Simple Summary
Zinc ions are involved in the biology of cell growth, proliferation, differentiation or apoptosis by regulating many biological molecules, such as transcription factors, enzymes and growth factors. In this study, the time-dependent cytotoxicity, cell proliferation and gene expression in human keratinocytes HaCaT cells were evaluated when exposed to ZnCl2. The results of this study showed non-cytotoxic effects up to 10 µg/mL after 24 h, no significant effect on cell proliferation when exposed to 5 or 1 µg/mL ZnCl2 at 72 h and upregulation of eight genes, with great potential in the biomedical field, particularly for regenerative-medicine applications and wound healing.
Abstract
The use of ionic metals such as zinc (Zn2+) is providing promising results in regenerative medicine. In this study, human keratinocytes (HaCaT cells) were treated with different concentrations of zinc chloride (ZnCl2), ranging from 1 to 800 µg/mL, for 3, 12 and 24 h. The results showed a time–concentration dependence with three non-cytotoxic concentrations (10, 5 and 1 µg/mL) and a median effective concentration value of 13.5 µg/mL at a cell exposure to ZnCl2 of 24 h. However, the zinc treatment with 5 or 1 µg/mL had no effect on cell proliferation in HaCaT cells in relation to the control sample at 72 h. The effects of the Zn2+ treatment on the expression of several genes related to glycoprotein synthesis, oxidative stress, proliferation and differentiation were assessed at the two lowest non-cytotoxic concentrations after 24 h of treatment. Out of 13 analyzed genes (superoxide dismutase 1 (SOD1), catalase (CAT), matrix metallopeptidase 1 (MMP1), transforming growth factor beta 1 (TGFB1), glutathione peroxidase 1 (GPX1), fibronectin 1 (FN1), hyaluronan synthase 2 (HAS2), laminin subunit beta 1 (LAMB1), lumican (LUM), cadherin 1 (CDH1), collagen type IV alpha (COL4A1), fibrillin (FBN) and versican (VCAN)), Zn2+ was able to upregulate SOD1, CAT, TGFB1, GPX1, LUM, CDH1, FBN and VCAN, with relative expression levels of at least 1.9-fold with respect to controls. We found that ZnCl2 promoted glycoprotein synthesis and antioxidant gene expression, thus confirming its great potential in biomedicine.
1. Introduction
Zinc ions (Zn2+) are involved in all the crucial decisions in the life of mammalian cells related to growth, proliferation, differentiation or apoptosis, both in its ionic or protein-bound form [1]. They are implicated in regulating many biological molecules, such as transcription factors, enzymes and growth factors [2,3], and have been shown to be crucial in hundreds of enzymatic reactions and required for thousands of transcription factors which regulate gene expression [4]. Zn2+ has also been recently recognized as an intra- and intercellular signaling mediator, acting similar to calcium as a second messenger to transduce extracellular stimuli into intracellular signaling events [3,4].
Zn2+, with a mass of 0.8–3 g in the human body, can be found in tissues such as muscle and bone, which act as a major tissue reservoir (85% of the whole body); skin (5%); liver (5%); and the remaining percentage is distributed around other organs, such as the brain, pancreas and kidneys. Zn2+ deficiency is related to delayed bone development and dwarfism; in addition, several skin disorders are associated with Zn2+ deficiency [4,5,6,7].
The essential role played by Zn2+ ions in cell behavior, modulating cell signaling, has attracted increasing attention in the biomedical field, particularly for regenerative-medicine applications and wound healing [8,9,10,11,12,13,14,15]. In addition, Zn+2 presents antimicrobial activity, both as an antibacterial and antifungal agent [16,17]. However, it has been reported that, to achieve effective antimicrobial activity on common wound pathogens, such as Staphylococcus aureus, Escherichia coli, Pseudomonas aeruginosa and Candida albicans, higher concentrations of Zn compounds are required than those used to promote cell response [18]. In that study, doses of zinc sulfate and zinc gluconate lower than 10 µg/mL showed no antimicrobial properties, but they exerted bioactivity in nutrient-deficient environments.
Regenerative medicine involves the regeneration of human tissues, with the aim of returning the patient to full health, using bioactive factors to act in conjunction with the natural healing potential [19]. Among the bioactive factors that have shown a great potential, metallic ions (including Zn2+) have emerged, based on their unique properties to modulate cell response [12]. Zn2+-based biomaterials have recently been developed, including bioactive glasses [8,9], nanocomposites with ZnO nanoparticles [20] and hydrogels [21,22,23,24]. Mesoporous bioactive glasses containing Zn2+ have been reported as potential biomaterials for soft-tissue repair and wound healing [9], whilst ZnO nanoparticles with different nanostructures have been shown to promote adhesion, growth and differentiation in several cell lines [25,26]. Nanocomposite hydrogels based on alginate/graphene oxide [21] or carboxymethylcellulose/ZnO [24] crosslinked with Zn2+ ions have been prepared to provide bioactivity and antimicrobial properties. Zn2+ ions have been shown to promote tissue formation and inhibit resorption in musculoskeletal disorders [27].
Zn2+-based compounds such as calamine or zinc oxide have been applied topically for centuries to sooth and calm skin irritations and enhance wound healing [28]. Its use has also expanded over the years for several dermatological conditions, including infections inflammatory dermatoses, pigmentary disorders and neoplasia. Therapeutically, Zn2+ can be used both topically and in systemic form; however, systemic Zn2+ therapy needs further experimental and clinical evidences [28,29]. Zn2+ plays an important physiological function throughout the stages of wound healing, associated with inflammation and immune response [4,30,31]. The importance of Zn2+ concentration for wound healing has been reported in patients with thermal injuries or exposure to surgical stress [15]. In human skin, the epidermis contains higher concentrations of Zn2+ than the dermis (60 µg/g vs. 40 µg/g), due to a Zn2+ requirement for epidermal keratinocytes proliferation and differentiation [32]. Keratinocytes are the main cell components of the epidermis (95%), with several key function in the wound-healing process. The signaling interaction between keratinocytes and other cells that participate in wound healing is crucial to wound closure [33]. Zn2+ therapies have been used in wound care to promote healing in patients with Zn2+ deficiency, as has topical zinc sulfate or zinc chloride (ZnCl2), due to its antioxidant effect [15]. Zn2+ is crucial for normal skin function and wound healing, acting as a bioactive factor mimicking the action of growth factors by promoting intracellular mitogenic signaling pathways [34]. It also acts as a stabilizer of the cell membrane of keratinocytes, being found both intracellularly and within the skin extracellular matrix [35]. However, high concentrations of extracellular Zn2+ ions have cytotoxic effects, due to the balance between extracellular–intracellular Zn2+ concentration and cell survival [35]. Thus, whilst Zn2+-based therapy is a promising tool in wound healing and dermatological diseases, a deeper understanding of the cellular behavior induced by Zn2+ is still needed in order to identify the boundaries that limit its safe therapeutic application. Therefore, the aim of this study was to analyze the biological response in terms of time-dependent cytotoxicity, cell proliferation and gene expression in human keratinocytes HaCaT cells when exposed to extracellular Zn2+(from ZnCl2). Gene expression associated with glycoprotein synthesis, oxidative stress, proliferation and differentiation was investigated for the first time to obtain further insight into the physiological roles and mechanisms of Zn2+ action.
