
Characterizations of Hamster Retina as a Model for Studies of Retinal Cholesterol Homeostasis

 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Study Design
2.3. Anesthesia
2.4. Genotyping
2.5. In Vivo Imaging
2.6. The Axial Eye Length Measurements by SD-OCT
2.7. Blood Glucose
2.8. Euthanasia
2.9. Blood Hemoglobin A1c (HbA1c)
2.10. Serum Lipids
2.11. Sterol Profiling
2.12. Histo- and Immunohistochemistry
2.13. Statistical Analysis
3. Results
3.1. Genotyping for the Mutations That Underlie Rd1 and Rd8
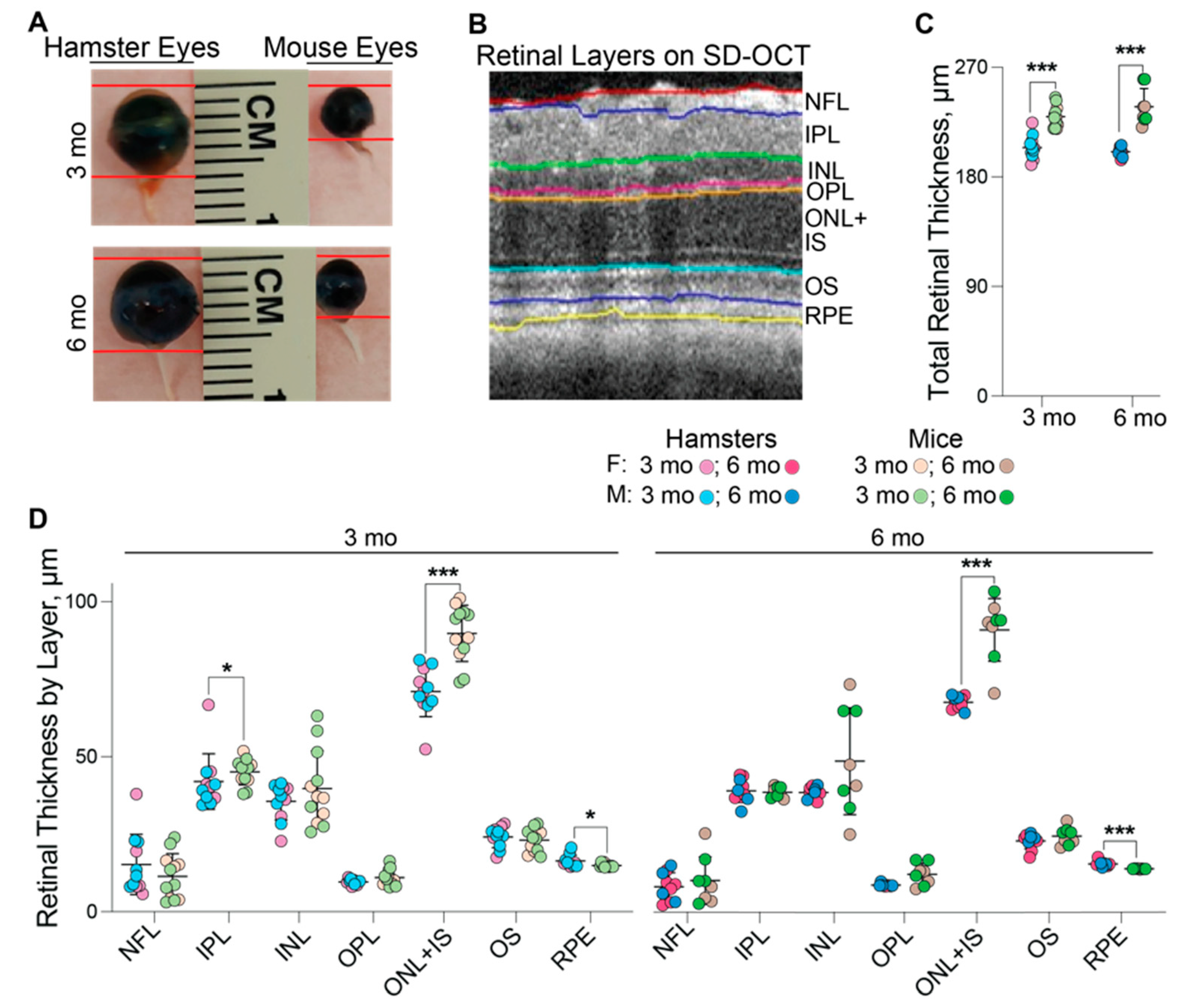
3.2. Ocular Biometric Assessments
3.3. In Vivo Imaging of the Retina and Area Centralis
3.4. Hamster Retinal Abnormalities
3.5. Blood Glucose and Glycemic Control
3.6. Serum Lipids
3.7. Retinal Sterol Profiling
3.8. Retinal Cholesterol Distribution
4. Discussion
5. Patents
Institutional Animal Care and Use Committee
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Clifford, C.B.; Simmons, J.H. The Clinical Chemistry of Laboratory Animals, 3rd ed.; CRC Press, Taylor and Francis: Boca Raton, FL, USA, 2017. [Google Scholar]
- Fliesler, S.J.; Bretillon, L. The ins and outs of cholesterol in the vertebrate retina. J. Lipid Res. 2010, 51, 3399–3413. [Google Scholar] [CrossRef]
- Pikuleva, I.A.; Curcio, C.A. Cholesterol in the retina: The best is yet to come. Prog. Retin. Eye Res. 2014, 41, 64–89. [Google Scholar] [CrossRef]
- Singh, V.; Tiwari, R.L.; Dikshit, M.; Barthwal, M.K. Models to study atherosclerosis: A mechanistic insight. Curr. Vasc. Pharmacol. 2009, 7, 75–109. [Google Scholar] [CrossRef]
- Ha, Y.C.; Barter, P.J. Differences in plasma cholesteryl ester transfer activity in sixteen vertebrate species. Comp. Biochem. Physiol. B 1982, 71, 265–269. [Google Scholar] [CrossRef]
- Spady, D.K.; Woollett, L.A.; Dietschy, J.M. Regulation of plasma LDL-cholesterol levels by dietary cholesterol and fatty acids. Annu. Rev. Nutr. 1993, 13, 355–381. [Google Scholar] [CrossRef]
- Tall, A. Plasma lipid transfer proteins. Annu. Rev. Biochem. 1995, 64, 235–257. [Google Scholar] [CrossRef]
- Reaves, S.K.; Wu, J.Y.J.; Wu, Y.; Fanzo, J.; Wang, Y.R.; Lei, P.P.; Lei, K.Y. Regulation of intestinal apolipoprotein B mRNA editing levels by a Zinc-deficient diet and cDNA cloning of editing protein in hamsters. J. Nutr. 2000, 130, 2166–2173. [Google Scholar] [CrossRef]
- Jeon, H.; Blacklow, S.C. Structure and physiologic function of the low-density lipoprotein receptor. Annu. Rev. Biochem. 2005, 74, 535–562. [Google Scholar] [CrossRef]
- Marsh, J.B.; Welty, F.K.; Lichtenstein, A.H.; Lamon-Fava, S.; Schaefer, E.J. Apolipoprotein B metabolism in humans: Studies with stable isotope-labeled amino acid precursors. Atherosclerosis 2002, 162, 227–244. [Google Scholar] [CrossRef]
