
Circadian Gene PER2 Silencing Downregulates PPARG and SREBF1 and Suppresses Lipid Synthesis in Bovine Mammary Epithelial Cells

 ,
,  ,
, 

Abstract
:Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. The Isolation and Culture of Primary BMECs
2.2. Cell Transfection
2.3. RNA Extraction and RT-PCR
2.4. The Cell Proliferation Activity, Cell Cycles, and Apoptosis Assays
2.5. Triacylglycerol Content and Lipid Droplet Determination
2.6. Statistical Analyses
3. Results
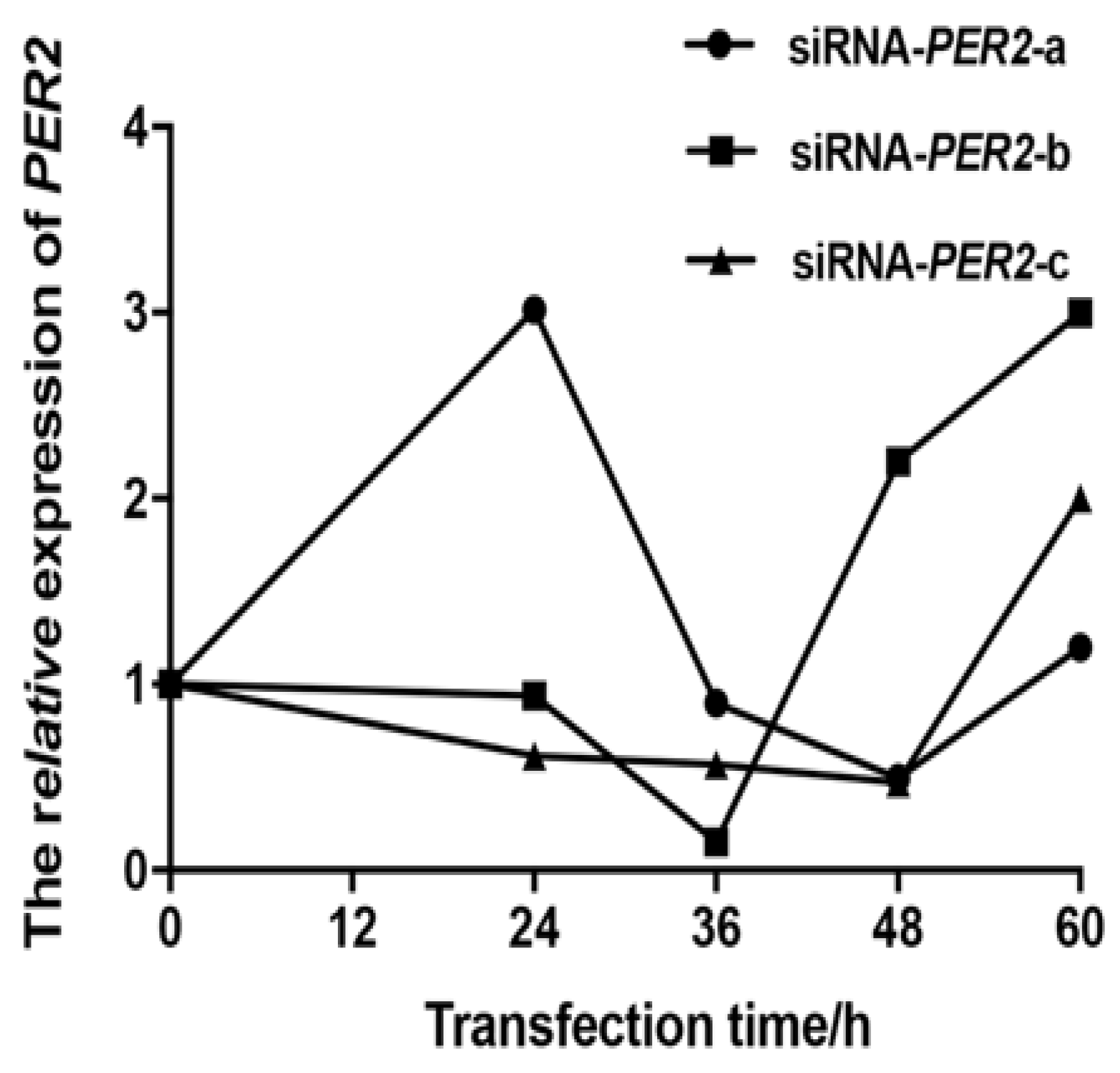
3.1. Screening for Transfection Efficiency
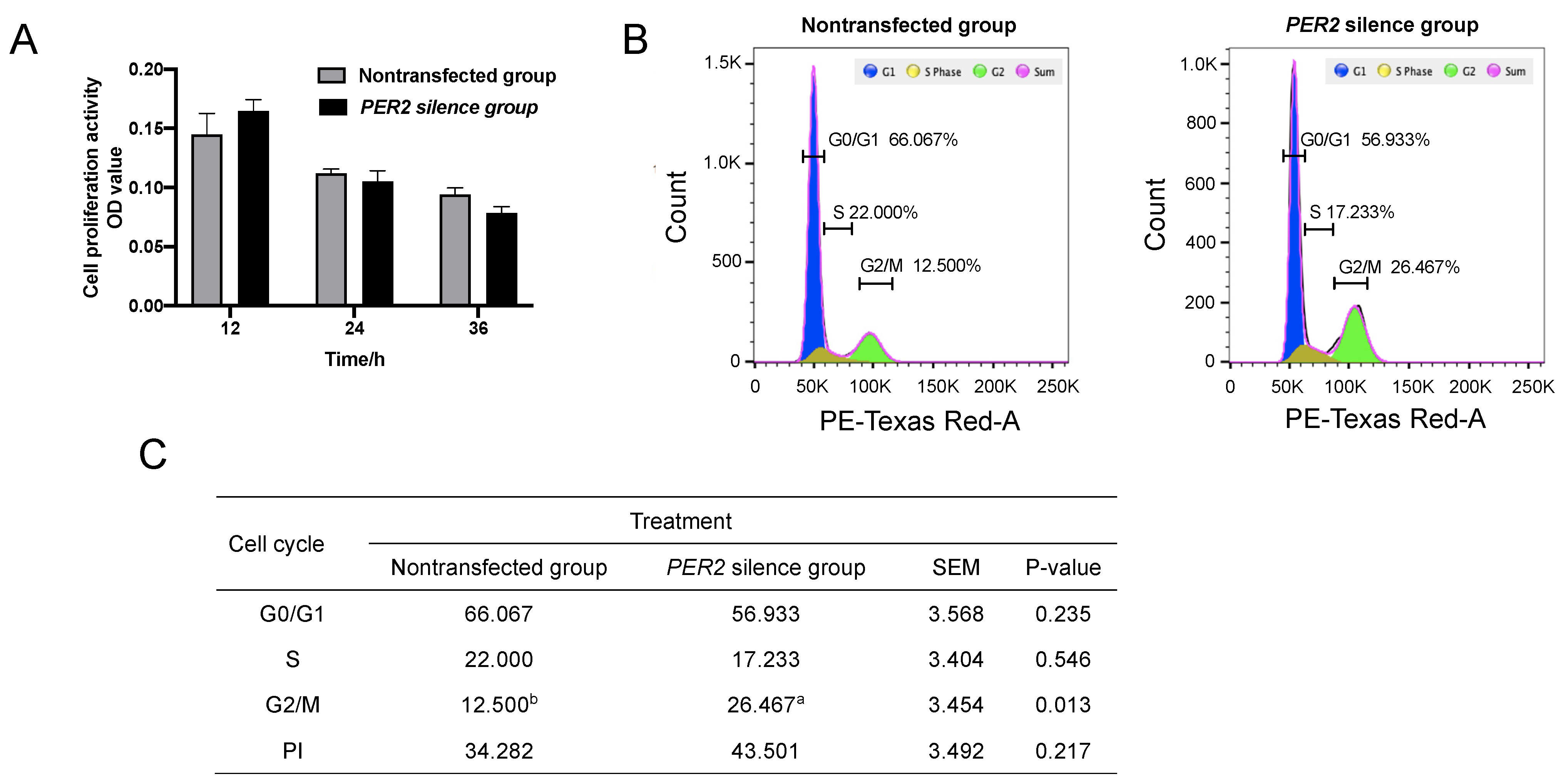
3.2. The Cell Proliferation Activity and Cell Cycles
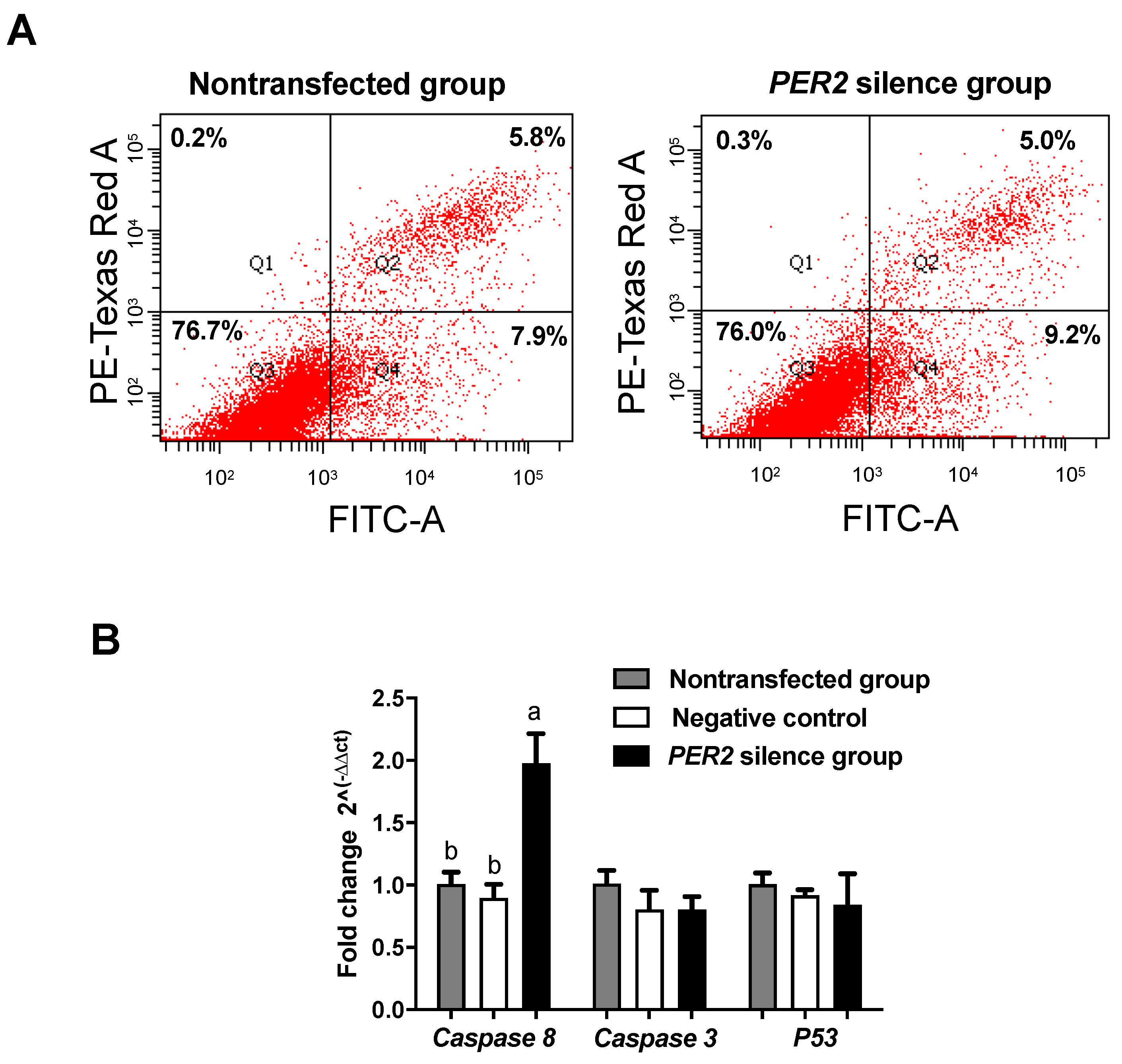
3.3. Cell Apoptosis
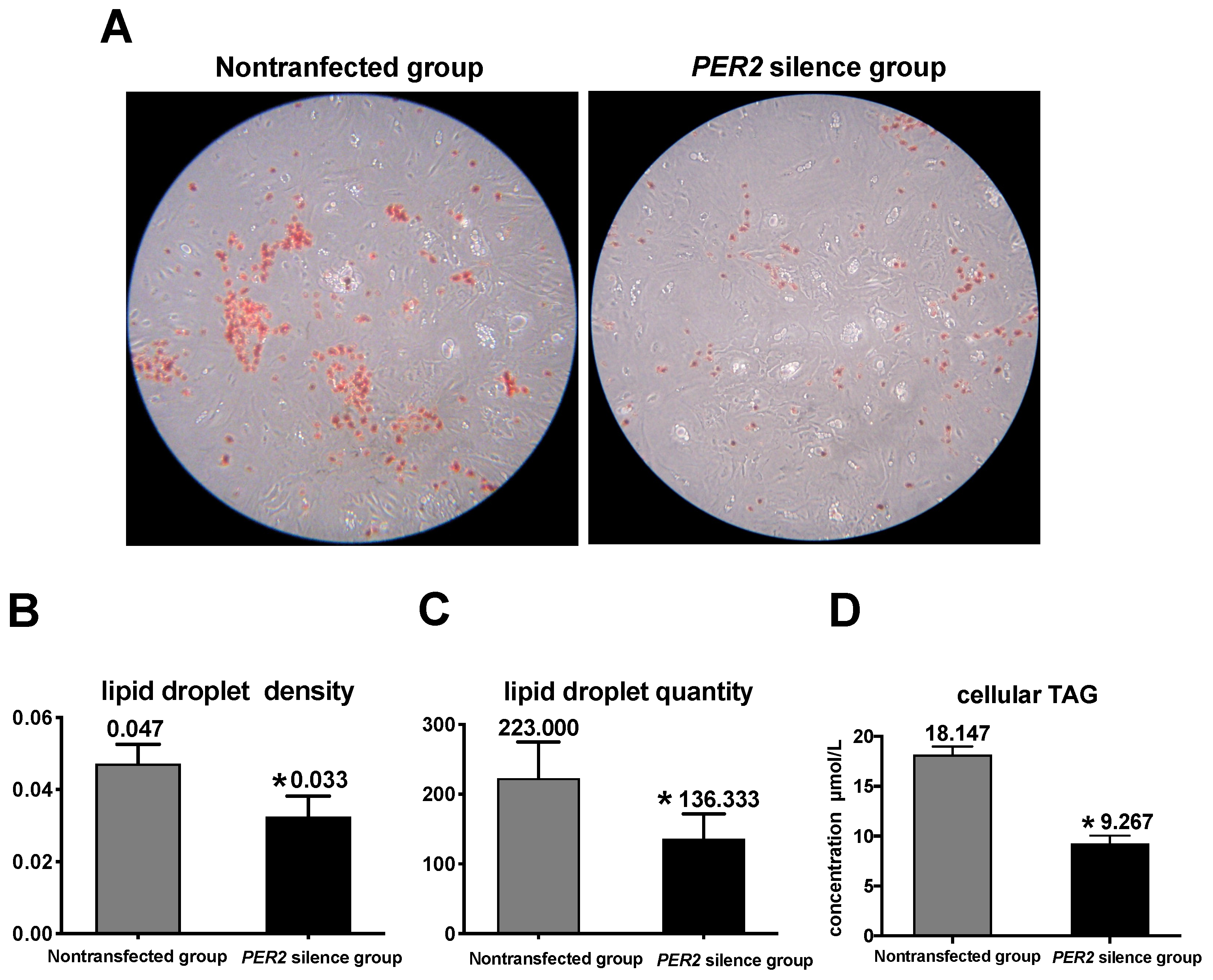
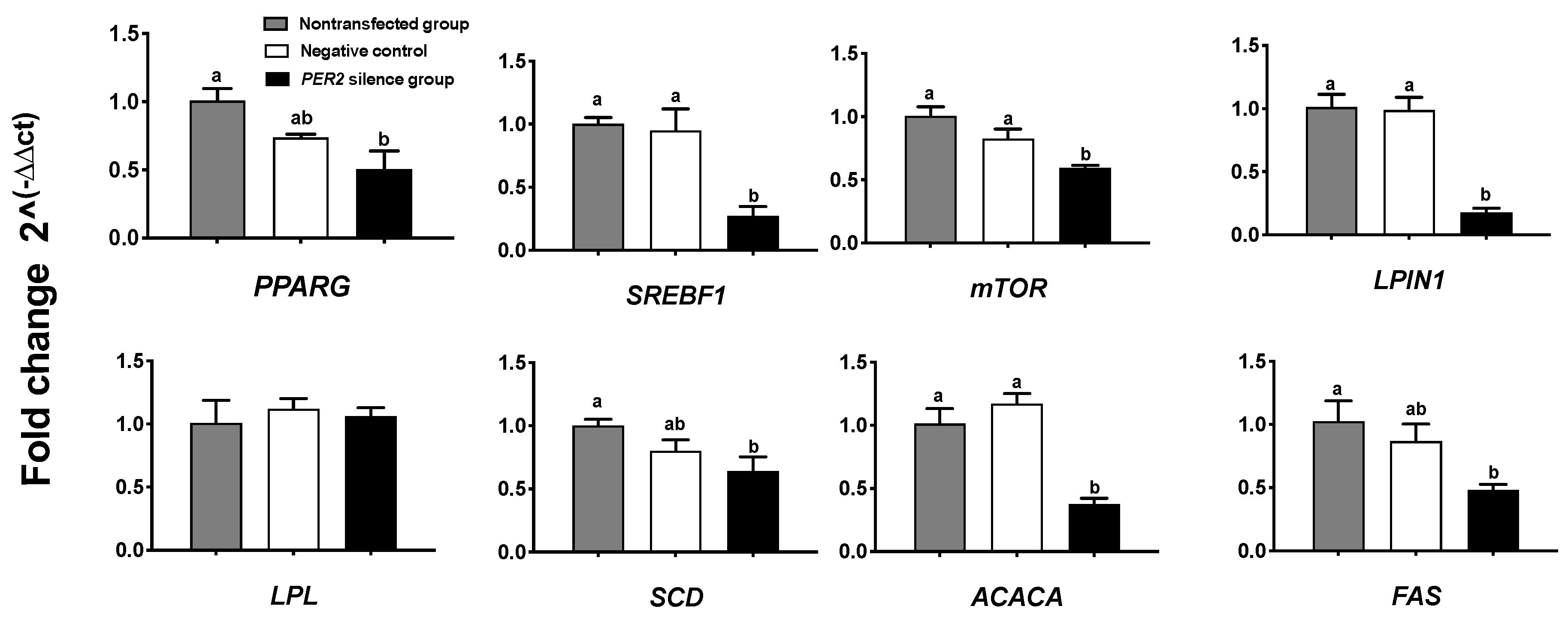
3.4. Lipid Synthesis
4. Discussion
4.1. PER2 Silencing Downregulates Mammary Lipogenic Genes
4.2. PER2 Silencing Suppresses Cellular TAG Accumulation and Lipid Droplet Formation
4.3. PER2 Silencing Regulates the Cell Cycle, but Not Apoptosis
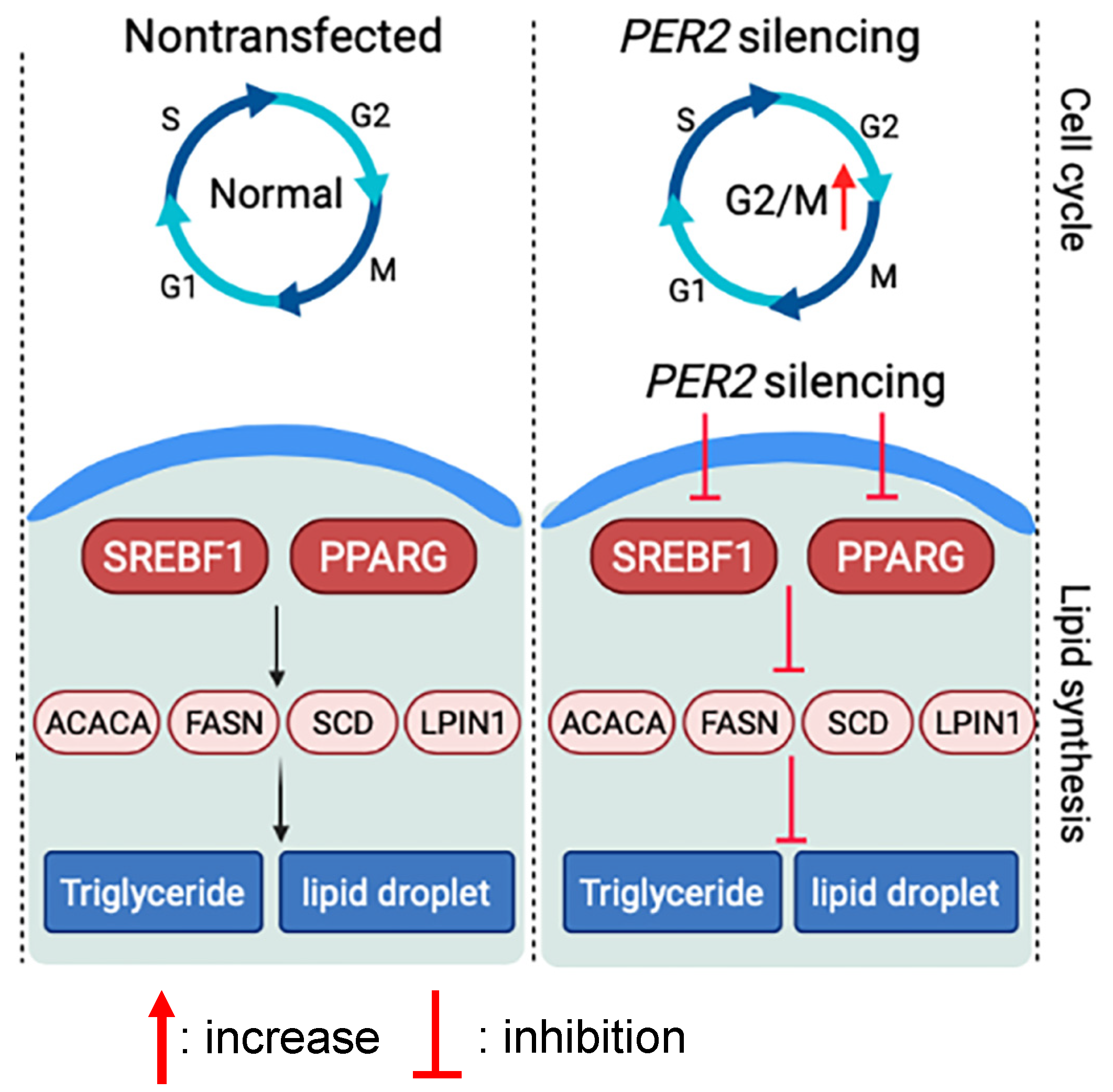
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Canaple, L.; Kakizawa, T.; Laudet, V. The days and nights of cancer cells. Cancer Res. 2003, 63, 7545–7552. [Google Scholar] [PubMed]
- Xiang, S.; Mao, L.; Duplessis, T.; Dauchy, R.; Dauchy, E.; Blask, D.E.; Frasch, T.; Hill, S.M. Oscillation of clock and clock controlled genes induced by serum shock in human breast epithelial and breast cancer cells: Regulation by melatonin. Breast Cancer-Basic 2012, 6, 137–150. [Google Scholar] [CrossRef]