2. Materials and Methods
2.1. Materials
ZnCl2 (≥97.0%, bioreagent for molecular biology suitable for cell culture) and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) were acquired from Sigma-Aldrich (St. Louis, MO, USA). Dulbecco’s modified Eagle’s medium (DMEM), fetal bovine serum (FBS), L-Glutamine, penicillin–streptomycin (P/S), phosphate-buffered saline (PBS), trypsin–EDTA, dimethyl sulfoxide (DMSO) and epidermal growth factor were purchased from Life Technologies (Gibco, Karlsruhe, Germany).
2.2. Preparation of ZnCl2 Stock Solution
A stock solution of ZnCl2 was prepared in sterile DMEM low glucose supplemented with P/S and L-Glutamine by sonication for 2 h to dissolve the salt completely and obtain a homogeneous solution. The solution was used immediately after the sonication process as source of Zn2+ for in vitro cytotoxicity experiments.
2.3. Cell Culture
Immortalized human keratinocytes (cell line HaCaT) were supplied by the Medical Research Institute Hospital La Fe (Valencia, Spain). They were cultured in DMEM low glucose, containing l-glutamine, supplemented with 10% FBS and 1% P/S (complete medium) in a humidified atmosphere, at 5% CO2 and 37 °C. Culture medium was changed every 2 days, and cells were trypsinized for 3 min and resuspended in the same medium when culture achieved 80% confluence.
2.4. Cell Viability Assay
Cytotoxicity was assessed for keratinocytes exposed to different Zn2+ concentrations by performing an MTT assay [36], based on the metabolic reduction of the tetrazolium dye to a colored compound (formazan) through the mitochondrial succinate dehydrogenase. Cells were trypsinized and resuspended in complete medium, centrifuged and resuspended again in medium with 1% P/S and L-Glutamine, but without FBS, in order to avoid any chemical interaction with the zinc divalent cations. Then 96-micro-well plates were seeded at a low density (104 cells/well) and placed in the incubator. After 24 h to allow cell adhesion, the culture medium was substituted by growth medium supplemented with 100 µL of ZnCl2 solution at concentrations between 1 and 800 µg/mL of Zn2+. Cells with medium without Zn2+ were used as control, and culture medium was used as reagent blank. Cytotoxicity was measured at different end points: 3, 12 and 24 h. After the exposure time, the culture medium was removed, and 100 µL of MTT working solution was added to each well. Plates were incubated for 3 h, at 37 °C. After removing the reagent solution and rinsing with PBS, we added 100 µL of DMSO to dissolve the formazan crystals and incubated for 1 h, at 37 °C. The absorbance of the resulting solution was measured at 550 nm to determine cell viability, using the Varioskan Lux microplate reader (ThermoScientificTM, Dreieich, Germany). Median effective concentration (EC50 assay) was obtained for ZnCl2 concentrations 1, 14, 16 and 18 µg/mL (see Results). EC50 values were calculated as the concentration where the sigmoidal curve attains values of 50%. Experiments were performed in sextuplicate on the same plate.
2.5. Proliferation Assay
Cell proliferation was determined by using the MTT assay [36] 72 h after ZnCl2 treatment. Following the same protocol described to assess cell viability, 5·103 cells/well were seeded in a 96-multi-well culture plate. After 24 h, the growth medium was replaced by DMEM+1%P/S medium supplemented with 0.5%FBS to avoid complete starvation [37], together with ZnCl2 at specific concentrations, and cultured in a humidified atmosphere of 5% CO2 and 37 °C. As positive control, cells treated with growth medium (DMEM+1%P/S+0.5%FBS) and were supplemented with Epidermal Growth Factor (EGF) at a concentration of 15 ng/mL. Moreover, a control group with only growth medium was included. Based on the cell viability assay results, two non-cytotoxic concentrations of ZnCl2 at 24 h were chosen (1 and 5 µg/mL) to perform this assay. Experiments were performed in sextuplicate on the same plate.
2.6. Gene Expression
Real-time reverse-transcription polymerase chain reaction (RT-qPCR) was carried out to analyze the gene expression in HaCaT cells upon treatment with extracellular Zn2+. Cells were seeded at a density of 1.5·105 cells/well in a 6-well plate and cultured in a CO2 incubator, following the same protocol used in the cytotoxicity assays. After 24 h, cells were treated with two non-cytotoxic concentrations of ZnCl2 (1 and 5 µg/mL), i.e., the same used in the proliferation assay, and cultured for 24 h. After removing culture medium and rinsing with PBS, we added the extraction solution, and the plates were frozen in liquid nitrogen. RNA purification kit (Norgen, Thorold, ON, Canada) was used for RNA isolation, following the protocol provided by the manufacturer. The quality and the concentrations of the different samples were determined by using a Nanodrop™ One (ThermoScientific™, Dreieich, Germany) and a PrimeScript™ RT Reagent Kit (Perfect Real Time) (Takara Bio, Inc., Kusatsu, Japan) to synthetized cDNA. Quantitative PCR, TB Green Premix Ex Taq (Takara Bio, Inc.), was performed by following the protocol provided by the manufacturer in a 384 QuantStudio 5 (ThermoScientific™, Dreieich, Germany). Data analysis was carried out by using QuantStudioTM software. Primers of target genes and the reference gene (β-actin/ACTB), which are reported in Table 1, were measured by using Primer-Blast software (available on: http://www.ncbi.nlm.nih.gov/tools/primer-blast, accessed on 18 October 2021). Data normalization was performed based on the reference gene expression. Experiments were performed in triplicate on the same plate.

Table 1.
Details of specific genes used in the RT-qPCR assay.