- Dietschy, J.M.; Turley, S.D. Control of cholesterol turnover in the mouse. J. Biol. Chem. 2002, 277, 3801–3804. [Google Scholar] [CrossRef]
- Horton, J.D.; Cuthbert, J.A.; Spady, D.K. Regulation of hepatic 7α-hydroxylase expression and response to dietary cholesterol in the rat and hamster. J. Biol. Chem. 1995, 270, 5381–5387. [Google Scholar] [CrossRef]
- De Fabiani, E.; Crestani, M.; Marrapodi, M.; Pinelli, A.; Chiang, J.Y.; Galli, G. Regulation of the hamster cholesterol 7 alpha-hydroxylase gene (CYP7A): Prevalence of negative over positive transcriptional control. Biochem. Biophys. Res. Commun. 1996, 226, 663–671. [Google Scholar] [CrossRef]
- Russell, D.W. Fifty years of advances in bile acid synthesis and metabolism. J. Lipid Res. 2009, 50, S120–S125. [Google Scholar] [CrossRef]
- Fliesler, S.J.; Florman, R.; Rapp, L.M.; Pittler, S.; Keller, R. In vivo biosynthesis of cholesterol in the rat retina. FEBS Lett. 1993, 335, 234–238. [Google Scholar] [CrossRef]
- Fliesler, S.; Keller, R. Metabolism of (3H) farnesol to cholesterol and cholesterogenic intermediates in the living rat eye. Biochem. Biophys. Res. Commun. 1995, 210, 695–702. [Google Scholar] [CrossRef]
- Fliesler, S.J.; Keller, R.K. Isoprenoid metabolism in the vertebrate retina. Int. J. Biochem. Cell Biol. 1997, 29, 877–894. [Google Scholar] [CrossRef]
- Elner, V. Retinal pigment epithelial acid lipase activity and lipoprotein receptors: Effects of dietary omega-3 fatty acids. Trans. Am. Ophthalmol. Soc. 2002, 100, 301–338. [Google Scholar]
- Tserentsoodol, N.; Sztein, J.M.; Campos, M.; Gordiyenko, N.V.; Fariss, R.N.; Lee, J.W.; Fliesler, S.J.; Rodriguez, I.R. Uptake of cholesterol by the retina occurs primarily via a low density lipoprotein receptor-mediated process. Mol. Vis. 2006, 12, 1306–1318. [Google Scholar]
- Duncan, K.G.; Hosseini, K.; Bailey, K.R.; Yang, H.; Lowe, R.J.; Matthes, M.T.; Kane, J.P.; Lavail, M.M.; Schwartz, D.M.; Duncan, J.L. Expression of reverse cholesterol transport proteins ATP-binding cassette A1 (ABCA1) and scavenger receptor BI (SR-BI) in the retina and retinal pigment epithelium. Br. J. Ophthalmol. 2009, 93, 1116–1120. [Google Scholar] [CrossRef]
- Zheng, W.; Reem, R.E.; Omarova, S.; Huang, S.; DiPatre, P.L.; Charvet, C.D.; Curcio, C.A.; Pikuleva, I.A. Spatial distribution of the pathways of cholesterol homeostasis in human retina. PLoS ONE 2012, 7, e37926. [Google Scholar] [CrossRef]
- Lin, J.B.; Mast, N.; Bederman, I.R.; Li, Y.; Brunengraber, H.; Björkhem, I.; Pikuleva, I.A. Cholesterol in mouse retina originates primarily from in situ de novo biosynthesis. J. Lipid Res. 2016, 57, 258–264. [Google Scholar] [CrossRef]
- Curcio, A.C.; Johnson, M.; Rudolf, M.; Huang, J.-D. The oil spill in ageing Bruch membrane. Br. J. Ophthalmol. 2011, 95, 1638–1645. [Google Scholar] [CrossRef]