- Deng, F.; Yang, K. Current status of research on the period family of clock genes in the occurrence and development of cancer. J. Cancer 2019, 10, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Casey, T.M.; Plaut, K. Lactation biology symposium: Circadian clocks as mediators of the homeorhetic response to lactation. J. Anim. Sci. 2012, 90, 744–754. [Google Scholar] [CrossRef] [PubMed]
- Porter, W. Circadian clocks in mammary gland development and differentiation. J. Anim. Sci. 2011, 89, 185. [Google Scholar]
- Giannetto, C.; Piccione, G. Daily rhythms of 25 physiological variables in bos taurus maintained under natural conditions. J. Appl. Biomed. 2009, 7, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Lefcourt, A.M.; Bitman, J.; Wood, D.L.; Akers, R.M. Circadian and ultradian rhythms of peripheral growth hormone concentrations in lactating dairy cows. Domest. Anim. Endocrinol. 1995, 12, 247–256. [Google Scholar] [CrossRef]
- Lefcourt, A.M.; Akers, R.M.; Wood, D.L.; Bitman, J. Circadian and ultradian rhythms of peripheral prolactin concentrations in lactating dairy cows. Am. Physiol.-Reg. I 1994, 267, R1461–R1466. [Google Scholar] [CrossRef]
- Lefcourt, A.M.; Bitman, J.; Kahl, S.; Wood, D.L. Circadian and ultradian rhythms of peripheral cortisol concentrations in lactating dairy cows. J. Dairy Sci. 1993, 76, 2607–2612. [Google Scholar] [CrossRef]
- Harvatine, K.; Allen, M. Effects of fatty acid supplements on feed intake, and feeding and chewing behavior of lactating dairy cows. J. Dairy. Sci. 2006, 89, 1104–1112. [Google Scholar] [CrossRef]
