
A Novel Vitronectin Peptide Facilitates Differentiation of Oligodendrocytes from Human Pluripotent Stem Cells (Synthetic ECM for Oligodendrocyte Differentiation)

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Designing New Peptide Motifs from Vitronectin
2.2. Production and Coating of Engineered Peptides
2.3. Culturing and Differentiation of Human ESCs
2.4. Immunofluorescent Staining
2.5. Isolation of RNA and Quantitative Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.6. Acquisition and Analysis of RNAseq Data
2.7. Statistical Analysis
3. Results
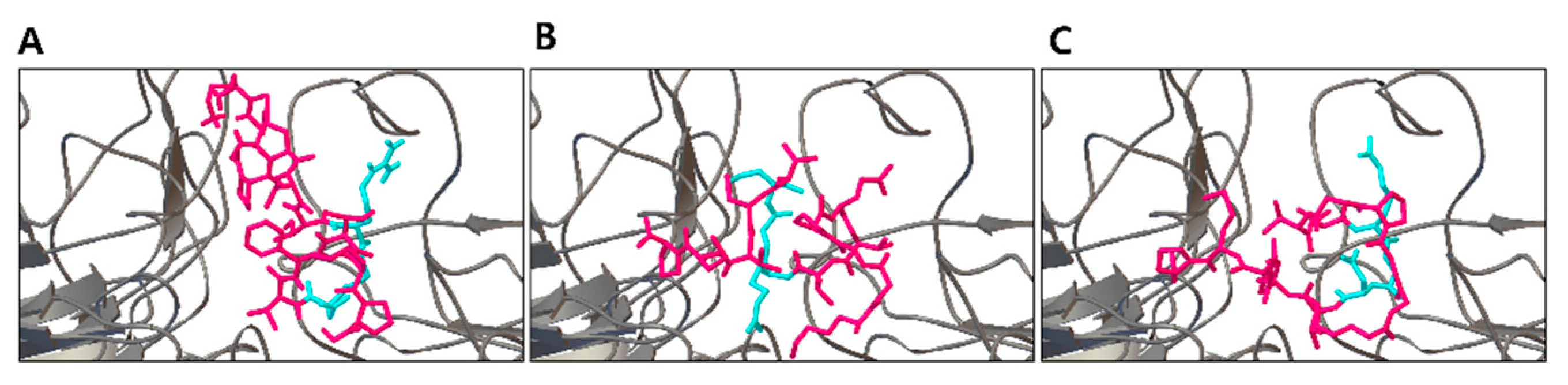
3.1. Prediction and Identification of Novel Peptide Motifs from VN
3.2. Differentiation of OPCs from hPSCs
3.3. Differentiation of OPCs into Oligodendrocytes through Forced Expression of SOX10
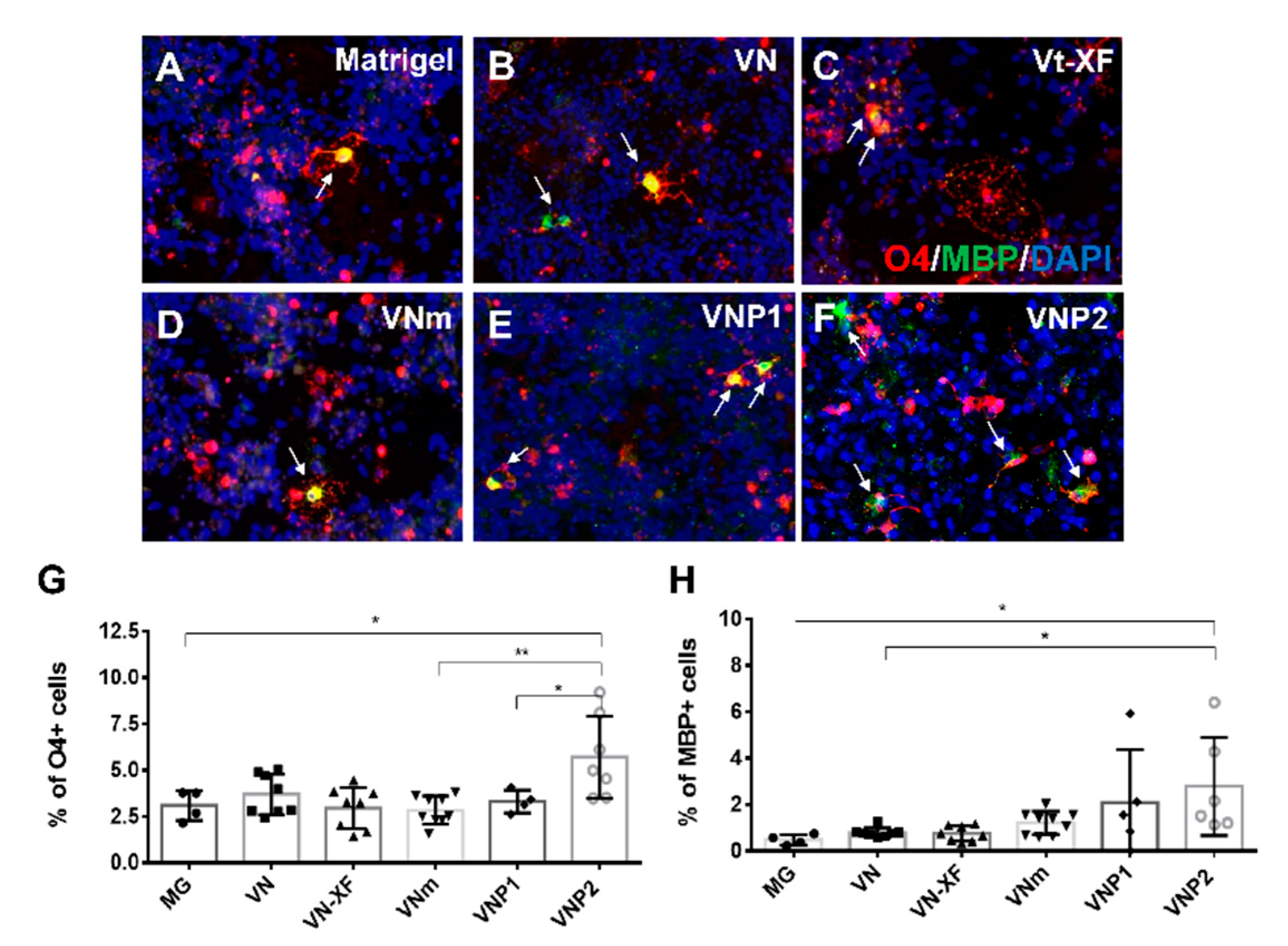
3.4. Screening of Various Substrates for Oligodendrocyte Differentiation
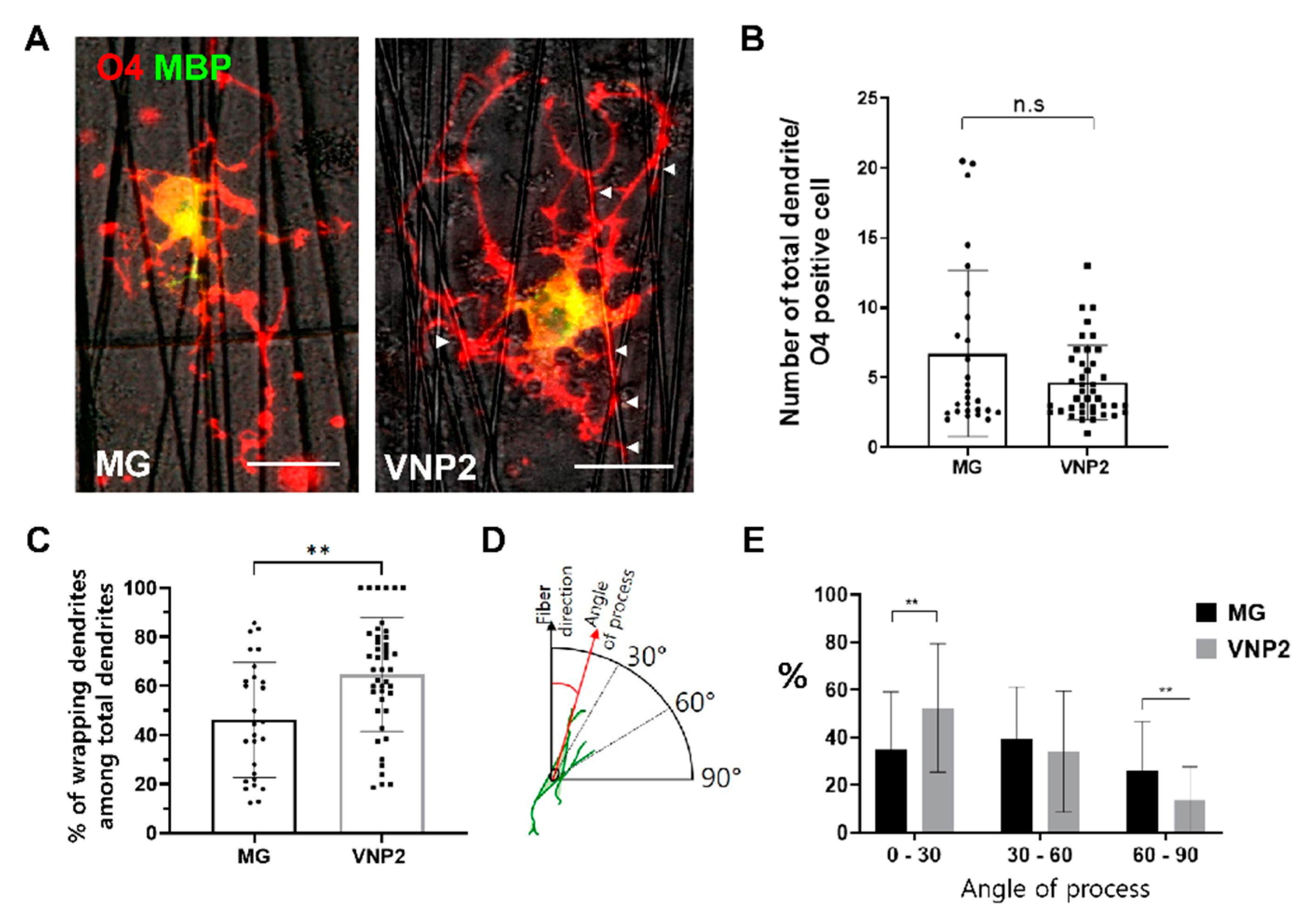
3.5. Differentiation of Oligodendrocytes on Nanofibers Coated with Matrigel or VNP2
3.6. Differential Gene Expression in OPCs Differentiated on Different Matrices
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barcelos, I.P.; Troxell, R.M.; Graves, J.S. Mitochondrial dysfunction and multiple sclerosis. Biology 2019, 8, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Knaap, M.S.; Bugiani, M. Leukodystrophies: A proposed classification system based on pathological changes and pathogenetic mechanisms. Acta Neuropathol. 2017, 134, 351–382. [Google Scholar] [CrossRef] [Green Version]