2.7. Statistical Analysis
Data were reported as mean ± standard deviation (SD). Statistical analysis was conducted through GraphPad Prism 6 software (GraphPad Inc., San Diego, CA, USA). Statistical differences were tested by one-way ANOVA with Tukey’s correction for multiple comparisons. The EC50 values were determined by using Probit analysis. Significance was assumed at p-values < 0.05 (95% confidence).
3. Results and Discussion
3.1. Cytotoxicity Assay
To evaluate the cytotoxicity of ZnCl2, HaCaT cells were cultured in culture medium supplemented with increasing concentrations of ZnCl2, ranging from 1 to 800 µg/mL. The results were assessed at different exposure times (3, 12 and 24 h), and cell viability was determined by using the MTT-based colorimetric assay. Figure 1 shows the cytotoxicity results at 3, 12 and 24 h after exposure.
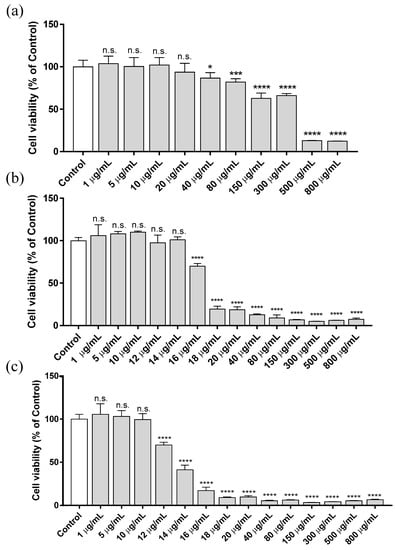
Figure 1.
Cytotoxicity assay (MTT method) in human keratinocytes HaCaT cell line after exposing the cells for 3 h (a), 12 h (b) and 24 h (c) to several concentrations of ZnCl2, ranging from 1 to 800 µg/mL. Results represented are normalized to the control group (culture media without ZnCl2). Data depicted as mean ± SD of six replicates. Significant differences with respect to control were determined by one-way ANOVA with Tukey’s correction for multiple comparisons: * p > 0.05; *** p > 0.001; **** p > 0.0001; n.s. = not significant.
The EC50 calculated for each exposure time is reported in Table 2. The highest cytotoxic effect due to ZnCl2 occurs within the first 12 h of exposure. The EC50 at 12 h was more than 90% lower than that of the concentration obtained after 3 h of treatment. However, between 12 and 24 h of treatment, there was only a 20% reduction in EC50. In fact, it was only possible to detect this difference in EC50 between 12 and 24 h with the addition of several concentrations between 10 and 20 µg/mL. These results indicate that the main cytotoxic effect of Zn2+ added to the culture medium takes place in the first hours of exposure.
Table 2.
Exposure of human keratinocyte HaCaT cells to ZnCl2 for 3, 12 and 24 h. Median effective concentration (EC50), confidence limits 95% (CI) as mass/volume in µg/mL and R square.
Zn2+-induced cytotoxicity depends on the cell line [38]. Studies in human lens epithelial cell line (HLE B-3) have shown a high ZnCl2 sensitive dose-dependent response, where the cytotoxic effect was obtained with 5 µg/mL of ZnCl2 after 72 h of treatment [39]. Evident cytotoxicity produced by ZnCl2 exposure was observed in C6 rat glioma at a concentration of 25 µg/mL (24 h after the treatment) [40]. In vivo assays performed in Wistar rats showed that concentrations below 10 mg/kg did not induce mortality in the exposed animals. An increase in lethality was observed when the dose concentration was increased, inducing a 10% mortality at 20 mg/kg body weight, with 60 mg/kg of LC50 (lethal concentration 50) [41]. In that study, a drastic reduction in cell viability was observed with an in vitro concentration of 20 µg/mL at 12 and 24 h of exposure, which is in the same order of magnitude as our results. Other Zn2+-based compounds, such as ZnO, induced higher toxicity in laryngeal cells (viability lower than 40%) at a concentration of 10 µg/mL after 24 h of exposure [42].
3.2. Proliferation Assay
In the cytotoxicity assay, it was found that Zn2+ concentration became cytotoxic at concentrations from 12 µg/mL onwards after 24 h of Zn2+ exposure. Thus, two non-cytotoxic ZnCl2 concentrations were chosen for the proliferation assays, namely 1 and 5 µg/mL, to minimize the cytotoxic effect at a longer time (72 h). Thus, proliferation was assessed after 72 h of ZnCl2 exposure in human keratinocytes (Figure 2).
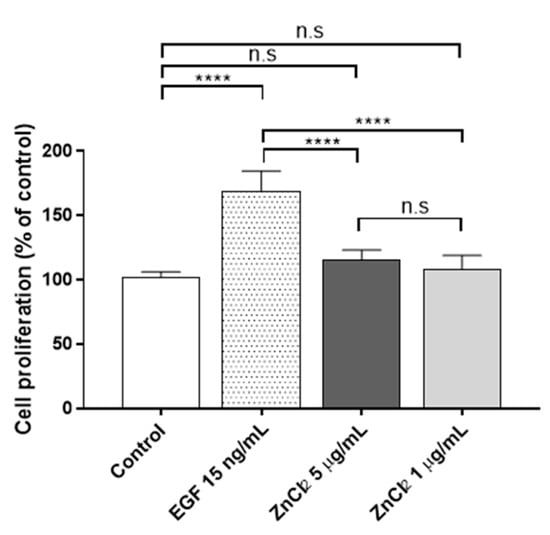
Figure 2.
Proliferative activity of ZnCl2 after 72 h of exposure in human keratinocytes. Results represented are normalized to the control group (culture media without ZnCl2). Data depicted as mean ± SD of six replicates. Significant differences were determined by one-way ANOVA with Tukey’s correction for multiple comparisons: **** p > 0.0001; n.s. = not significant.