- Quist, M.A.; LeBlanc, S.J.; Hand, K.J.; Lazenby, D.; Miglior, F.; Kelton, D.F. Milking-to-milking variability for milk yield, fat and protein percentage, and somatic cell count. J. Dairy Sci. 2008, 91, 3412–3423. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, G.R.; Hargrove, G.L.; Kroger, M. Diurnal variation in milk yield, fat yield, milk fat percentage, and milk protein percentage of Holstein-Friesian cows. J. Dairy Sci. 1972, 56, 409–410. [Google Scholar] [CrossRef]
- Adamovich, Y.; Rousso-Noori, L.; Zwighaft, Z.; Neufeld-Cohen, A.; Golik, M.; Kraut-Cohen, J.; Wang, M.; Han, X.L.; Asher, G. Circadian clocks and feeding time regulate the oscillations and levels of hepatic triglycerides. Cell Metab. 2014, 19, 319–330. [Google Scholar] [CrossRef] [Green Version]
- Grimaldi, B.; Bellet, M.M.; Katada, S.; Astarita, G.; Hirayama, J.; Amin, R.H.; Granneman, J.G.; Piomelli, D.; Leff, T.; Sassone-Corsi, P. PER2 controls lipid metabolism by direct regulation of PPARg. Cell Metab. 2010, 12, 509–520. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.M.; Huang, S.F.; Zeng, J.M.; Liu, D.D.; Xiao, Q.; Tian, W.J.; Zhu, X.D.; Huang, Z.G.; Feng, W.L. Per2 inhibits k562 leukemia cell growth in vitro and in vivo through cell cycle arrest and apoptosis induction. Pathol. Oncol. Res. 2010, 16, 403–411. [Google Scholar] [CrossRef]
- Cheng, A.Y.; Zhang, Y.; Mei, H.J.; Fang, S.; Ji, P.; Yang, J.; Yu, L.; Guo, W.C. Construction of a plasmid for overexpression of human circadian gene period2 and its biological activity in osteosarcoma cells. Tumor Biol. 2015, 36, 3735–3743. [Google Scholar] [CrossRef]