- Huntemer-Silveira, A.; Patil, N.; Brickner, M.A.; Parr, A.M. Strategies for oligodendrocyte and myelin repair in traumatic CNS injury. Front. Cell. Neurosci. 2020, 14, 619–707. [Google Scholar] [CrossRef]
- Zhou, Q.; Choi, G.; Anderson, D.J. The bHLH transcription factor Olig2 promotes oligodendrocyte differentiation in collaboration with Nkx2.2. Neuron 2001, 31, 791–807. [Google Scholar] [CrossRef] [Green Version]
- Stolt, C.C.; Rehberg, S.; Ader, M.; Lommes, P.; Riethmacher, D.; Schachner, M.; Bartsch, U.; Wegner, M. Terminal differentiation of myelin-forming oligodendrocytes depends on the transcription factor Sox10. Genes Dev. 2002, 16, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Emery, B.; Lu, Q.R. Transcriptional and epigenetic regulation of oligodendrocyte development and myelination in the central nervous system. Cold Spring Harb. Perspect. Biol. 2015, 7, a020461. [Google Scholar] [CrossRef] [Green Version]
- Michalski, D.; Keck, A.L.; Grosche, J.; Martens, H.; Hartig, W. Immunosignals of oligodendrocyte markers and myelin-associated proteins are critically affected after experimental stroke in wild-type and alzheimer modeling mice of different ages. Front. Cell. Neurosci. 2018, 12, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinosa-Hoyos, D.; Burstein, S.R.; Cha, J.; Jain, T.; Nijsure, M.; Jagielska, A.; Fossati, V.; Van Vliet, K.J. Mechanosensitivity of Human Oligodendrocytes. Front. Cell. Neurosci. 2020, 14, 222. [Google Scholar] [CrossRef]
