Does KRAS Play a Role in the Regulation of Colon Cancer Cells-Derived Exosomes?



 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Viability Assay
2.3. Detection of Exosome Formation in Colon Cancer Cells
2.4. Membrane-Dye Labeling of Cells and Exosomes
2.5. Exosome Isolation
2.6. Transmission Electron Microscopy
2.7. Particle Size Analysis
2.8. Western Blot Analysis
2.9. Protein Lysis and Digestion
2.10. Nano-Liquid Chromatography-Mass Spectrometry/Mass Spectrometry
2.11. Protein Identification and Network Analysis
2.12. Imaging Flow Cytometry
2.13. Statistical Analysis
3. Results
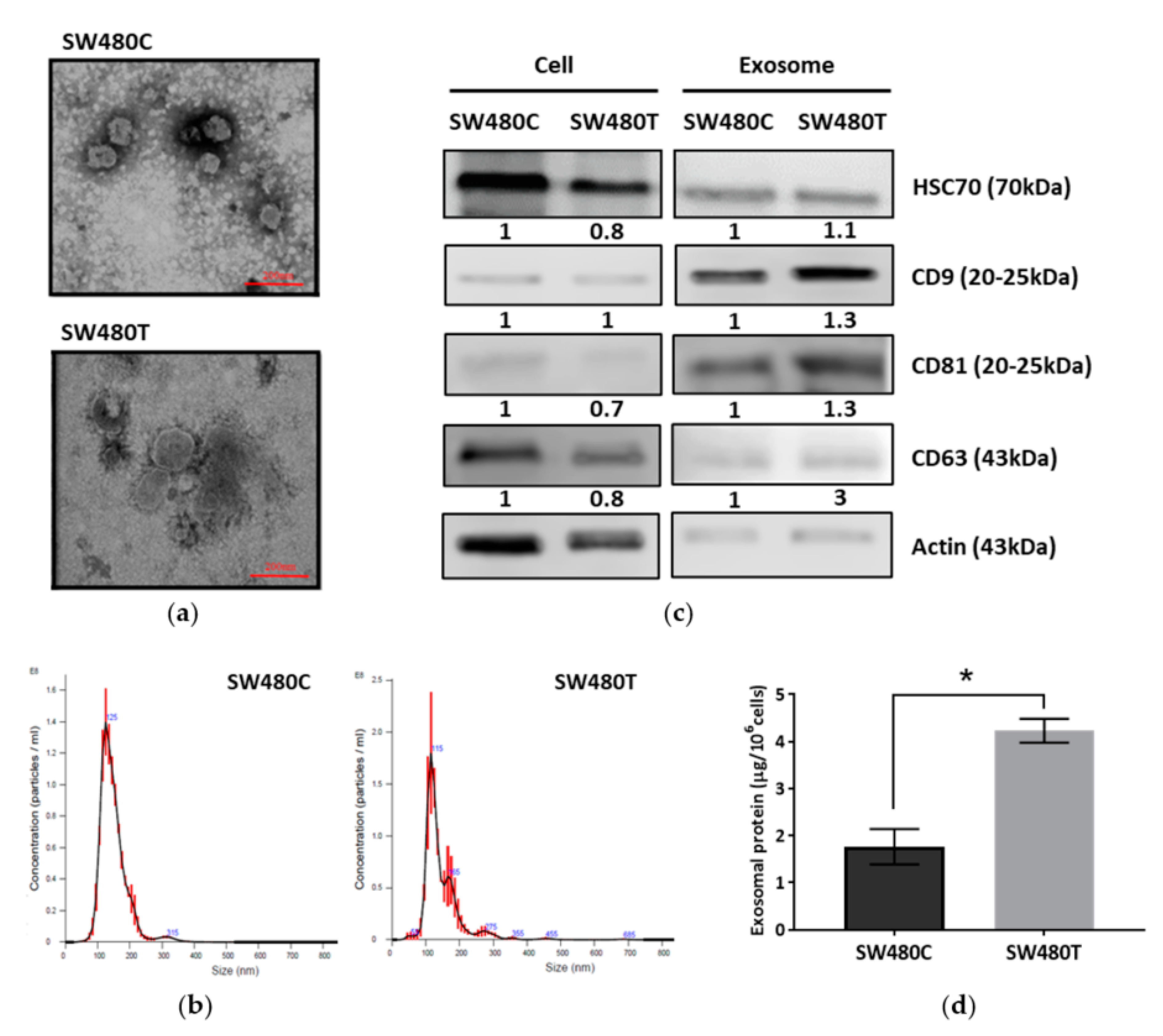
3.1. Detection of Exosome Formation in Colon Cancer Cells
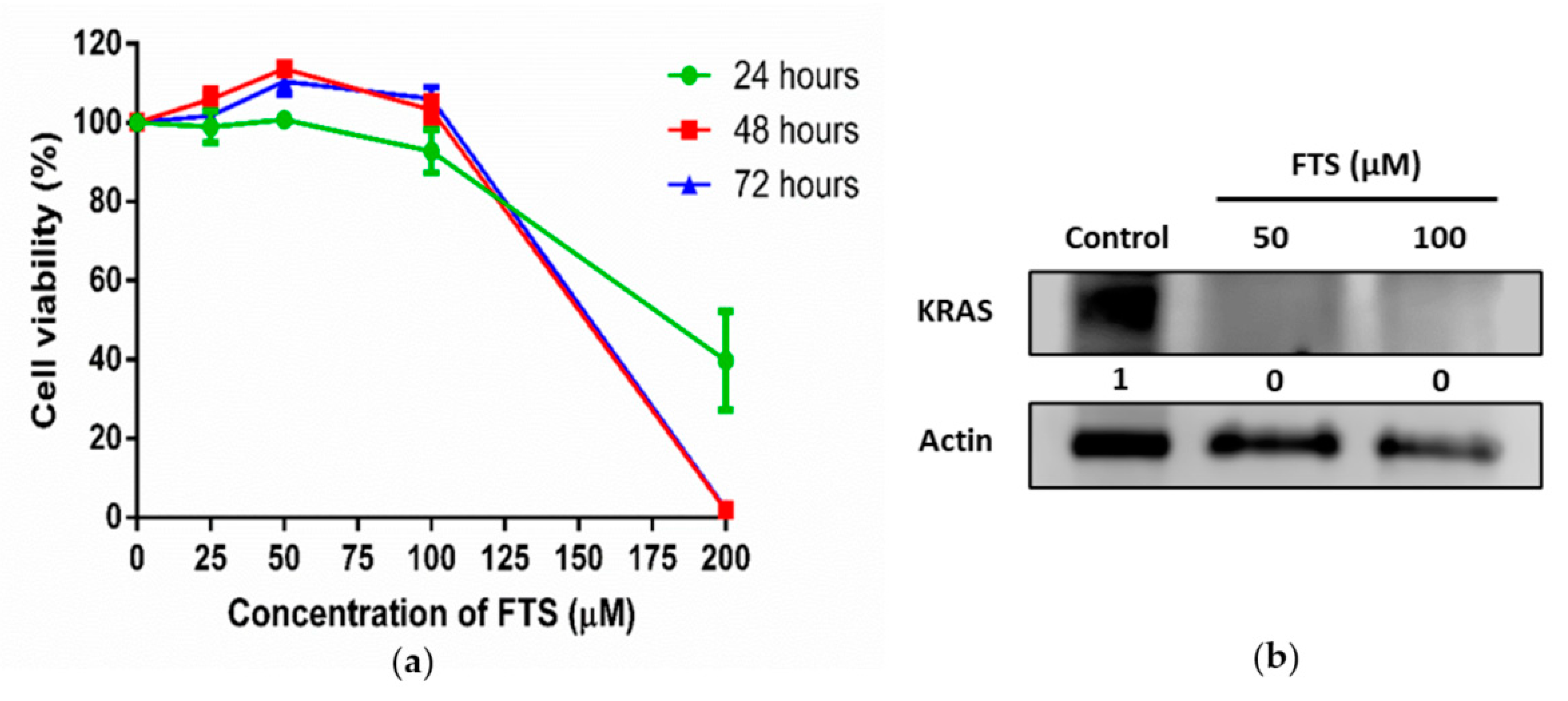
3.2. Inhibition of KRAS Signaling in SW480 Cells
3.3. Isolation and Characterization of Exosomes
3.4. Proteomic Analysis of SW480 Cells-Derived Exosomes
3.4.1. Protein Identification and Network Analysis
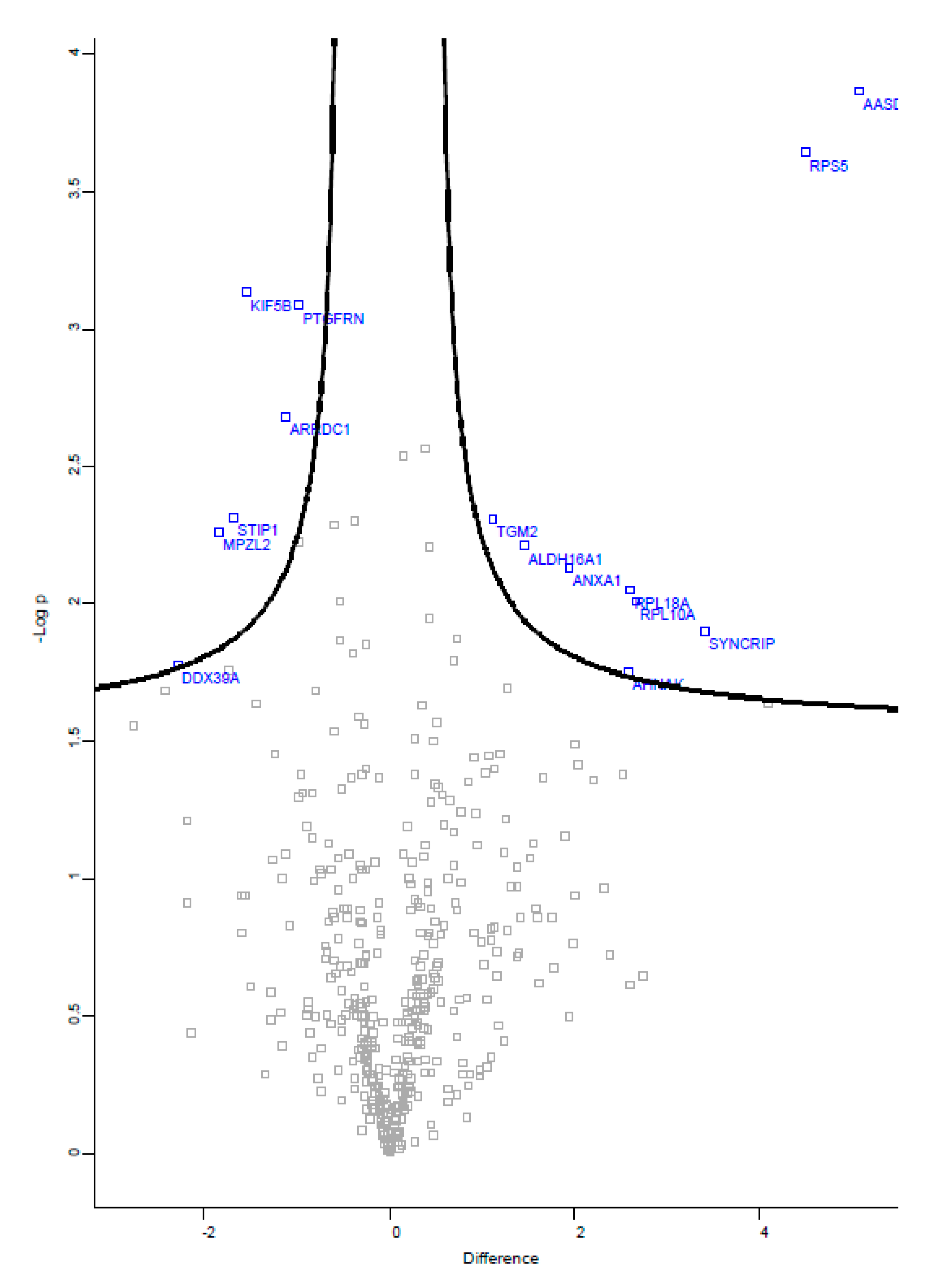
3.4.2. Comparative Proteomics
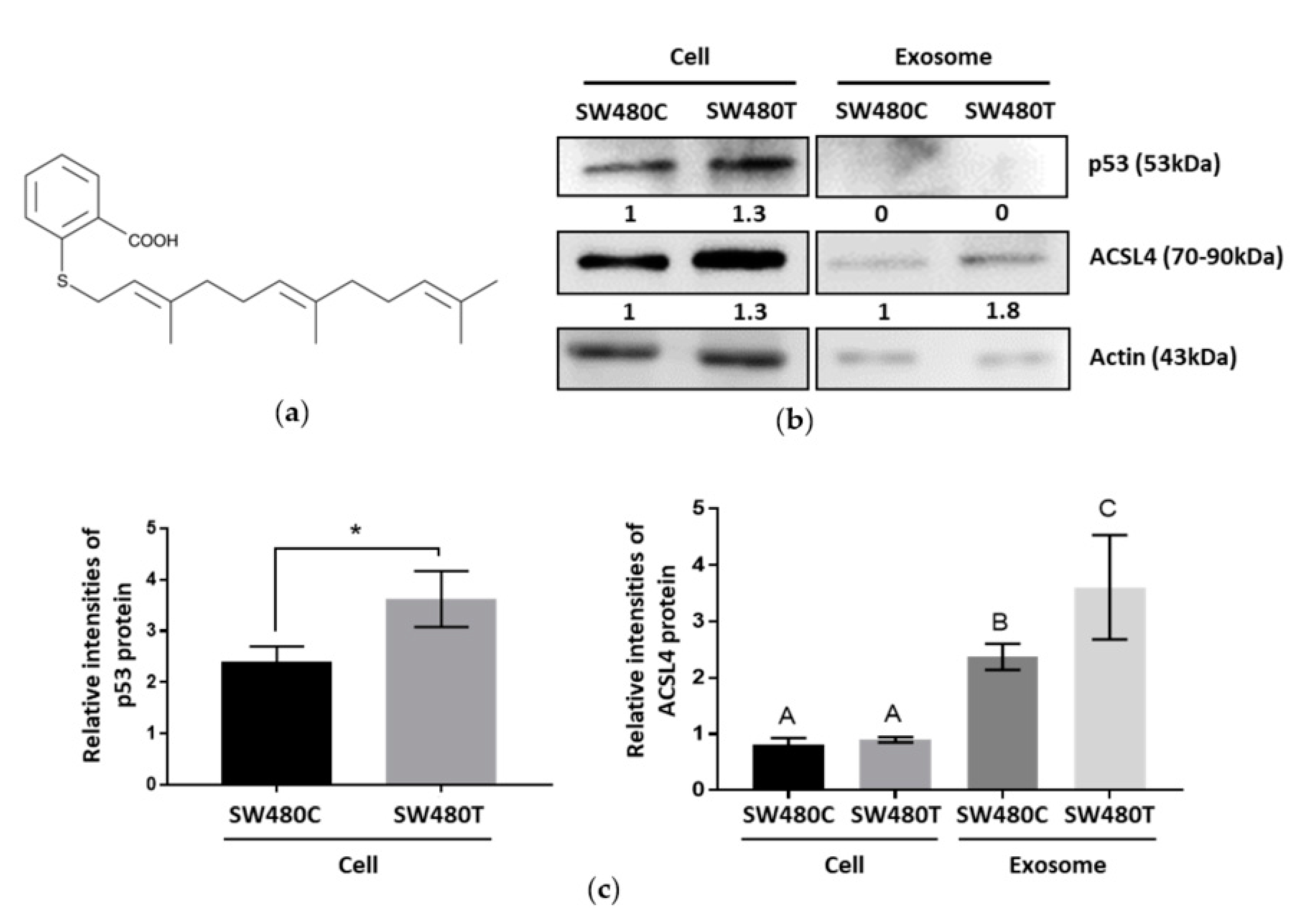
3.4.3. Verification
3.5. Effect of SW480 Cells-Derived Exosomes on Survival of Recipient Cells
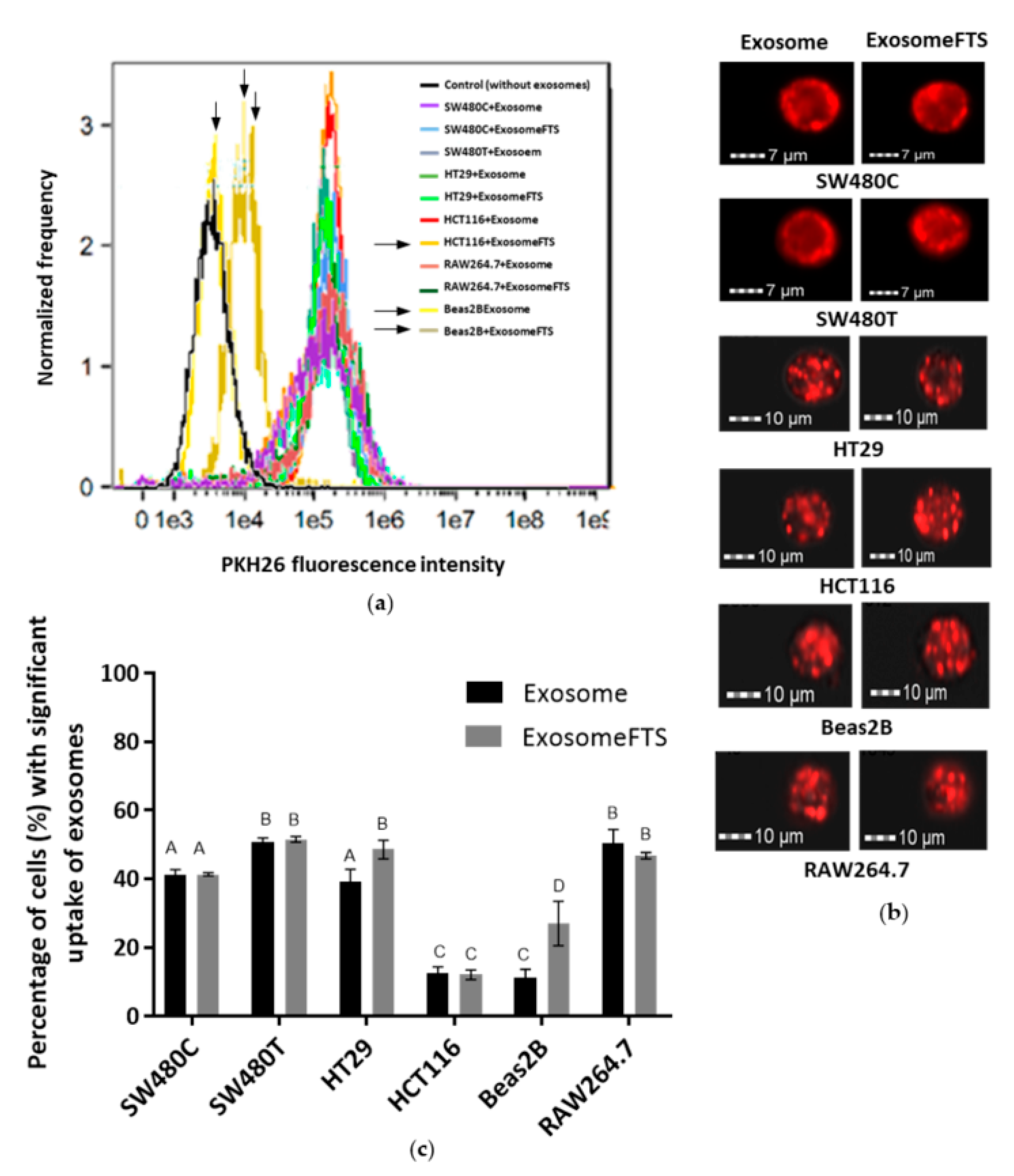
3.5.1. Exosome Uptake
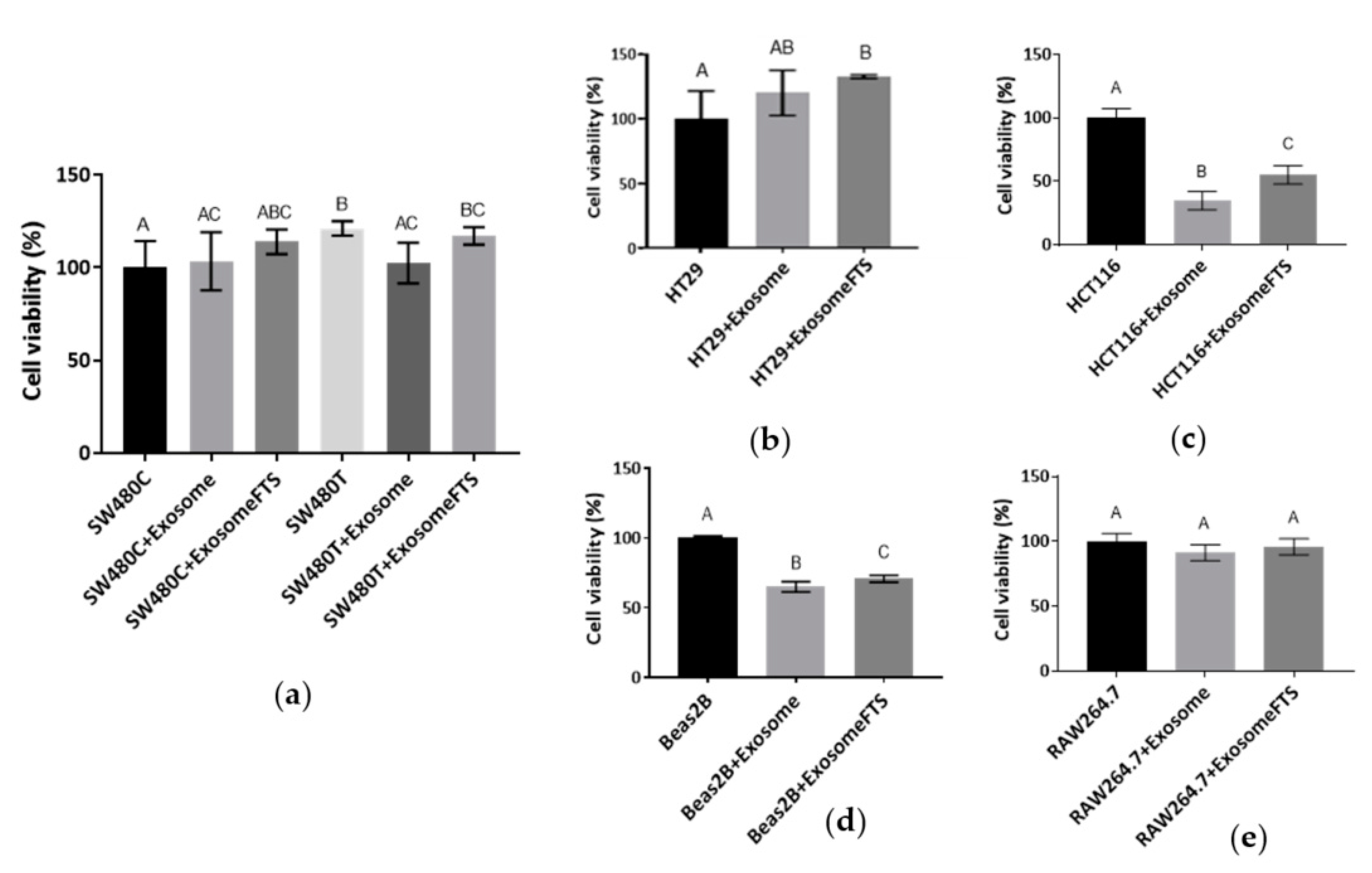
3.5.2. Viability of Recipient Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rajalingam, K.; Schreck, R.; Rapp, U.R.; Albert, Š. Ras oncogenes and their downstream targets. BBA Mol. Cell Res. 2007, 1773, 1177–1195. [Google Scholar] [CrossRef]