- Gaucher, J.; Montellier, E.; Sassone-Cors, P. Molecular cogs: Interplay between circadian clock and cell cycle. Trends Cell Biol. 2018, 28, 368–379. [Google Scholar] [CrossRef]
- Bionaz, M.; Thering, B.J.; Loor, J.J. Fine metabolic regulation in ruminants via nutrient–gene interactions: Saturated long-chain fatty acids increase expression of genes involved in lipid metabolism and immune response partly through PPAR-α activation. Brit. J. Nutr. 2012, 107, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.Y.; Wang, M.Z.; Ouyang, J.L.; Li, P.F.; Loor, J.J. Period2 gene silencing increases the synthesis of αs-casein protein in bovine mammary epithelial cells. J. Anim. Sci. 2017, 95, 4510–4513. [Google Scholar] [CrossRef] [PubMed]
- Chappuis, S.; Ripperger, J.A.; Schnell, A.; Rando, G.; Jud, C.; Wahli, W.; Albrecht, U. Role of the circadian clock gene Per2 in adaptation to cold temperature. Mol. Metab. 2013, 2, 184–193. [Google Scholar] [CrossRef]
- Sun, L.; Wang, Y.; Song, Y.; Cheng, X.R.; Xia, S.F.; Rahman, M.R.T.; Shi, Y.H.; Le, G.W. Resveratrol restores the circadian rhythmic disorder of lipid metabolism induced by high-fat diet in mice. Biochem. Bioph. Res. Commun. 2015, 458, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Gooley, J.J. Circadian regulation of lipid metabolism. Proc. Nutr. Soc. 2016, 75, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Bionaz, M.; Chen, S.; Khan, M.J.; Loor, J.J. Functional role of PPARs in ruminants: Potential targets for fine-tuning metabolism during growth and lactation. PPAR Res. 2013, 