In both concentrations, the zinc treatment had no significant effect on cell proliferation after 72 h of culture. In a previous study [43], it was found that a higher concentration of extracellular ZnCl2 (100 µM (~13 µg/mL)) induced slight but significant cell proliferation after 72 h of exposure (1.3-fold; p = 0.045) in HaCaT cells. In the present study, concentrations of extracellular Zn2+ higher than 12 µg/mL were found to show a slight cytotoxicity after 24 h of treatment and therefore were discarded for the proliferation assays. In good agreement with our results, Emri et al. [43] showed that a close extracellular concentration of Zn2+ 50 µM (~6.5 µg/mL) had no effect on cell proliferation in HaCaT cells. However, many controversial results have been published in the literature with regards to the toxicity of Zn2+ ions [35], and further investigation is therefore required. It should be noted that the culture media, as well as the different techniques used in the experiments, can give rise to differences in the outcomes. In Holmes’s study [35], the researchers found that the results from the cytotoxicity assay, performed after 24 h of the Zn2+ treatment in HaCaT cells with extracellular ZnSO2, depended on the medium used in the cell culture: complete DMEM, complete DMEM with 1.75% bovine serum albumin (BSA) or complete DMEM with 1 mM Ethylenediaminetetraacetic acid solution (EDTA). Both BSA and EDTA can act as Zn2+ chelators, reducing the Zn2+ toxicity. The techniques used in the cytotoxic studies (EZ4U or MTS assays) also affect the results [35,43].
3.3. Gene Expression
The protective effect of non-cytotoxic ZnCl2 concentrations against UVB radiation was studied in human epidermal keratinocytes [43]. Although the data showed a reduction on pyrimidine dimers in cells pretreated with ZnCl2, significantly enhanced superoxide generation was found 10 h after UVB treatment. The results of that study therefore suggested that the exposure of human keratinocytes to non-toxic concentrations of zinc chloride impacts gene expression, cell proliferation and the responses to external stress (such as UVB radiation) in the skin. It was also demonstrated that primary keratinocytes exposed to nM concentration of zinc pyrithione (ZnPT) loosed genomic integrity [44]. Exposure of HaCaT cells to ZnO nanoparticles induces the expression of genes involved in oxidative stress [45]. SOD genes were significantly higher in cells exposed to Zn nanoparticles, suggesting its potential to induce intracellular reactive oxygen species (ROS) and oxidative stress.
To determine the effects of exposure to extracellular Zn2+ ions on the expression of genes involved in different metabolic routes, thirteen genes, which are related to the glycoprotein synthesis associated with repair and maintenance of different tissues, oxidative stress and damage, proliferation, differentiation and cell growth, were analyzed (Table 1). Figure 3 shows the results obtained after analyzing gene expression in HaCaT cells treated for 24 h with 1 or 5 µg/mL of extracellular Zn2+, referred to the housekeeping gene (β-actin/ACTB).
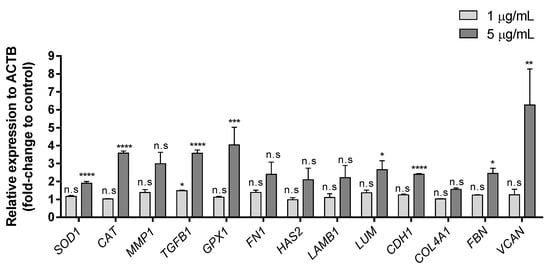
Figure 3.
Effects of Zn2+ on HaCaT cell line after 24-h treatment. Expression of different genes at 1 and 5 µg/mL ZnCl2 concentration. Data depicted as mean ± SD of three replicates. Results represented as fold-change to control and relative expression to ACTB. Significant differences were determined by one-way ANOVA with Tukey’s correction for multiple comparisons: * p < 0.05; ** p < 0.01; *** p < 0.001; **** p > 0.0001; n.s. = not significant.
The lowest concentration (1 µg/mL) did not show significant differences in any of the genes analyzed (only TGFB1 showed a slight upregulation). Nevertheless, exposure to 5 µg/mL induced significant upregulation in eight genes (SOD1, CAT, TGFB1, GPX1, LUM, CDH1, FBN and VCAN) with relative expression levels of at least 1.9-fold with respect to untreated controls. VCAN gene showed the highest overexpression after the extracellular Zn2+ treatment (6.265-fold). This gene is involved in cell adhesion, proliferation, migration and angiogenesis and plays an important role in tissue morphogenesis and maintenance [46]. It has also been reported that keratinocytes express versican during active cell proliferation [47]. The expression on this gene was upregulated in association with healing/restructuring processes following non-surgical periodontal treatments [48]. In addition, cell exposure to ZnCl2 at a concentration of 5 µg/mL upregulated the genes involved in glycoprotein synthesis. Gene expression of FBN, LUM and CDH1 was upregulated (2-fold). Type I cadherins includes vital transmembrane glycoproteins for the morphogenesis and development of normal animal tissue [49]. Studies carried out on UV-irradiated HaCaT cells showed how the decrease in the synthesis of these glycoproteins compromises cell–cell adhesion in epithelial tissues, thus compromising the integrity of the skin after radiation [50]. Moreover, LUM is the major keratan sulfate proteoglycan of the cornea, and it is distributed throughout the body in interstitial collagenous matrices. It has been reported in this cell line that lumican is involved in inflammatory cells infiltration [51] and angiogenesis [52], which are essential in wound healing. Studies performed on fibroblasts and keratinocytes demonstrated that lumican can modulate fibroblast activation via integrin a2, whereas keratinocytes were not affected [53]. In our study, LUM gene expression was activated by the exposure to zinc chloride, which could lead to triggering the signaling cascade and subsequent activation of cells in the lower layers of the skin. FBN is an extracellular matrix glycoprotein that provides force-bearing structural support in elastic and non-elastic connective tissue throughout the body and serves as a structural component of calcium-binding microfibrils [54]. It has also been related to growth-factor signaling, being crucial for correct growth and balance of skin homeostasis [55]. During metabolic processes, ROS are present due to the requirements for normal cell signaling. In the same way as with lumican, the expression of fibrillin increased in fibroblasts after treatment with xanthohumol, which improves the structure and firmness of the skin [56]; moreover, it has also been demonstrated that these glycoproteins can be synthetized by HaCaT cells [57]. However, high ROS levels cause severe cellular damage and may lead to the development of skin diseases, including cancer [58,59]. Induction of gene expression involved in oxidative stress could protect cells against oxidative cellular damage in the presence of different external stressors. Vitamin E has been used as antioxidant to pre-protect human epidermal keratinocytes and enhance their therapeutic ability under induced oxidative microenvironments [60]. Other exogenous antioxidants have also been reported recently, such as extract of Rhodiola rosea L. roots [61], cannabidiol [62] and other different antioxidant agents [63]. Recently, a compound of nicotinamide, a soluble vitamin B3 well-known as an antioxidant, and a jellyfish peptide (nicotinyl–isoleucine–valine–histidine peptide) have also been reported to promote antioxidant gene expression in HaCaT Cells [64].