- Niklason, L.E. Understanding the extracellular matrix to enhance stem cell-based tissue regeneration. Cell Stem Cell 2018, 22, 302–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardwell, M.C.; Rome, L.H. Evidence that an RGD-dependent receptor mediates the binding of oligodendrocytes to a novel ligand in a glial-derived matrix. J. Cell Biol. 1988, 107, 1541–1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil, J.E.; Woo, D.H.; Shim, J.H.; Kim, S.E.; You, H.J.; Park, S.H.; Paek, S.H.; Kim, S.K.; Kim, J.H. Vitronectin promotes oligodendrocyte differentiation during neurogenesis of human embryonic stem cells. FEBS Lett. 2009, 583, 561–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Gautam, A.; Yang, J.; Qiu, L.; Melkoumian, Z.; Weber, J.; Telukuntla, L.; Srivastava, R.; Whiteley, E.M.; Brandenberger, R. Differentiation of oligodendrocyte progenitor cells from human embryonic stem cells on vitronectin-derived synthetic peptide acrylate surface. Stem Cells Dev. 2013, 22, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Schymkowitz, J.; Borg, J.; Stricher, F.; Nys, R.; Rousseau, F.; Serrano, L. The FoldX web server: An online force field. Nucleic Acids Res. 2005, 33, W382–W388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dayem, A.A.; Won, J.; Goo, H.G.; Yang, G.M.; Seo, D.S.; Jeon, B.M.; Choi, H.Y.; Park, S.E.; Lim, K.M.; Jang, S.H.; et al. The immobilization of fibronectin- and fibroblast growth factor 2-derived peptides on a culture plate supports the attachment and proliferation of human pluripotent stem cells. Stem Cell Res. 2020, 43, 101700. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.S.; Sim, S.B.; Cha, H.J. Cell adhesion biomaterial based on mussel adhesive protein fused with RGD peptide. Biomaterials 2007, 28, 4039–4046. [Google Scholar] [CrossRef]
- Melkoumian, Z.; Weber, J.L.; Weber, D.M.; Fadeev, A.G.; Zhou, Y.; Dolley-Sonneville, P.; Yang, J.; Qiu, L.; Priest, C.A.; Shogbon, C.; et al. Synthetic peptide-acrylate surfaces for long-term self-renewal and cardiomyocyte differentiation of human embryonic stem cells. Nat. Biotechnol. 2010, 28, 606–610. [Google Scholar] [CrossRef] [PubMed]
- Yeon, G.; Shin, W.; Yoo, S.H.; Kim, D.; Jeon, B.; Park, W.; Bae, Y.; Park, J.; You, S.; Na, D.K.; et al. NF1B induces functional astrocytes from human pluripotent stem cell-derived neural precursor cells mimicking in vivo astroglioenesis. J. Cell. Physiol. 2021, 236, 7625–7641. [Google Scholar] [CrossRef]
- Douvaras, P.; Fossati, V. Generation and isolation of oligodendrocyte progenitor cells from human pluripotent stem cells. Nat. Protoc. 2015, 10, 1143–1154. [Google Scholar] [CrossRef] [PubMed]
- Krueger, F. Trim Galore!: A Wrapper Tool around Cutadapt and FastQC to Consistently Apply Quality and Adapter Trimming to FastQ Files. 2015. Available online: https://github.com/FelixKrueger/TrimGalore (accessed on 1 May 2020).