84159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porstmann, T.; Santos, C.R.; Griffiths, B.; Cully, M.; Wu, M.; Leevers, S.; Griffiths, J.R.; Chung, Y.L.; Schulze, A. SREBP activity is regulated by mTORC1 and contributes to Akt-dependent cell growth. Cell Metab. 2008, 8, 224–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osorio, J.S.; Lohakare, J.; Bionaz, M. Biosynthesis of milk fat, protein, and lactose: Roles of transcriptional and posttranscriptional regulation. Physiol. Genom. 2016, 48, 231–256. [Google Scholar] [CrossRef] [Green Version]
- Lewis, C.A.; Griffiths, B.; Santos, C.R.; Pende, M.; Schulze, A. Regulation of SREBP transcription factors by mTORC1. Biochem. Soc. Trans. 2011, 39, 495–499. [Google Scholar] [CrossRef] [Green Version]
- Bakan, I.; Laplante, M. Connecting mTORC1 signaling to SREBP-1 activation. Curr. Opin. Lipidol. 2012, 23, 226–234. [Google Scholar] [CrossRef]
- Laplante, M.; Sabatini, D.M. An emerging role of mTOR in lipid biosynthesis. Curr. Biol. 2009, 19, R1046–R1052. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhao, F.; Si, Y.; Huang, Y.L.; Yu, C.P.; Luo, C.C.; Zhang, N.; Li, Q.Z.; Gao, X.J. GSK3β regulates milk synthesis in and proliferation of dairy cow mammary epithelial cells via the mTOR/S6K1 signaling pathway. Molecules 2014, 19, 9435–9452. [Google Scholar] [CrossRef] [Green Version]