CAT, GPX1 and TGFB1 were upregulated with fold-changes between 3 and 4. CAT and GPX1 genes act as relevant antioxidants, encoding the synthesis of enzymes related in the neutralization of H2O2. UVA and UVB irradiation causes biological damage by inducing an increase in ROS levels in human keratinocytes [65,66], causing cellular oxidative stress, damage to cell components and genomic instability [67,68]. UVB induces an increase in ROS levels in two stages, immediately following UVB irradiation and several hours after irradiation [69]. It has been shown that CAT overexpression has a protective role against UVB irradiation by preventing DNA damage mediated by the late ROS increase [66]. In addition, UVA induces upregulation of the transcription factor AP-2α, involved in epidermal differentiation, which can be attenuated by the overexpression of GPX1 [65]. Both CAT and GPX1 were upregulated after extracellular Zn2+ treatment for 24 h at a concentration of 5 µg/mL. Exposure to 1 µg/mL did not show any change in gene regulation; thus, a higher concentration exposure is required to activate these defense mechanisms in HaCaT cells. In addition, previous studies in the same cell line treated with caffeic acid or ferulic acid [70] concluded that this upregulation could result in a protective effect on cells against oxidative stressors. TGFB 1, a well-established melanoma suppressor gene [71], which is also involved in cell proliferation, differentiation and growth regulation, was also activated when cells were treated with the highest extracellular Zn2+ concentration (5 µg/mL).
The protein encoded by SOD1 gene binds copper and zinc ions and it is one of two isozymes responsible for destroying free superoxide radicals in the body. SOD 1 is the representative antioxidant enzyme among all SOD enzymes. It can be activated by different substances, such as genoposidic acid [72], sinapic acid [73], or nicotinyl–isoleucine–valine–histidine [64]. However, other promising compounds such as graphene oxide, are not able to upregulate the SOD1 gene using non-cytotoxic concentrations [74]. In the present study, SOD1 was upregulated with extracellular Zn2+ at the highest concentration (5 µg/mL), suggesting that non-cytotoxically extracellular Zn2+ concentrations are able to induce an antioxidant effect in HaCaT keratocytes.
Therefore, we have shown that ZnCl2 could be a promising bioactive antioxidant against external factors and promote skin regeneration in wound healing or skin-disease therapeutics, thus confirming its great potential in biomedicine. However, further research is required to fully assess the efficacy and safety before clinical use.
4. Conclusions
The results show a time-dependent cytotoxity of ZnCl2 with non-cytotoxic concentrations up to 10 µg/mL in human keratinocyte HaCaT cells treated for 3, 12 and 24 h. However, 5 µg/mL of ZnCl2 induced significant upregulation in eight genes (SOD1, CAT, TGFB1, GPX1, LUM, CDH1, FBN and VCAN), with relative expression levels of at least 1.9-fold with respect to untreated controls. ZnCl2 promoted glycoprotein synthesis and antioxidant gene expression in this cell line. Therefore, we have shown that ZnCl2 could be a promising antioxidant compound against external factors. It could be used as a bioactive agent and promoter of regeneration in wound healing or skin-disease treatments. This study demonstrates the great potential of Zn2+ ions in the form of ZnCl2 salt in biomedical applications.
Author Contributions
Conceptualization, resources and writing—original draft preparation, B.S., R.S.i.S. and Á.S.-A.; methodology, software, formal analysis, investigation, data curation, validation and visualization, B.S. and Á.S.-A.; funding acquisition and writing—review and editing, R.S.i.S. and Á.S.-A.; supervision and project administration, Á.S.-A. All authors have read and agreed to the published version of the manuscript.
Funding
This study was founded by the Fundación Universidad Católica de Valencia San Vicente Mártir, Grant 2020-231-006UCV, the Spanish Ministry of Science and Innovation (PID2020-119333RB-I00/AEI/10.13039/501100011033) (awarded to Á.S.-A.) and the FEDER/Spanish Ministry of Science and Innovation-Agencia Estatal de Investigación) through the Project RTI2018-097862-B-C21 (awarded to R.S.i.S). CIBER-BBN is an initiative funded by the VI National R&D&I Plan 2008–2011, Iniciativa Ingenio 2010, Consolider Program. CIBER Actions are financed by the Instituto de Salud Carlos III with assistance from the European Regional Development Fund.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors would like to thank the founding institutions of this research study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Beyersmann, D.; Haase, H. Functions of zinc in signaling, proliferation and differentiation of mammalian cells. BioMetals 2001, 14, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Fukada, T.; Kambe, T. (Eds.) Zinc Signaling; Springer Nature Singapore Pte Ltd.: Singapore, 2019; ISBN 9789811505577. [Google Scholar]
- Hara, T.; Takeda, T.A.; Takagishi, T.; Fukue, K.; Kambe, T.; Fukada, T. Physiological roles of zinc transporters: Molecular and genetic importance in zinc homeostasis. J. Physiol. Sci. 2017, 67, 283–301. [Google Scholar] [CrossRef]
- Ogawa, Y.; Kawamura, T.; Shimada, S. Zinc and skin biology. Arch. Biochem. Biophys. 2016, 611, 113–119. [Google Scholar] [CrossRef]
- Jackson, M.J. (Ed.) Physiology of Zinc: General Aspects; Springer: Berlin/Heidelberg, Germany, 1989. [Google Scholar]