- Prior, I.A.; Lewis, P.D.; Mattos, C. A comprehensive survey of Ras mutations in cancer. Cancer Res. 2012, 72, 2457–2467. [Google Scholar] [CrossRef] [PubMed]
- Baines, A.T.; Xu, D.; Der, C.J. Inhibition of Ras for cancer treatment: The search continues. Future Med. Chem. 2011, 3, 1787–1808. [Google Scholar] [CrossRef] [PubMed]
- Janes, M.R.; Zhang, J.; Li, L.S.; Hansen, R.; Peters, U.; Guo, X.; Chen, Y.; Babbar, A.; Firdaus, S.J.; Darjania, L.; et al. Targeting KRAS mutant cancers with a covalent G12C-specific inhibitor. Cell 2018, 172, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Makrodouli, E.; Oikonomou, E.; Koc, M.; Andera, L.; Sasazuki, T.; Shirasawa, S.; Pintzas, A. BRAF and RAS Oncogenes regulate rho GTPase pathways to mediate migration and invasion properties in human colon cancer cells: A comparative study. Mol. Cancer 2011, 10, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Simons, M.; Raposo, G. Exosomes—Vesicular carriers for intercellular communication. Curr. Opin. Cell Biol. 2009, 21, 575–581. [Google Scholar] [CrossRef] [PubMed]
- McAndrews, K.M.; Kalluri, R. Mechanisms associated with biogenesis of exosomes in cancer. Mol. Cancer 2019, 18, 52. [Google Scholar] [CrossRef]
- Lafitte, M.; Lecointre, C.; Roche, S. Roles of exosomes in metastatic colorectal cancer. Am. J. Physiol. Cell Physiol. 2019, 317, C869–C880. [Google Scholar] [CrossRef]