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Soneson, C.; Love, M.I.; Robinson, M.D. Differential analyses for RNA-seq: Transcript-level estimates improve gene-level inferences. F1000Research 2015, 4, 1521. [Google Scholar] [CrossRef] [PubMed]
- Pantano, L. R package version. In DEGreport: Report of DEG Analysis; Lorena Pantano Harvard TH Chan School of Public Health: Boston, MA, USA, 2019; p. 1. [Google Scholar]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takebayashi, H.; Nabeshima, Y.; Yoshida, S.; Chisaka, O.; Ikenaka, K.; Nabeshima, Y. The basic helix-loop-helix factor olig2 is essential for the development of motoneuron and oligodendrocyte lineages. Curr. Biol. 2002, 12, 1157–1163. [Google Scholar] [CrossRef] [Green Version]
- Ono, K.; Takebayashi, H.; Ikeda, K.; Furusho, M.; Nishizawa, T.; Watanabe, K.; Ikenaka, K. Regional- and temporal-dependent changes in the differentiation of Olig2 progenitors in the forebrain, and the impact on astrocyte development in the dorsal pallium. Dev. Biol. 2008, 320, 456–468. [Google Scholar] [CrossRef] [Green Version]
- Williamson, J.M.; Lyons, D.A. Myelin dynamics throughout life: An ever-changing landscape? Front. Cell. Neurosci. 2018, 12, 424. [Google Scholar] [CrossRef]
- Hu, B.Y.; Du, Z.W.; Li, X.J.; Ayala, M.; Zhang, S.C. Human oligodendrocytes from embryonic stem cells: Conserved SHH signaling networks and divergent FGF effects. Development 2009, 136, 1443–1452. [Google Scholar] [CrossRef] [Green Version]
- Douvaras, P.; Wang, J.; Zimmer, M.; Hanchuk, S.; O’Bara, M.A.; Sadiq, S.; Sim, F.J.; Goldman, J.; Fossati, V. Efficient generation of myelinating oligodendrocytes from primary progressive multiple sclerosis patients by induced pluripotent stem cells. Stem Cell Rep. 2014, 3, 250–259. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Leon, J.A.; Kumar, M.; Boon, R.; Chau, D.; One, J.; Wolfs, E.; Eggermont, K.; Berckmans, P.; Gunhanlar, N.; de Vrij, F.; et al. SOX10 single transcription factor-based fast and efficient generation of oligodendrocytes from human pluripotent stem cells. Stem Cell Rep. 2018, 10, 655–672. [Google Scholar] [CrossRef] [Green Version]
- Park, S.E.; Yeon, G.B.; Goo, H.G.; Seo, D.S.; Dayem, A.A.; Lee, K.E.; Park, H.M.; Cho, S.G.; Kim, D.S. Maintenance and differentiation of human ES cells on polyvinylidene fluoride scaffolds immobilized with a vitronectin-derived peptide. J. Cell Physiol. 2020, 236, 7291–7292. [Google Scholar] [CrossRef]
- Lee, S.; Leach, M.K.; Redmond, S.A.; Chong, S.Y.; Mellon, S.H.; Tuck, S.J.; Feng, Z.Q.; Corey, J.M.; Chan, J.R. A culture system to study oligodendrocyte myelination processes using engineered nanofibers. Nat. Methods 2012, 9, 917–922. [Google Scholar] [CrossRef] [Green Version]
- Spitzer, S.O.; Sitnikov, S.; Kamen, Y.; Evans, K.A.; Kronenberg-Versteeg, D.; Dietmann, S.; de Faria, O., Jr.; Agathou, S.; Karadottir, R.T. Oligodendrocyte progenitor cells become regionally diverse and heterogeneous with age. Neuron 2019, 101, 459–471.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walz, W. Chloride/anion channels in glial cell membranes. Glia 2002, 40, 1–10. [Google Scholar] [CrossRef]
- Gautier, H.O.; Evans, K.A.; Volbracht, K.; James, R.; Sitnikov, S.; Lundgaard, I.; James, F.; Lao-Peregrin, C.; Reynolds, R.; Franklin, R.J.; et al. Neuronal activity regulates remyelination via glutamate signalling to oligodendrocyte progenitors. Nat. Commun. 2015, 6, 8518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bansal, R.; Kumar, M.; Murray, K.; Morrison, R.S.; Pfeiffer, S.E. Regulation of FGF receptors in the oligodendrocyte lineage. Mol. Cell. Neurosci. 1996, 7, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.B.; Zhang, Z.; Li, J.N.; Yang, T.; Du, S.; Cao, R.J.; Cui, S.S. SPP1 promotes Schwann cell proliferation and survival through PKCalpha by binding with CD44 and alphavbeta3 after peripheral nerve injury. Cell Biosci. 2020, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, R.S.; Stevens, S.; Wing, M.G.; Compston, D.A. Microglia-derived IGF-2 prevents TNFalpha induced death of mature oligodendrocytes in vitro. J. Neuroimmunol. 2002, 124, 36–44. [Google Scholar] [CrossRef]