- Tapiero, H.; Tew, K.D. Trace elements in human physiology and pathology: Zinc and metallothioneins. Biomed. Pharmacother. 2003, 57, 399–411. [Google Scholar] [CrossRef]
- Ogawa, Y.; Kinoshita, M.; Shimada, S.; Kawamura, T. Zinc and skin disorders. Nutrients 2018, 10, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamani, D.; Moztarzadeh, F.; Bizari, D. Alginate-bioactive glass containing Zn and Mg composite scaffolds for bone tissue engineering. Int. J. Biol. Macromol. 2019, 137, 1256–1267. [Google Scholar] [CrossRef] [PubMed]
- Neščáková, Z.; Zheng, K.; Liverani, L.; Nawaz, Q.; Galusková, D.; Kaňková, H.; Michálek, M.; Galusek, D.; Boccaccini, A.R. Multifunctional zinc ion doped sol–gel derived mesoporous bioactive glass nanoparticles for biomedical applications. Bioact. Mater. 2019, 4, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Salcedo, S.; Shruti, S.; Salinas, A.J.; Malavasi, G.; Menabue, L.; Vallet-Regí, M. In vitro antibacterial capacity and cytocompatibility of SiO 2-CaO-P2O5 meso-macroporous glass scaffolds enriched with ZnO. J. Mater. Chem. B 2014, 2, 4836–4847. [Google Scholar] [CrossRef]
- Paramita, P.; Ramachandran, M.; Narashiman, S.; Nagarajan, S.; Sukumar, D.K.; Chung, T.W.; Ambigapathi, M. Sol–gel based synthesis and biological properties of zinc integrated nano bioglass ceramics for bone tissue regeneration. J. Mater. Sci. Mater. Med. 2021, 32, 5. [Google Scholar] [CrossRef]
- Mouriño, V.; Cattalini, J.P.; Boccaccini, A.R. Metallic ions as therapeutic agents in tissue engineering scaffolds: An overview of their biological applications and strategies for new developments. J. R. Soc. Interface 2012, 9, 401–419. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Jia, B.; Zhang, Z.; Qu, X.; Li, G.; Lin, W.; Zhu, D.; Dai, K.; Zheng, Y. Alloying design of biodegradable zinc as promising bone implants for load-bearing applications. Nat. Commun. 2020, 11, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, G.K.; Goldman, J.; Aghion, E. The prospects of zinc as a structural material for biodegradable implants—A review paper. Metals 2017, 7, 402. [Google Scholar] [CrossRef] [Green Version]
- Lin, P.H.; Sermersheim, M.; Li, H.; Lee, P.H.U.; Steinberg, S.M.; Ma, J. Zinc in wound healing modulation. Nutrients 2018, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Pasquet, J.; Chevalier, Y.; Pelletier, J.; Couval, E.; Bouvier, D.; Bolzinger, M.A. The contribution of zinc ions to the antimicrobial activity of zinc oxide. Colloids Surf. A Physicochem. Eng. Asp. 2014, 457, 263–274. [Google Scholar] [CrossRef]
- Frígols, B.; Martí, M.; Salesa, B.; Hernández-Oliver, C.; Aarstad, O.; Ulset, A.-S.T.; Sætrom, G.I.; Aachmann, F.L.; Serrano-Aroca, Á. Graphene oxide in zinc alginate films: Antibacterial activity, cytotoxicity, zinc release, water sorption/diffusion, wettability and opacity. PLoS ONE 2019, 14, e0212819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rembe, J.D.; Boehm, J.K.; Fromm-Dornieden, C.; Hauer, N.; Stuermer, E.K. Comprehensive analysis of zinc derivatives pro-proliferative, anti-apoptotic and antimicrobial effect on human fibroblasts and keratinocytes in a simulated, nutrient-deficient environment in vitro. Int. J. Mol. Cell. Med. 2020, 9, 165–179. [Google Scholar] [CrossRef] [PubMed]
- Tabata, Y. Biomaterial technology for tissue engineering applications. J. R. Soc. Interface 2009, 6, S311–S324. [Google Scholar] [CrossRef] [Green Version]
- Laurenti, M.; Cauda, V. ZnO nanostructures for tissue engineering applications. Nanomaterials 2017, 7, 374. [Google Scholar] [CrossRef] [Green Version]
- Sabater i Serra, R.; Molina-mateo, J.; Torregrosa-cabanilles, C.; Andrio-Balado, A.; Meseguer Dueñas, J.; Serrano-Aroca, A. Bio-Nanocomposite Hydrogel Based on Zinc Conformation, Thermal Behavior/Degradation, and Dielectric Properties. Polymers 2020, 12, 702. [Google Scholar] [CrossRef] [Green Version]
- Tiffany, A.S.; Gray, D.L.; Woods, T.J.; Subedi, K.; Harley, B.A.C. The inclusion of zinc into mineralized collagen scaffolds for craniofacial bone repair applications. Acta Biomater. 2019, 93, 86–96. [Google Scholar] [CrossRef]
- De Aragão Tavares, E.; De Medeiros, W.M.T.Q.; De Assis Pontes, T.P.; Barbosa, M.M.; De Araújo, A.A.; De Araújo, R.F.; Figueiredo, J.G.; Leitão, R.C.; Da Silva Martins, C.; Da Silva, F.O.N.; et al. Chitosan membrane modified with a new zinc(II)-vanillin complex improves skin wound healing in diabetic rats. Front. Pharmacol. 2019, 9, 1511. [Google Scholar] [CrossRef]
- Priyadarshi, R.; Kumar, B.; Rhim, J.W. Green and facile synthesis of carboxymethylcellulose/ZnO nanocomposite hydrogels crosslinked with Zn2+ ions. Int. J. Biol. Macromol. 2020, 162, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Ciofani, G.; Genchi, G.G.; Mattoli, V. ZnO nanowire arrays as substrates for cell proliferation and differentiation. Mater. Sci. Eng. C 2012, 32, 341–347. [Google Scholar] [CrossRef]
- Lee, J.; Kang, B.S.; Hicks, B.; Chancellor, T.F.; Chu, B.H.; Wang, H.T.; Keselowsky, B.G.; Ren, F.; Lele, T.P. The control of cell adhesion and viability by zinc oxide nanorods. Biomaterials 2008, 29, 3743–3749. [Google Scholar] [CrossRef]
- Jiménez, M.; Abradelo, C.; San Román, J.; Rojo, L. Bibliographic review on the state of the art of strontium and zinc based regenerative therapies. Recent developments and clinical applications. J. Mater. Chem. B 2019, 7, 1974–1985. [Google Scholar] [CrossRef]
- Bae, Y.S.; Hill, N.D.; Bibi, Y.; Dreiher, J.; Cohen, A.D. Innovative uses for zinc in dermatology. Dermatol. Clin. 2010, 28, 587–597. [Google Scholar] [CrossRef]