- Ren, R.; Sun, H.; Ma, C.; Liu, J.; Wang, H. Colon cancer cells secrete exosomes to promote self-proliferation by shortening mitosis duration and activation of STAT3 in a hypoxic environment. Cell Biosci. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Beckler, M.D.; Higginbotham, J.N.; Franklin, J.L.; Ham, A.J.; Halvey, P.J.; Imasuen, I.E.; Whitwell, C.; Li, M.; Liebler, D.C.; Coffey, R.J. Proteomic analysis of exosomes from mutant KRAS colon cancer cells identifies intercellular transfer of mutant KRAS. Mol. Cell. Proteom. 2013, 12, 343–355. [Google Scholar] [CrossRef]
- Haklai, R.; Weisz, M.G.; Elad, G.; Paz, A.; Marciano, D.; Egozi, Y.; Ben-Baruch, G.; Kloog, Y. Dislodgment and accelerated degradation of Ras. Biochemistry 1998, 37, 1306–1314. [Google Scholar] [CrossRef] [PubMed]
- Marom, M.; Haklai, R.; Ben-Baruch, G.; Marciano, D.; Egozi, Y.; Kloog, Y. Selective inhibition of Ras-dependent cell growth by farnesylthiosalisylic acid. J. Biol. Chem. 1995, 270, 22263–22270. [Google Scholar] [CrossRef] [PubMed]
- Khoo, X.H.; Paterson, I.C.; Goh, B.H.; Lee, W.L. Cisplatin-resistance in oral squamous cell carcinoma: Regulation by tumor cell-derived extracellular vesicles. Cancers 2019, 11, 1166. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2016, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Kucharzewska, P.; Belting, M. Emerging roles of extracellular vesicles in the adaptive response of tumour cells to microenvironmental stress. J. Extracell Vesicles 2013, 2, 20304. [Google Scholar] [CrossRef]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Thery, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef]