- Düvel, K.; Yecies, J.L.; Menon, S.; Raman, P.; Lipovsky, A.I.; Souza, A.L.; Triantafellow, E.; Ma, Q.; Gorski, R.; Cleaver, S.; et al. Activation of a metabolic gene regulatory network downstream of mTOR complex 1. Mol. Cell. 2010, 39, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Xu, G.; Qin, Y.; Zhang, C.; Tang, H.; Yin, Y.; Xiang, X.; Zhao, J.; Mulholland, M.; Zhang, W. Ghrelin promotes hepatic lipogenesis by activation of mTOR-PPARγ signaling pathway. Proc. Natl. Acad. Sci. USA. 2014, 111, 13163–13168. [Google Scholar] [CrossRef] [Green Version]
- Peterson, T.R.; Sengupta, S.S.; Harris, T.E.; Carmack, A.E.; Kang, S.A.; Balderas, E.; Guertin, D.A.; Madden, K.L.; Carpenter, A.E.; Finck, B.N.; et al. mTOR complex 1 regulates lipin 1 localization to control the SREBP pathway. Cell 2011, 146, 408–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.X.; Zhao, K.Y.; Feng, X.; Yang, D.D.; Yao, R.Y.; Chen, Y.H.; Bao, L.L.; Wang, Z.G. mTORC2 regulates lipogenic gene expression through PPARγ to control lipid synthesis in bovine mammary epithelial cells. Biomed. Res. Int. 2019, 5196028. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Zhao, F.; Wei, C.J.; Liang, M.Y.; Zhang, N.; Wang, C.M.; Li, Q.Z.; Gao, X.J. Function of SREBP1 in the milk fat synthesis of dairy cow mammary epithelial cells. Int. J. Mol. Sci. 2014, 15, 16998–17013. [Google Scholar] [CrossRef] [Green Version]
- Kadegowda, A.K.G.; Bionaz, M.; Piperova, L.S.; Erdman, R.A. Lipogenic gene expression in MAC-T cells is affected differently by fatty acids and enhanced by PPAR-gamma activation. J. Dairy Sci. 2008, 91, 678. [Google Scholar]
