuals. PLoS ONE 2014, 9, e112373. [Google Scholar] [CrossRef] [Green Version]
- Bi, X.; Forde, C.G.; Goh, A.T.; Henry, C.J. Basal Metabolic Rate and Body Composition Predict Habitual Food and Macronutrient Intakes: Gender Differences. Nutrients 2019, 11, 2653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarpellini, E.; Arts, J.; Karamanolis, G.; Laurenius, A.; Siquini, W.; Suzuki, H.; Ukleja, A.; Van Beek, A.; Vanuytsel, T.; Bor, S.; et al. International consensus on the diagnosis and management of dumping syndrome. Nat. Rev. Endocrinol. 2020, 16, 448–466. [Google Scholar] [CrossRef]
- Moradi, S.; Pasdar, Y.; Hamzeh, B.; Najafi, F.; Nachvak, S.M.; Mostafai, R.; Niazi, P.; Rezaei, M. Comparison of 3 Nutritional Questionnaires to Determine Energy Intake Accuracy in Iranian Adults. Clin. Nutr. Res. 2018, 7, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Regulation of basal metabolic rate in uncomplicated pregnancy and in gestational diabetes mellitus. Hormones 2017, 16, 235–250. [CrossRef]
- Luy, S.C.; Center, Q.C.S.L.M.; Dampil, O.A. Comparison of the Harris-Benedict Equation, Bioelectrical Impedance Analysis, and Indirect Calorimetry for Measurement of Basal Metabolic Rate among Adult Obese Filipino Patients with Prediabetes or Type 2 Diabetes Mellitus. J. ASEAN Fed. Endocr. Soc. 2018, 33, 152–159. [Google Scholar] [CrossRef] [Green Version]
- A Yu, E.; Le, N.-A.; Stein, A.D. Measuring Postprandial Metabolic Flexibility to Assess Metabolic Health and Disease. J. Nutr. 2021, 151, 3284–3291. [Google Scholar] [CrossRef]
- Wang, T.; Lu, J.; Shi, L.; Chen, G.; Xu, M.; Xu, Y.; Su, Q.; Mu, Y.; Chen, L.; Hu, R.; et al. Association of insulin resistance and β-cell dysfunction with incident diabetes among adults in China: A nationwide, population-based, prospective cohort study. Lancet Diabetes Endocrinol. 2020, 8, 115–124. [Google Scholar] [CrossRef]
- Nadimi, H.; Yousefinejad, A.; Djazayery, A.; Hosseini, M.; Hosseini, S. Association of vegan diet with RMR, body composition and oxidative stress. Acta Sci. Pol. Technol. Aliment. 2013, 12, 311–318. [Google Scholar]
- Muñoz-Garcia, A.; Williams, J.B. Basal Metabolic Rate in Carnivores Is Associated with Diet after Controlling for Phylogeny. Physiol. Biochem. Zoo 2005, 78, 1039–1056. [Google Scholar] [CrossRef] [Green Version]
- Fallon, N.; Dillon, S.A. Low Intakes of Iodine and Selenium Amongst Vegan and Vegetarian Women Highlight a Potential Nutritional Vulnerability. Front. Nutr. 2020, 7, 72. [Google Scholar] [CrossRef]
- Broeders, E.P.M.; Vijgen, G.H.E.J.; Havekes, B.; Bouvy, N.D.; Mottaghy, F.M.; Kars, M.; Schaper, N.; Schrauwen, P.; Brans, B.; Lichtenbelt, W.D.V.M. Thyroid Hormone Activates Brown Adipose Tissue and Increases Non-Shivering Thermogenesis—A Cohort Study in a Group of Thyroid Carcinoma Patients. PLoS ONE 2016, 11, e0145049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, A.C.; Hyatt, T.C.; Hunter, G.R.; Gower, B.A. Respiratory Quotient Predicts Fat Mass Gain in Premenopausal Women. Obesity 2010, 18, 2255–2259. [Google Scholar] [CrossRef]
- Toubro, S.; Sørensen, T.I.A.; Hindsberger, C.; Christensen, N.J.; Astrup, A. Twenty-Four-Hour Respiratory Quotient: The Role of Diet and Familial Resemblance1. J. Clin. Endocrinol. Metab. 1998, 83, 2758–2764. [Google Scholar] [CrossRef] [Green Version]