- Parolini, I.; Federici, C.; Raggi, C.; Lugini, L.; Palleschi, S.; de Milito, A.; Coscia, C.; Iessi, E.; Logozzi, M.; Molinari, A.; et al. Microenvironmental pH is a key factor for exosome traffic in tumor cells. J. Biol. Chem. 2009, 284, 34211–34222. [Google Scholar] [CrossRef]
- Lee, S.J.; Jung, Y.S.; Lee, S.H.; Chung, H.Y.; Park, B.J. Isolation of a chemical inhibitor against K-Ras-induced p53 suppression through natural compound screening. Int. J. Oncol. 2009, 34, 1637–1643. [Google Scholar]
- Brady, C.A.; Attardi, L.D. p53 at a glance. J. Cell Sci. 2010, 123, 2527–2532. [Google Scholar] [CrossRef]
- Jin, A.; Itahana, K.; O’Keefe, K.; Zhang, Y. Inhibition of HDM2 and activation of p53 by ribosomal protein L23. Mol. Cell. Biol. 2004, 24, 7669–7680. [Google Scholar] [CrossRef]
- Küch, E.M.; Vellaramkalayil, R.; Zhang, I.; Lehnen, D.; Brügger, B.; Stremmel, W.; Ehehalt, R.; Poppelreuther, M.; Füllekrug, J. Differentially localized acyl-CoA synthetase 4 isoenzymes mediate the metabolic channeling of fatty acids towards phosphatidylinositol. BBA Mol. Cell Biol. Lipids 2014, 1841, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Wang, Y.; Sun, B.; Xiao, Z.; Ye, L.; Zhang, X. MiR-205 modulates abnormal lipid metabolism of hepatoma cells via targeting acyl-CoA synthetase long-chain family member 1 (ACSL1) mRNA. Biochem. Biophys. Res. Commun. 2014, 444, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Martínez, R.; Cruz-Gil, S.; García-Álvarez, M.S.; Reglero, G.; de Molina, A.R. Complementary ACSL isoforms contribute to a non-Warburg advantageous energetic status characterizing invasive colon cancer cells. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cai, X.; Zhang, S.; Cui, M.; Liu, F.; Sun, B.; Zhang, W.; Zhang, X.; Ye, L. HBXIP up-regulates ACSL1 through activating transcriptional factor Sp1 in breast cancer. Biochem. Biophys. Res. Commun. 2017, 484, 565–571. [Google Scholar] [CrossRef]
- Padanad, M.S.; Konstantinidou, G.; Venkateswaran, N.; Melegari, M.; Rindhe, S.; Mitsche, M.; Yang, C.; Batten, K.; Huffman, K.E.; Liu, J.; et al. Fatty acid oxidation mediated by Acyl-CoA synthetase long chain 3 is required for mutant KRAS lung tumorigenesis. Cell Rep. 2016, 16, 1614–1628. [Google Scholar] [CrossRef]
- Eldh, M.; Ekström, K.; Valadi, H.; Sjöstrand, M.; Olsson, B.; Jernås, M.; Lötvall, J. Exosomes communicate protective messages during oxidative stress; possible role of exosomal shuttle RNA. PloS ONE 2010, 5, e15353. [Google Scholar] [CrossRef]
- Yaari-Stark, S.; Shaked, M.; Nevo-Caspi, Y.; Jacob-Hircsh, J.; Shamir, R.; Rechavi, G.; Kloog, Y. Ras inhibits endoplasmic reticulum stress in human cancer cells with amplified Myc. Int. J. Cancer 2010, 126, 2268–2281. [Google Scholar]
- Montecalvo, A.; Larregina, A.T.; Shufesky, W.J.; Beer Stolz, D.; Sullivan, M.L.; Karlsson, J.M.; Baty, C.J.; Gibson, G.A.; Erdos, G.; Wang, Z.; et al. Mechanism of transfer of functional microRNAs between mouse dendritic cells via exosomes. Blood 2012, 119, 756–766. [Google Scholar] [CrossRef]
- McKelvey, K.J.; Powell, K.L.; Ashton, A.W.; Morris, J.M.; McCracken, S.A. Exosomes: Mechanisms of uptake. J. Circ. Biomark. 2015, 4, 7. [Google Scholar] [CrossRef]