- Miller, J.W. Age-related macular degeneration revisited—Piecing the puzzle: The LXIX Edward Jackson memorial lecture. Am. J. Ophthalmol. 2013, 155, 1–35.e13. [Google Scholar] [CrossRef] [PubMed]
- Hammer, S.S.; Busik, J.V. The role of dyslipidemia in diabetic retinopathy. Vis. Res. 2017, 139, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, E.M.; Emri, E.; Merle, B.M.J.; Colijn, J.M.; Kersten, E.; Cougnard-Gregoire, A.; Dammeier, S.; Meester-Smoor, M.; Pool, F.M.; de Jong, E.K.; et al. A new perspective on lipid research in age-related macular degeneration. Prog. Retin. Eye Res. 2018, 67, 56–86. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Sengupta, P. Age of laboratory hamster and human: Drawing the connexion. Biomed. Pharmacol. J. 2019, 12, 49–56. [Google Scholar] [CrossRef]
- Dutta, S.; Sengupta, P. Men and mice: Relating their ages. Life Sci. 2016, 152, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Omarova, S.; Charvet, C.D.; Reem, R.E.; Mast, N.; Zheng, W.; Huang, S.; Peachey, N.S.; Pikuleva, I.A. Abnormal vascularization in mouse retina with dysregulated retinal cholesterol homeostasis. J. Clin. Investig. 2012, 122, 3012–3023. [Google Scholar] [CrossRef] [PubMed]
- Saadane, A.; Mast, N.; Charvet, C.D.; Omarova, S.; Zheng, W.; Huang, S.S.; Kern, T.S.; Peachey, N.S.; Pikuleva, I.A. Retinal and nonocular abnormalities in Cyp27a1(-/-) Cyp46a1 (-/-) mice with dysfunctional metabolism of cholesterol. Am. J. Pathol. 2014, 184, 2403–2419. [Google Scholar] [CrossRef]
- Mast, N.; Shafaati, M.; Zaman, W.; Zheng, W.; Prusak, D.; Wood, T.; Ansari, G.A.S.; Lövgren-Sandblom, A.; Olin, M.; Björkhem, I.; et al. Marked variability in hepatic expression of cytochromes CYP7A1 and CYP27A1 as compared to cerebral CYP46A1. Lessons from a dietary study with omega 3 fatty acids in hamsters. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2010, 1801, 674–681. [Google Scholar] [CrossRef]
- Mast, N.; Reem, R.; Bederman, I.; Huang, S.; DiPatre, P.L.; Björkhem, I.; Pikuleva, I. Cholestenoic acid is an important elimination product of cholesterol in the retina: Comparison of retinal cholesterol metabolism with that in the brain. Investig. Opthalmol. Vis. Sci. 2011, 52, 594–603. [Google Scholar] [CrossRef] [PubMed]
- Mast, N.; El-Darzi, N.; Petrov, A.M.; Li, Y.; Pikuleva, I.A. CYP46A1-dependent and independent effects of efavirenz treatment. Brain Commun. 2020, 2, fcaa180. [Google Scholar] [CrossRef] [PubMed]
- Saadane, A.; Mast, N.; Trichonas, G.; Chakraborty, D.; Hammer, S.; Busik, J.V.; Grant, M.B.; Pikuleva, I.A. Retinal vascular abnormalities and microglia activation in mice with deficiency in cytochrome P450 46A1—Mediated cholesterol removal. Am. J. Pathol. 2019, 189, 405–425. [Google Scholar] [CrossRef] [PubMed]