- AlSaleh, A.; Sanders, T.A.B.; O’Dell, S.D. Effect of interaction between PPARG, PPARA and ADIPOQ gene variants and dietary fatty acids on plasma lipid profile and adiponectin concentration in a large intervention study. Proc. Nutr. Soc. 2012, 71, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Ji, P.; Drackley, J.K.; Khan, M.J.; Loor, J.J. Overfeeding energy upregulates peroxisome proliferator-activated receptor (PPAR) γ-controlled adipogenic and lipolytic gene networks but does not affect proinflammatory markers in visceral and subcutaneous adipose depots of Holstein cows. J. Dairy Sci. 2014, 97, 3431–3440. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.S.; Goldstein, J.L. The SREBP pathway: Regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell 1997, 89, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Bionaz, M.; Loor, J.J. Gene networks driving bovine milk fat synthesis during the lactation cycle. BMC Genom. 2008, 9, 366. [Google Scholar] [CrossRef] [Green Version]
- Bionaz, M.; Loor, J.J. ACSL1, AGPAT6, FABP3, LPIN1, and SLC27A6 are the most abundant isoforms in bovine mammary tissue and their expression is affected by stage of lactation. J. Nutr. 2008, 138, 1019–1024. [Google Scholar] [CrossRef]
- Ma, L.; Corl, B.A. Transcriptional regulation of lipid synthesis in bovine mammary epithelial cells by sterol regulatory element binding protein-1. J. Dairy Sci. 2012, 95, 3743–3755. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.C. Tumor suppression by the mammalian period genes. Cancer Causes Control 2006, 17, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Rosbash, M.; Takahashi, J.S. Circadian rhythms: The cancer connection. Nature 2002, 420, 373–374. [Google Scholar] [CrossRef]