- Feng, D.; Zhao, W.L.; Ye, Y.Y.; Bai, X.C.; Liu, R.Q.; Chang, L.F.; Zhou, Q.; Sui, S.F. Cellular internalization of exosomes occurs through phagocytosis. Traffic 2010, 11, 675–687. [Google Scholar] [CrossRef]
- Horibe, S.; Tanahashi, T.; Kawauchi, S.; Murakami, Y.; Rikitake, Y. Mechanism of recipient cell-dependent differences in exosome uptake. BMC Cancer 2018, 18, 47. [Google Scholar] [CrossRef] [PubMed]
- Laheru, D.; Shah, P.; Rajeshkumar, N.V.; McAllister, F.; Taylor, G.; Goldsweig, H.; Le, D.T.; Donehower, R.; Jimeno, A.; Linden, S.; et al. Integrated preclinical and clinical development of S-trans, trans-Farnesylthiosalicylic Acid (FTS, Salirasib) in pancreatic cancer. Investig. New Drugs 2012, 30, 2391–2399. [Google Scholar] [CrossRef] [PubMed]
- Furuse, J.; Kurata, T.; Okano, N.; Fujisaka, Y.; Naruge, D.; Shimizu, T.; Kitamura, H.; Iwasa, T.; Nagashima, F.; Nakagawa, K. An early clinical trial of Salirasib, an oral RAS inhibitor, in Japanese patients with relapsed/refractory solid tumors. Cancer Chemother. Pharmacol. 2018, 82, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Schmukler, E.; Grinboim, E.; Schokoroy, S.; Amir, A.; Wolfson, E.; Kloog, Y.; Pinkas-Kramarski, R. Ras inhibition enhances autophagy, which partially protects cells from death. Oncotarget 2013, 4, 142. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cellular Components (GO) | |||
|---|---|---|---|
| Pathway ID | Pathway Description | Count in Gene Set | False Discovery Rate |
| GO:0070062 | Extracellular exosomes | 319 | 1.51 × 10−189 |