- Azgın, İ.; Arbağ, H.; Eryılmaz, M.A.; Çelik, Z.E. The effects of local and intraperitoneal zinc treatments on maxillofacial fracture healing in rabbits. J. Cranio-Maxillofac. Surg. 2020, 48, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Sharir, H.; Zinger, A.; Nevo, A.; Sekler, I.; Hershfinkel, M. Zinc released from injured cells is acting via the Zn2+-sensing receptor, ZnR, to trigger signaling leading to epithelial repair. J. Biol. Chem. 2010, 285, 26097–26106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, D.; Varigos, G.; Ackland, M.L. Apoptosis may underlie the pathology of zinc-deficient skin. Immunol. Cell Biol. 2006, 84, 28–37. [Google Scholar] [CrossRef]
- Michaelsson, G.; Ljunghall, K.; Danielson, B.G. Zinc in epidermis and dermis in healthy subjects. Acta Derm. Venereol. 1980, 60, 295–299. [Google Scholar]
- Pastar, I.; Stojadinovic, O.; Tomic-Canic, M. Role of keratinocytes in healing of chronic wounds. Surg. Technol. Int. 2008, 17, 105–112. [Google Scholar] [PubMed]
- Lansdown, A.B.G.; Mirastschijski, U.; Stubbs, N.; Scanlon, E.; Ågren, M.S. Zinc in wound healing: Theoretical, experimental, and clinical aspects. Wound Repair Regen. 2007, 15, 2–16. [Google Scholar] [CrossRef]
- Holmes, A.M.; Mackenzie, L.; Roberts, M.S. Disposition and measured toxicity of zinc oxide nanoparticles and zinc ions against keratinocytes in cell culture and viable human epidermis. Nanotoxicology 2020, 14, 263–274. [Google Scholar] [CrossRef]
- Li, Y.; Maret, W. Transient fluctuations of intracellular zinc ions in cell proliferation. Exp. Cell Res. 2009, 315, 2463–2470. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Tsuruoka, S.; Ioka, T.; Ando, H.; Ito, C.; Akimoto, T.; Fujimura, A.; Asano, Y.; Kusano, E. Indoxyl sulfate stimulates proliferation of rat vascular smooth muscle cells. Kidney Int. 2006, 69, 1780–1785. [Google Scholar] [CrossRef] [Green Version]
- Bozym, R.A.; Chimienti, F.; Giblin, L.J.; Gross, G.W.; Korichneva, I.; Li, Y.; Libert, S.; Maret, W.; Parviz, M.; Frederickson, C.J.; et al. Free zinc ions outside a narrow concentration range are toxic to a variety of cells in vitro. Exp. Biol. Med. Maywood 2010, 235, 741–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.; Guo, D.; Wu, Q.; Liu, D.; Bi, H. Zinc chloride inhibits human lens epithelial cell migration and proliferation involved in TGF-β1 and TNF-α signaling pathways in HLE B-3 cells. Biol. Trace Elem. Res. 2014, 159, 425–433. [Google Scholar] [CrossRef]
- Salazar-García, S.; García-Rodrigo, J.F.; Martínez-Castañón, G.A.; Ruiz-Rodríguez, V.M.; Portales-Pérez, D.P.; Gonzalez, C. Silver nanoparticles (AgNPs) and zinc chloride (ZnCl2) exposure order determines the toxicity in C6 rat glioma cells. J. Nanopart. Res. 2020, 22, 253. [Google Scholar] [CrossRef]
- Tekuri, S.K.; Bassaiahgari, P.; Gali, Y.; Amuru, S.R.; Pabbaraju, N. Determination of Median Lethal Dose of Zinc chloride in Wistar Rat. Adv. Anim. Vet. Sci. 2021, 9, 393–399. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Guo, Y.; Lu, J.; Veeraraghavan, V.P.; Mohan, S.K.; Wang, C.; Yu, X. Synthesis of Zinc oxide nanoparticles from Marsdenia tenacissima inhibits the cell proliferation and induces apoptosis in laryngeal cancer cells (Hep-2). J. Photochem. Photobiol. B Biol. 2019, 201, 111624. [Google Scholar] [CrossRef] [PubMed]
- Emri, E.; Miko, E.; Bai, P.; Boros, G.; Nagy, G.; Rózsa, D.; Juhász, T.; Hegeds, C.; Horkay, I.; Remenyik, É.; et al. Effects of non-toxic zinc exposure on human epidermal keratinocytes. Metallomics 2015, 7, 499–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamore, S.D.; Cabello, C.M.; Wondrak, G.T. The topical antimicrobial zinc pyrithione is a heat shock response inducer that causes DNA damage and PARP-dependent energy crisis in human skin cells. Cell Stress Chaperones 2010, 15, 309–322. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Pie, J.E.; Kim, Y.R.; Lee, H.R.; Son, S.W.; Kim, M.K. Effects of zinc oxide nanoparticles on gene expression profile in human keratinocytes. Mol. Cell. Toxicol. 2012, 8, 113–118. [Google Scholar] [CrossRef]
- Sato, R.; Yamamoto, H.; Kasai, K.; Yamauchi, M. Distribution pattern of versican, link protein and hyaluronic acid in the rat periodontal ligament during experimental tooth movement. J. Periodontal Res. 2002, 37, 15–22. [Google Scholar] [CrossRef]
- Zimmermann, D.R.; Dours-Zimmermann, M.T.; Schubert, M.; Bruckner-Tuderman, L. Versican is expressed in the proliferating zone in the epidermis and in association with the elastic network of the dermis. J. Cell Biol. 1994, 124, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Beikler, T.; Peters, U.; Prior, K.; Eisenacher, M.; Flemmig, T.F. Gene expression in periodontal tissues following treatment. BMC Med. Genom. 2008, 1, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, J.J.; Harris, T.J.C. Cadherin Trafficking for Tissue Morphogenesis: Control and Consequences. Traffic 2016, 17, 1233–1243. [Google Scholar] [CrossRef] [Green Version]
- Hung, C.F.; Chiang, H.S.; Lo, H.M.; Jian, J.S.; Wu, W. Bin E-cadherin and its downstream catenins are proteolytically cleaved in human HaCaT keratinocytes exposed to UVB. Exp. Dermatol. 2006, 15, 315–321. [Google Scholar] [CrossRef]
- Yeh, J.T.; Yeh, L.K.; Jung, S.M.; Chang, T.J.; Wu, H.H.; Shiu, T.F.; Liu, C.Y.; Kao, W.W.Y.; Chu, P.H. Impaired skin wound healing in lumican-null mice. Br. J. Dermatol. 2010, 163, 1174–1180. [Google Scholar] [CrossRef]