- Péronnet, F.; Haman, F. Low capacity to oxidize fat and body weight. Obes. Rev. 2019, 20, 1367–13839. [Google Scholar] [CrossRef] [PubMed]
- Mentella, M.C.; Scaldaferri, F.; Ricci, C.; Gasbarrini, A.; Miggiano, G.A.D. Cancer and Mediterranean Diet: A Review. Nutrients 2019, 11, 2059. [Google Scholar] [CrossRef] [Green Version]
- Tuttolomondo, A.; Simonetta, I.; Daidone, M.; Mogavero, A.; Ortello, A.; Pinto, A. Metabolic and Vascular Effect of the Mediterranean Diet. Int. J. Mol. Sci. 2019, 20, 4716. [Google Scholar] [CrossRef] [Green Version]
- Ashworth, A.; Cutler, C.; Farnham, G.; Liddle, L.; Burleigh, M.C.; Rodiles, A.; Sillitti, C.; Kiernan, M.; Moore, M.; Hickson, M.; et al. Dietary intake of inorganic nitrate in vegetarians and omnivores and its impact on blood pressure, resting metabolic rate and the oral microbiome. Free Radic. Biol. Med. 2019, 138, 63–72. [Google Scholar] [CrossRef]
- Morse, S.A.; Stein, S.; Hines, J. Glucose Metabolism in Neisseria gonorrhoeae. J. Bacteriol. 1974, 120, 702–714. [Google Scholar] [CrossRef] [Green Version]
- Kahleova, H.; Rembert, E.; Alwarith, J.; Yonas, W.N.; Tura, A.; Holubkov, R.; Agnello, M.; Chutkan, R.; Barnard, N.D. Effects of a Low-Fat Vegan Diet on Gut Microbiota in Overweight Individuals and Relationships with Body Weight, Body Composition, and Insulin Sensitivity. A Randomized Clinical Trial. Nutrients 2020, 12, 2917. [Google Scholar] [CrossRef]
- Van Dam, R.; Seidell, J. Carbohydrate intake and obesity. Eur. J. Clin. Nutr. 2007, 61, S75–S99. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyotylainen, T.; Nielsen, T.; Jensen, B.A.H.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature 2016, 535, 376–381. [Google Scholar] [CrossRef]
- Indiani, C.M.D.S.P.; Rizzardi, K.F.; Castelo, P.M.; Ferraz, L.F.C.; Darrieux, M.; Parisotto, T. Childhood Obesity and Firmicutes/Bacteroidetes Ratio in the Gut Microbiota: A Systematic Review. Child. Obes. 2018, 14, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Dhakal, S.; McCormack, L.; Dey, M. Association of the Gut Microbiota with Weight-Loss Response within a Retail Weight-Management Program. Microorganisms 2020, 8, 1246. [Google Scholar] [CrossRef] [PubMed]
- Zijnge, V.; Van Leeuwen, M.B.M.; Degener, J.E.; Abbas, F.; Thurnheer, T.; Gmuer, R.; Harmsen, H.J.M. Oral Biofilm Architecture on Natural Teeth. PLoS ONE 2010, 5, e9321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Tang, Y.; Hou, X.; Wang, H.; Huang, L.; Wen, J.; Niu, H.; Zeng, W.; Bai, Y. Novel Lactobacillus reuteri HI120 Affects Lipid Metabolism in C57BL/6 Obese Mice. Front. Veter. Sci. 2020, 7, 560241. [Google Scholar] [CrossRef] [PubMed]
- Buron-Moles, G.; Chailyan, A.; Dolejs, I.; Forster, J.; Mikš, M.H. Uncovering carbohydrate metabolism through a genotype-phenotype association study of 56 lactic acid bacteria genomes. Appl. Microbiol. Biotechnol. 2019, 103, 3135–3152. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| Reverse | Forward | T Annealing | |
|---|---|---|---|
| 16S | AGGGTTGCGCTCGTTG | GTGCCAGCAGCCGCGGTAA | 64 °C |
| All Lactobacillus | CCACCTTCCTCCGGTTTGTCA | AGGGTGAAGTCGTAACAAGTAGCC | 60 °C |
| Nesseria subflava | TGGAAGACGGATTTGGTGTAAT | CCAACGATGTTGCCGAATTG | 58 °C |
| Streptococcus pneumoniae | GTACAGTTGCTTCAGGACGTATC | ACGTTCGATTTCATCACGTTG | 55 °C |
| Prevotella | GTGGCGCGTATTTTATGTATGTG | ATCCGCCATACGCCCTTAG | 60 °C |
| Group | Age | Weight | Height | BMI | BMR Harris– Benedict | BMR | RQ | CHO% | Lipids% |