| GO:0044421 | Extracellular region part | 324 | 4.36 × 10−158 |
| GO:0031988 | Membrane-bound vesicle | 316 | 1.81 × 10−157 |
| GO:0031982 | Vesicle | 314 | 4.01 × 10−151 |
| GO:0005576 | Extracellular region | 327 | 2.3e1 × 10−138 |
| Molecular Functions (GO) | |||
|---|---|---|---|
| Pathway ID | Pathway Description | Count in Gene Set | False Discovery Rate |
| GO:0005515 | Protein binding | 236 | 3.52 × 10−51 |
| GO:0005488 | Binding | 317 | 3.14 × 10−28 |
| GO:0097159 | Organic cyclic compound binding | 213 | 4.42 × 10−25 |
| GO:1901363 | Heterocyclic compound binding | 211 | 5.77 × 10−25 |
| GO:0003674 | Molecular function | 299 | 7.49 × 10−7 |
| Biological Processes (GO) | |||
|---|---|---|---|
| Pathway ID | Pathway Description | Count in Gene Set | False Discovery Rate |
| GO:0051234 | Establishment of localization | 176 | 2.52 × 10−34 |
| GO:0006810 | Transport | 171 | 2.94 × 10−33 |
| GO:0051179 | Localization | 193 | 9.41 × 10−33 |
| GO:0006950 | Response to stress | 163 | 6.69 × 10−29 |
| GO:0016043 | Cellular component organization | 185 | 1.18 × 10−23 |
| Gene ID | Protein Name | Fold Change |
|---|---|---|
| Proteins up-regulated in FTS-treated cells | ||
| AASDH | Acyl-CoA synthetase family member 4 (ACSL4) | 34 |
| RPS5 | 40S ribosomal protein S5;40S ribosomal protein S5, N-terminally processed | 23 |
| SYNCRIP | Heterogeneous nuclear ribonucleoprotein Q | 11 |
| RPL10A | 60S ribosomal protein L10a | 6 |
| RPL18A | 60S ribosomal protein L18a | 6 |
| AHNAK | Neuroblast differentiation-associated protein AHNAK | 6 |