- Yang, X.; Wood, P.A.; Oh, E.Y.; Quiton, J.D.; Ansell, C.M.; Hrushesky, W.J.M. Down regulation of circadian clock gene Period 2 accelerates breast cancer growth by altering its daily growth rhythm. Breast Cancer Res. Treat. 2009, 117, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Wang, J. Initiator caspases in apoptosis signaling pathways. Apoptosis 2002, 7, 313–319. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, Y.; Chen, Y.; Wang, S.; Ouyang, J.; Hu, L.; Yang, Q.; Wang, M.; Zhang, B.; Loor, J.J. Circadian Gene PER2 Silencing Downregulates PPARG and SREBF1 and Suppresses Lipid Synthesis in Bovine Mammary Epithelial Cells. Biology 2021, 10, 1226. https://doi.org/10.3390/biology10121226
Jing Y, Chen Y, Wang S, Ouyang J, Hu L, Yang Q, Wang M, Zhang B, Loor JJ. Circadian Gene PER2 Silencing Downregulates PPARG and SREBF1 and Suppresses Lipid Synthesis in Bovine Mammary Epithelial Cells. Biology. 2021; 10(12):1226. https://doi.org/10.3390/biology10121226
Chicago/Turabian StyleJing, Yujia, Yifei Chen, Shan Wang, Jialiang Ouyang, Liangyu Hu, Qingyong Yang, Mengzhi Wang, Bin Zhang, and Juan J. Loor. 2021. "Circadian Gene PER2 Silencing Downregulates PPARG and SREBF1 and Suppresses Lipid Synthesis in Bovine Mammary Epithelial Cells" Biology 10, no. 12: 1226. https://doi.org/10.3390/biology10121226
APA StyleJing, Y., Chen, Y., Wang, S., Ouyang, J., Hu, L., Yang, Q., Wang, M., Zhang, B., & Loor, J. J. (2021). Circadian Gene PER2 Silencing Downregulates PPARG and SREBF1 and Suppresses Lipid Synthesis in Bovine Mammary Epithelial Cells. Biology, 10(12), 1226. https://doi.org/10.3390/biology10121226