- Dent, M.A.; Sumi, Y.; Morris, R.J.; Seeley, P.J. Urokinase-type plasminogen activator expression by neurons and oligodendrocytes during process outgrowth in developing rat brain. Eur. J. Neurosci. 1993, 5, 633–647. [Google Scholar] [CrossRef]
- Rigby, M.J.; Gomez, T.M.; Puglielli, L. Glial cell-axonal growth cone interactions in neurodevelopment and regeneration. Front. Neurosci. 2020, 14, 203. [Google Scholar] [CrossRef] [Green Version]
- Rothberg, J.M.; Jacobs, J.R.; Goodman, C.S.; Artavanis-Tsakonas, S. slit: An extracellular protein necessary for development of midline glia and commissural axon pathways contains both EGF and LRR domains. Genes Dev. 1990, 4, 2169–2187. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Mino, R.E.; Fisher, E.S.; Bhat, M.A. A versatile genetic tool to study midline glia function in the Drosophila CNS. Dev. Biol. 2017, 429, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Celiz, A.D.; Smith, J.G.; Langer, R.; Anderson, D.G.; Winkler, D.A.; Barrett, D.A.; Davies, M.C.; Young, L.E.; Denning, C.; Alexander, M.R. Materials for stem cell factories of the future. Nat. Mater. 2014, 13, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Klim, J.R.; Li, L.; Wrighton, P.J.; Piekarczyk, M.S.; Kiessling, L.L. A defined glycosaminoglycan-binding substratum for human pluripotent stem cells. Nat. Methods 2010, 7, 989–994. [Google Scholar] [CrossRef]
- Tan, Y.; Tian, T.; Liu, W.; Zhu, Z.; Yang, C.J. Advance in phage display technology for bioanalysis. Biotechnol. J. 2016, 11, 732–745. [Google Scholar] [CrossRef]
- Ferreira, L.G.; Dos Santos, R.N.; Oliva, G.; Andricopulo, A.D. Molecular docking and structure-based drug design strategies. Molecules 2015, 20, 13384–13421. [Google Scholar] [CrossRef]
- Lee, B.Y.; Hur, E.M. A role of microtubules in oligodendrocyte differentiation. Int. J. Mol. Sci. 2020, 21, 1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, N.G.; Richter-Landsberg, C.; Ffrench-Constant, C. Role of the oligodendroglial cytoskeleton in differentiation and myelination. Glia 2009, 57, 1691–1705. [Google Scholar] [CrossRef]
- Terada, N.; Kidd, G.J.; Kinter, M.; Bjartmar, C.; Moran-Jones, K.; Trapp, B.D. Beta IV tubulin is selectively expressed by oligodendrocytes in the central nervous system. Glia 2005, 50, 212–222. [Google Scholar] [CrossRef]
- Tonduti, D.; Aiello, C.; Renaldo, F.; Dorboz, I.; Saaman, S.; Rodriguez, D.; Fettah, H.; Elmaleh, M.; Biancheri, R.; Barresi, S.; et al. TUBB4A-related hypomyelinating leukodystrophy: New insights from a series of 12 patients. Eur. J. Paediatr. Neurol. 2016, 20, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Bercury, K.K.; Ahrendsen, J.T.; Macklin, W.B. Olig1 function is required for oligodendrocyte differentiation in the mouse brain. J. Neurosci. 2015, 35, 4386–4402. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, W.U.; Yeon, G.-B.; Yu, M.-S.; Goo, H.-G.; Hwang, S.-H.; Na, D.; Kim, D.-S. A Novel Vitronectin Peptide Facilitates Differentiation of Oligodendrocytes from Human Pluripotent Stem Cells (Synthetic ECM for Oligodendrocyte Differentiation). Biology 2021, 10, 1254. https://doi.org/10.3390/biology10121254
Park WU, Yeon G-B, Yu M-S, Goo H-G, Hwang S-H, Na D, Kim D-S. A Novel Vitronectin Peptide Facilitates Differentiation of Oligodendrocytes from Human Pluripotent Stem Cells (Synthetic ECM for Oligodendrocyte Differentiation). Biology. 2021; 10(12):1254. https://doi.org/10.3390/biology10121254
Chicago/Turabian StylePark, Won Ung, Gyu-Bum Yeon, Myeong-Sang Yu, Hui-Gwan Goo, Su-Hee Hwang, Dokyun Na, and Dae-Sung Kim. 2021. "A Novel Vitronectin Peptide Facilitates Differentiation of Oligodendrocytes from Human Pluripotent Stem Cells (Synthetic ECM for Oligodendrocyte Differentiation)" Biology 10, no. 12: 1254. https://doi.org/10.3390/biology10121254
APA StylePark, W. U., Yeon, G.-B., Yu, M.-S., Goo, H.-G., Hwang, S.-H., Na, D., & Kim, D.-S. (2021). A Novel Vitronectin Peptide Facilitates Differentiation of Oligodendrocytes from Human Pluripotent Stem Cells (Synthetic ECM for Oligodendrocyte Differentiation). Biology, 10(12), 1254. https://doi.org/10.3390/biology10121254