- Niewiarowska, J.; Brézillon, S.; Sacewicz-Hofman, I.; Bednarek, R.; Maquart, F.X.; Malinowski, M.; Wiktorska, M.; Wegrowski, Y.; Cierniewski, C.S. Lumican inhibits angiogenesis by interfering with α2β1 receptor activity and downregulating MMP-14 expression. Thromb. Res. 2011, 128, 452–457. [Google Scholar] [CrossRef]
- Liu, X.J.; Kong, F.Z.; Wang, Y.H.; Zheng, J.H.; Wan, W.D.; Deng, C.L.; Mao, G.Y.; Li, J.; Yang, X.M.; Zhang, Y.L.; et al. Lumican Accelerates Wound Healing by Enhancing α2β1 Integrin-Mediated Fibroblast Contractility. PLoS ONE 2013, 8, e67124. [Google Scholar] [CrossRef] [PubMed]
- Handford, P.A. Fibrillin-1, a calcium binding protein of extracellular matrix. Biochim. Biophys. Acta BBA Mol. Cell Res. 2000, 1498, 84–90. [Google Scholar] [CrossRef] [Green Version]
- Adamo, C.S.; Zuk, A.V.; Sengle, G. The fibrillin microfibril/elastic fibre network: A critical extracellular supramolecular scaffold to balance skin homoeostasis. Exp. Dermatol. 2021, 30, 25–37. [Google Scholar] [CrossRef]
- Philips, N.; Samuel, M.; Arena, R.; Chen, Y.J.; Conte, J.; Natrajan, P.; Haas, G.; Gonzalez, S. Direct inhibition of elastase and matrixmetalloproteinases and stimulation of biosynthesis of fibrillar collagens, elastin, and fibrillins by xanthohumol. J. Cosmet. Sci. 2010, 61, 125–132. [Google Scholar] [CrossRef]
- Dzamba, B.J.; Keene, D.R.; Isogai, Z.; Charbonneau, N.L.; Karaman-Jurukovska, N.; Simon, M.; Sakai, L.Y. Assembly of epithelial cell fibrillins. J. Investig. Dermatol. 2001, 117, 1612–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masaki, H. Role of antioxidants in the skin: Anti-aging effects. J. Dermatol. Sci. 2010, 58, 85–90. [Google Scholar] [CrossRef]
- Kruk, J.; Duchnik, E. Oxidative stress and skin diseases: Possible role of physical activity. Asian Pac. J. Cancer Prev. 2014, 15, 561–568. [Google Scholar] [CrossRef]
- Butt, H.; Mehmood, A.; Ali, M.; Tasneem, S.; Tarar, M.N.; Riazuddin, S. Vitamin E preconditioning alleviates in vitro thermal stress in cultured human epidermal keratinocytes. Life Sci. 2019, 239, 116972. [Google Scholar] [CrossRef]
- Calcabrini, C.; De Bellis, R.; Mancini, U.; Cucchiarini, L.; Potenza, L.; De Sanctis, R.; Patrone, V.; Scesa, C.; Dachà, M. Rhodiola rosea ability to enrich cellular antioxidant defences of cultured human keratinocytes. Arch. Dermatol. Res. 2010, 302, 191–200. [Google Scholar] [CrossRef]
- Casares, L.; García, V.; Garrido-Rodríguez, M.; Millán, E.; Collado, J.A.; García-Martín, A.; Peñarando, J.; Calzado, M.A.; de la Vega, L.; Muñoz, E. Cannabidiol induces antioxidant pathways in keratinocytes by targeting BACH1. Redox Biol. 2020, 28, 101321. [Google Scholar] [CrossRef]
- Salucci, S.; Burattini, S.; Buontempo, F.; Martelli, A.M.; Falcieri, E.; Battistelli, M. Protective effect of different antioxidant agents in UVB-irradiated keratinocytes. Eur. J. Histochem. 2017, 61, 215–221. [Google Scholar] [CrossRef]
- Son, D.H.; Yang, D.J.; Sun, J.S.; Kim, S.K.; Kang, N.; Kang, J.Y.; Choi, Y.H.; Lee, J.H.; Moh, S.H.; Shin, D.M.; et al. A novel peptide, nicotinyl–isoleucine–valine–histidine (Na–IVH), promotes antioxidant gene expression and wound healing in HaCaT cells. Mar. Drugs 2018, 16, 262. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Venkataraman, S.; Coleman, M.C.; Spitz, D.R.; Wertz, P.W.; Domann, F.E. Glutathione peroxidase-1 inhibits UVA-induced AP-2α expression in human keratinocytes. Biochem. Biophys. Res. Commun. 2006, 351, 1066–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezvani, H.R.; Mazurier, F.; Cario-André, M.; Pain, C.; Ged, C.; Taïeb, A.; De Verneuil, H. Protective effects of catalase overexpression on UVB-induced apoptosis in normal human keratinocytes. J. Biol. Chem. 2006, 281, 17999–18007. [Google Scholar] [CrossRef] [Green Version]
- Miyachi, Y. Photoaging from an oxidative standpoint. J. Dermatol. Sci. 1995, 9, 79–86. [Google Scholar] [CrossRef]
- Dahle, J.; Kvam, E.; Stokke, T. Bystander effects in UV-induced genomic instability: Antioxidants inhibit delayed mutagenesis induced by ultraviolet A and B radiation. J. Carcinog. 2005, 4, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masaki, H.; Izutsu, Y.; Yahagi, S.; Okano, Y. Reactive oxygen species in HaCaT keratinocytes after UVB irradiation are triggered by intracellular Ca2+ levels. J. Investig. Dermatol. Symp. Proc. 2009, 14, 50–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluemsamran, T.; Onkoksoong, T.; Panich, U. Caffeic acid and ferulic acid inhibit UVA-induced matrix metalloproteinase-1 through regulation of antioxidant defense system in keratinocyte HaCaT cells. In Proceedings of the Photochemistry and Photobiology. Photochem. Photobiol. 2012, 88, 961–968. [Google Scholar] [CrossRef]
- Wu, F.; Cui, L. Resveratrol suppresses melanoma by inhibiting NF-κB/miR-221 and inducing TFG expression. Arch. Dermatol. Res. 2017, 309, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K. Preservation effects of geniposidic acid on human keratinocytes (HaCaT) against UVB. Biomed. Dermatol. 2018, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Lim, K.H.; Ku, J.-E.; Rhie, S.-J.; Ryu, J.Y.; Bae, S.; Kim, Y.-S. Anti-oxidant and Anti-inflammatory Effects of Sinapic Acid in UVB Irradiation-Damaged HaCaT Keratinocytes. Asian J. Beauty Cosmetol. 2017, 15, 513–522. [Google Scholar] [CrossRef]
- Salesa, B.; Serrano-Aroca, Á. Multi-Layer Graphene Oxide in Human Keratinocytes: Time-Dependent Cytotoxicity, Proliferation, and Gene Expression. Coatings 2021, 11, 414. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).