|---|---|---|---|---|---|---|---|---|---|
| VEG (Mean ± SD) | 33.6 ± 12.3 | 60.6 ± 12.3 | 165.3 ± 7.2 | 22.1 ± 4.2 | 1425.5 ± 134.8 | 1233.6 ± 417.8 | 0.8 ± 123 | 48.08 ± 33.2 | 53.03 ± 33.9 |
| MED (Mean ± SD) | 31.7 ± 8.9 | 72.6 ± 16.9 | 174.1 ± 8.6 | 23.6 ± 3.5 | 1677.8 ± 295.7 ** | 1630.5 ± 566.9 * | 0.7 ± 0.037 * | 25.6 ± 16.3 ** | 74.7 ± 16.5 * |
| Group | Lactobacillus (%/Total) | Subflava (%/Total) | Streptococcus (%/Total) | Klebsiella (%/Total) | Prevotella (%/Total) |
|---|---|---|---|---|---|
| VEG (Mean ± SD) | 10.08 ± 8.5 | 12.6 ± 7.3 | 23.4 ± 19.2 | 2.7 ± 6.4 | 2.6 ± 1.8 |
| MED (Mean ± SD) | 10.4 ± 8.5 | 33.7 ± 24.7 ** | 14.6 ± 14.2 | 0.6 ± 0.8 | 8.8 ± 8.6 * |
| Total Population | VEG | MED | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Correlation | Z-Value | p-Value | Correlation | Z-Value | p-Value | Correlation | Z-Value | p-Value | |
| Lactobacillus, Streptococcus | 0.559 | 3.838 | 0.0001 | 0.803 | 4.563 | <0.0001 | 0.336 | 1.440 | 0.1500 |
| Lactobacillus, Subflava | −0.382 | −2.448 | 0.0144 | −0.188 | −0.786 | 0.4321 | −0.663 | −3.294 | 0.0010 |
| Lactobacillus, BMR Harris–Benedict | −0.120 | −0.751 | 0.4529 | 0.512 | 2.464 | 0.0137 | −0.499 | −2.261 | 0.0238 |
| Subflava, BMR Harris–Benedict | 0.647 | 4.679 | <0.0001 | 0.159 | 0.659 | 0.5096 | 0.592 | 2.808 | 0.0050 |
| Klebsiella, BMI | −0.375 | −2.362 | 0.0182 | −0.406 | −1.725 | 0.0846 | −0.433 | −1.909 | 0.0563 |
| Klebsiella, CHO% | 0.312 | 1.939 | 0.0525 | 0.287 | 1.179 | 0.2382 | −0.207 | −0.866 | 0.3863 |
| Klebsiella, Lipids% | −0.315 | −1.954 | 0.0507 | −0.291 | −1.198 | 0.2309 | 0.194 | 0.808 | 0.4190 |
| Prevotella, CHO% | −0.359 | −2.024 | 0.0430 | −0.609 | −2.347 | 0.0189 | −0.335 | −1.348 | 0.1777 |
| Prevotella, BMI | −0.099 | −0.537 | 0.5913 | 0.792 | 3.574 | 0.0004 | −0.539 | −2.336 | 0.0195 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daniele, S.; Scarfò, G.; Ceccarelli, L.; Fusi, J.; Zappelli, E.; Biagini, D.; Lomonaco, T.; Di Francesco, F.; Franzoni, F.; Martini, C. The Mediterranean Diet Positively Affects Resting Metabolic Rate and Salivary Microbiota in Human Subjects: A Comparison with the Vegan Regimen. Biology 2021, 10, 1292. https://doi.org/10.3390/biology10121292
Daniele S, Scarfò G, Ceccarelli L, Fusi J, Zappelli E, Biagini D, Lomonaco T, Di Francesco F, Franzoni F, Martini C. The Mediterranean Diet Positively Affects Resting Metabolic Rate and Salivary Microbiota in Human Subjects: A Comparison with the Vegan Regimen. Biology. 2021; 10(12):1292. https://doi.org/10.3390/biology10121292
Chicago/Turabian StyleDaniele, Simona, Giorgia Scarfò, Lorenzo Ceccarelli, Jonathan Fusi, Elisa Zappelli, Denise Biagini, Tommaso Lomonaco, Fabio Di Francesco, Ferdinando Franzoni, and Claudia Martini. 2021. "The Mediterranean Diet Positively Affects Resting Metabolic Rate and Salivary Microbiota in Human Subjects: A Comparison with the Vegan Regimen" Biology 10, no. 12: 1292. https://doi.org/10.3390/biology10121292
APA StyleDaniele, S., Scarfò, G., Ceccarelli, L., Fusi, J., Zappelli, E., Biagini, D., Lomonaco, T., Di Francesco, F., Franzoni, F., & Martini, C. (2021). The Mediterranean Diet Positively Affects Resting Metabolic Rate and Salivary Microbiota in Human Subjects: A Comparison with the Vegan Regimen. Biology, 10(12), 1292. https://doi.org/10.3390/biology10121292