- Chang, B.; Hurd, R.; Wang, J.; Nishina, P. Survey of common eye diseases in laboratory mouse strains. Investig. Opthalmol. Vis. Sci. 2013, 54, 4974–4981. [Google Scholar] [CrossRef]
- Tiao, Y.C.; Blakemore, C. Regional specialization in the golden hamster′s retina. J. Comp. Neurol. 1976, 168, 439–457. [Google Scholar]
- Nadal-Nicolás, F.M.; Jiménez-López, M.; Sobrado-Calvo, P.; Nieto-López, L.; Cánovas-Martínez, I.; Salinas-Navarro, M.; Vidal-Sanz, M.; Agudo, M. Brn3a as a marker of retinal ganglion cells: Qualitative and quantitative time course studies in naive and optic nerve-injured retinas. Invest. Ophthalmol. Vis. Sci. 2009, 50, 3860–3868. [Google Scholar] [CrossRef]
- Xiang, M.; Zhou, L.; Macke, J.; Yoshioka, T.; Hendry, S.; Eddy, R.; Shows, T.; Nathans, J. The Brn-3 family of POU-domain factors: Primary structure, binding specificity, and expression in subsets of retinal ganglion cells and somatosensory neurons. J. Neurosci. 1995, 15, 4762–4785. [Google Scholar] [CrossRef]
- Staurenghi, G.; Sadda, S.; Chakravarthy, U.; Spaide, R.F. International nomenclature for optical coherence tomography P: Pro-posed lexicon for anatomic landmarks in normal posterior segment spectral-domain optical coherence tomography: The IN*OCT consensus. Ophthalmology 2014, 121, 1572–1578. [Google Scholar] [CrossRef]
- Mehalow, A.K.; Kameya, S.; Smith, R.S.; Hawes, N.L.; Denegre, J.M.; Young, J.A.; Bechtold, L.; Haider, N.B.; Tepass, U.; Heckenlively, J.R.; et al. CRB1 is essential for external limiting membrane integrity and photoreceptor morphogenesis in the mammalian retina. Hum. Mol. Genet. 2003, 12, 2179–2189. [Google Scholar] [CrossRef]
- Radu, M.; Chernoff, J. An in vivo assay to test blood vessel permeability. J. Vis. Exp. 2013, 73, e50062. [Google Scholar] [CrossRef]
- Campos, M.; Amaral, J.; Becerra, S.P.; Fariss, R.N. A novel imaging technique for experimental choroidal neovascularization. Invest. Ophthalmol. Vis. Sci. 2006, 47, 5163–5170. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Stitt, A.W.; Curtis, T.; Chen, M.; Medina, R.J.; McKay, G.; Jenkins, A.; Gardiner, T.A.; Lyons, T.J.; Hammes, H.-P.; Simó, R.; et al. The progress in understanding and treatment of diabetic retinopathy. Prog. Retin. Eye Res. 2016, 51, 156–186. [Google Scholar] [CrossRef] [PubMed]
- Hammes, H.P.; Wellensiek, B.; Klöting, I.; Sickel, E.; Bretzel, R.G.; Brownlee, M. The relationship of glycaemic level to advanced gly-cation end-product (AGE) accumulation and retinal pathology in the spontaneous diabetic hamster. Diabetologia 1998, 41, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Gerritsen, G.C. The Chinese hamster as a model for the study of diabetes mellitus. Diabetes 1982, 31, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Wolford, S.T.; Schroer, R.A.; Gohs, F.X.; Gallo, P.P.; Brodeck, M.; Falk, H.B.; Ruhren, R. Reference range data base for serum chemistry and hematology values in laboratory animals. J. Toxicol. Environ. Heal. Part. A 1986, 18, 161–188. [Google Scholar] [CrossRef]