| ANXA1 | Annexin A1 | 4 |
| ALDH16A1 | Aldehyde dehydrogenase family 16 member A1 | 3 |
| TGM2 | Protein-glutamine gamma-glutamyltransferase 2 | 2 |
| Proteins down-regulated in FTS-treated cells | ||
| PTGFRN | Prostaglandin F2 receptor negative regulator | 2 |
| ARRDC1 | Arrestin domain-containing protein 1 | 2 |
| KIF5B | Kinesin-1 heavy chain | 3 |
| STIP1 | Stress-induced-phosphoprotein 1 | 3 |
| MPZL2 | Myelin protein zero-like protein 2 | 4 |
| DDX39A; DDX39B | ATP-dependent RNA helicase DDX39A; Spliceosome RNA helicase DDX39B | 5 |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eng, S.-K.; Imtiaz, I.R.; Goh, B.-H.; Ming, L.C.; Lim, Y.-C.; Lee, W.-L. Does KRAS Play a Role in the Regulation of Colon Cancer Cells-Derived Exosomes? Biology 2021, 10, 58. https://doi.org/10.3390/biology10010058
Eng S-K, Imtiaz IR, Goh B-H, Ming LC, Lim Y-C, Lee W-L. Does KRAS Play a Role in the Regulation of Colon Cancer Cells-Derived Exosomes? Biology. 2021; 10(1):58. https://doi.org/10.3390/biology10010058
Chicago/Turabian StyleEng, Shu-Kee, Ilma Ruzni Imtiaz, Bey-Hing Goh, Long Chiau Ming, Ya-Chee Lim, and Wai-Leng Lee. 2021. "Does KRAS Play a Role in the Regulation of Colon Cancer Cells-Derived Exosomes?" Biology 10, no. 1: 58. https://doi.org/10.3390/biology10010058
APA StyleEng, S.-K., Imtiaz, I. R., Goh, B.-H., Ming, L. C., Lim, Y.-C., & Lee, W.-L. (2021). Does KRAS Play a Role in the Regulation of Colon Cancer Cells-Derived Exosomes? Biology, 10(1), 58. https://doi.org/10.3390/biology10010058