- Nieweg, K.; Schaller, H.; Pfrieger, F.W. Marked differences in cholesterol synthesis between neurons and glial cells from postnatal rats. J. Neurochem. 2009, 109, 125–134. [Google Scholar] [CrossRef]
- Mast, N.; Bederman, I.R.; Pikuleva, I.A. Retinal cholesterol content is reduced in simvastatin-treated mice due to inhibited local biosynthesis albeit increased uptake of serum cholesterol. Drug Metab. Dispos. 2018, 46, 1528–1537. [Google Scholar] [CrossRef] [PubMed]
- Pikuleva, I.A. Cytochrome P450s and cholesterol homeostasis. Pharmacol. Ther. 2006, 112, 761–773. [Google Scholar] [CrossRef]
- El-Darzi, N.; Mast, N.; Petrov, A.M.; Dao, T.; Astafev, A.A.; Saadane, A.; Prendergast, E.; Schwarz, E.; Bederman, I.; Pikuleva, I.A. Studies of ApoD (-/-) and ApoD (-/-)ApoE (-/-) mice uncover the APOD significance for retinal metabolism, function, and status of chorioretinal blood vessels. Cell Mol. Life Sci. 2021, 78, 963–983. [Google Scholar] [CrossRef]
- Saadane, A.; Petrov, A.; Mast, N.; El-Darzi, N.; Dao, T.; Alnemri, A.; Song, Y.; Dunaief, J.L.; Pikuleva, I.A. Mechanisms that minimize retinal impact of apolipoprotein E absence. J. Lipid Res. 2018, 59, 2368–2382. [Google Scholar] [CrossRef]
- Curcio, C.A.; Rudolf, M.; Wang, L. Histochemistry and lipid profiling combine for insights into aging and age-related maculopathy. Adv. Struct. Saf. Stud. 2009, 580, 267–281. [Google Scholar] [CrossRef]
- Den Hollander, A.I.; Davis, J.; van der Velde-Visser, S.D.; Zonneveld, M.N.; Pierrottet, C.O.; Koenekoop, R.K.; Kellner, U.; van den Born, L.I.; Heckenlively, J.R.; Hoyng, C.B.; et al. CRB1 mutation spectrum in inherited retinal dystro-phies. Hum. Mutat. 2004, 24, 355–369. [Google Scholar] [CrossRef] [PubMed]
- Chang, A. Biochemical abnormalities in the chinese hamster (Cricetulus griseus) with spontaneous diabetes. Int. J. Biochem. 1981, 13, 41–43. [Google Scholar] [CrossRef]
- Yu, S.-H.; Kim, H.J.; Lee, K.-P.; Lee, E.-S.; Lee, J.-Y.; Jeon, C.-J. Populations of rod and cone photoreceptors in the hamster retina. Appl. Microsc. 2009, 39, 291–299. [Google Scholar]
- Volland, S.; Esteve-Rudd, J.; Hoo, J.; Yee, C.; Williams, D.S. A comparison of some organizational characteristics of the mouse central retina and the human macula. PLoS ONE 2015, 10, e0125631. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biometric Assessment | Hamsters | Mice | ||
|---|---|---|---|---|
| 3-Month-Old | 6-Month-Old | 3-Month-Old | 6-Month-Old | |
| Axial eye length by ruler (mm) | 5.9 ± 0.1 a (3F + 3M) | 6.0 ± 0.0 a (3F + 3M) | 3.2 ± 0.1 (4F + 3M) | 3.5 ± 0.3 (4F + 3M) |
| Axial eye length by SD-OCT (mm) | 6.0 ± 0.1 (1F + 2M) | 6.1 ± 0.1 (3F + 3M) | ND | ND |
| Retina wet weight (mg) | 6.9 ± 0.5 a (5F + 5M) | 7.7 ± 0.9 a (7F + 6M) | 2.3 ± 0.1 (9F +10M) | 2.4 ± 0.1 (5F + 7M) |
| Total thickness of the retinal cross section by SD-OCT (µm) | 204.2 ± 9.7 a (5F + 6M) | 200.7 ± 3.7 a (6F + 4M) | 229.4 ± 8.8 (5F + 7M) | 237.6 ± 15.0 (4F + 4M) |
| Retinal Sterol or Sterol/Cholesterol Ratio | Hamsters, 3-mo (5F + 5M) | Hamsters, 6-mo (5F + 5M) | Mice * 6-mo (6F + 6M) | Humans ** (3M) | Change Factor Relevant to Humans: 6-mo Hamsters; 6-mo Mice |
|---|---|---|---|---|---|
| Cholesterol, nmol/mg protein | 86 ± 3 | 87 ± 4 | 35 ± 4 | 49 ± 7 | 1.78; 0.71 |
| Lathosterol, pmol/mg protein | 367 ± 62 | 115 ± 17 | 85 ± 6 | NM | NA |
| Desmosterol, pmol/mg protein | 310 ± 58 | 45± 8 | 54 ± 4 | NM | NA |
| Pregnenolone, pmol/mg protein | 124 ± 38 | 15 ± 3 | 25 ± 6 | 2.7 ± 0.8 | 5.55; 9.26 |
| 27HC, pmol/mg protein | 127 ± 21 | ND | ND | ND | NA |
| 27COOH, pmol/mg protein | 238 ± 46 | 94 ± 9 | 7.7 ± 0.9 | 79 ± 44 | 1.19; 0.10 |
| 24HC, pmol/mg protein | ND | ND | 6.1 ± 1.0 | 2.4 ± 1.7 | NA; 2.54 |
| Lathosterol/cholesterol, pmol/nmol | 4.3 | 1.3 | 2.5 | NM | NA |
| Desmosterol/cholesterol, pmol/nmol | 3.6 | 0.5 | 1.6 | NM | NA |
| Pregnenolone/cholesterol pmol/nmol | 1.4 | 0.17 | 0.84 | 0.06 | 2.83; 14 |
| 27HC/cholesterol, pmol/nmol | 1.5 | ND | ND | ND | NA |
| 27COOH/cholesterol, pmol/nmol | 2.8 | 1.1 | 0.22 | 1.6 | 0.69; 0.14 |
| 24HC/cholesterol, pmol/nmol | ND | ND | 0.20 | 0.05 | NA; 4.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Darzi, N.; Mast, N.; Dailey, B.; Denker, J.; Li, Y.; Vance, J.; Pikuleva, I.A. Characterizations of Hamster Retina as a Model for Studies of Retinal Cholesterol Homeostasis. Biology 2021, 10, 1003. https://doi.org/10.3390/biology10101003
El-Darzi N, Mast N, Dailey B, Denker J, Li Y, Vance J, Pikuleva IA. Characterizations of Hamster Retina as a Model for Studies of Retinal Cholesterol Homeostasis. Biology. 2021; 10(10):1003. https://doi.org/10.3390/biology10101003
Chicago/Turabian StyleEl-Darzi, Nicole, Natalia Mast, Brian Dailey, John Denker, Yong Li, Joseph Vance, and Irina A. Pikuleva. 2021. "Characterizations of Hamster Retina as a Model for Studies of Retinal Cholesterol Homeostasis" Biology 10, no. 10: 1003. https://doi.org/10.3390/biology10101003
APA StyleEl-Darzi, N., Mast, N., Dailey, B., Denker, J., Li, Y., Vance, J., & Pikuleva, I. A. (2021). Characterizations of Hamster Retina as a Model for Studies of Retinal Cholesterol Homeostasis. Biology, 10(10), 1003. https://doi.org/10.3390/biology10101003