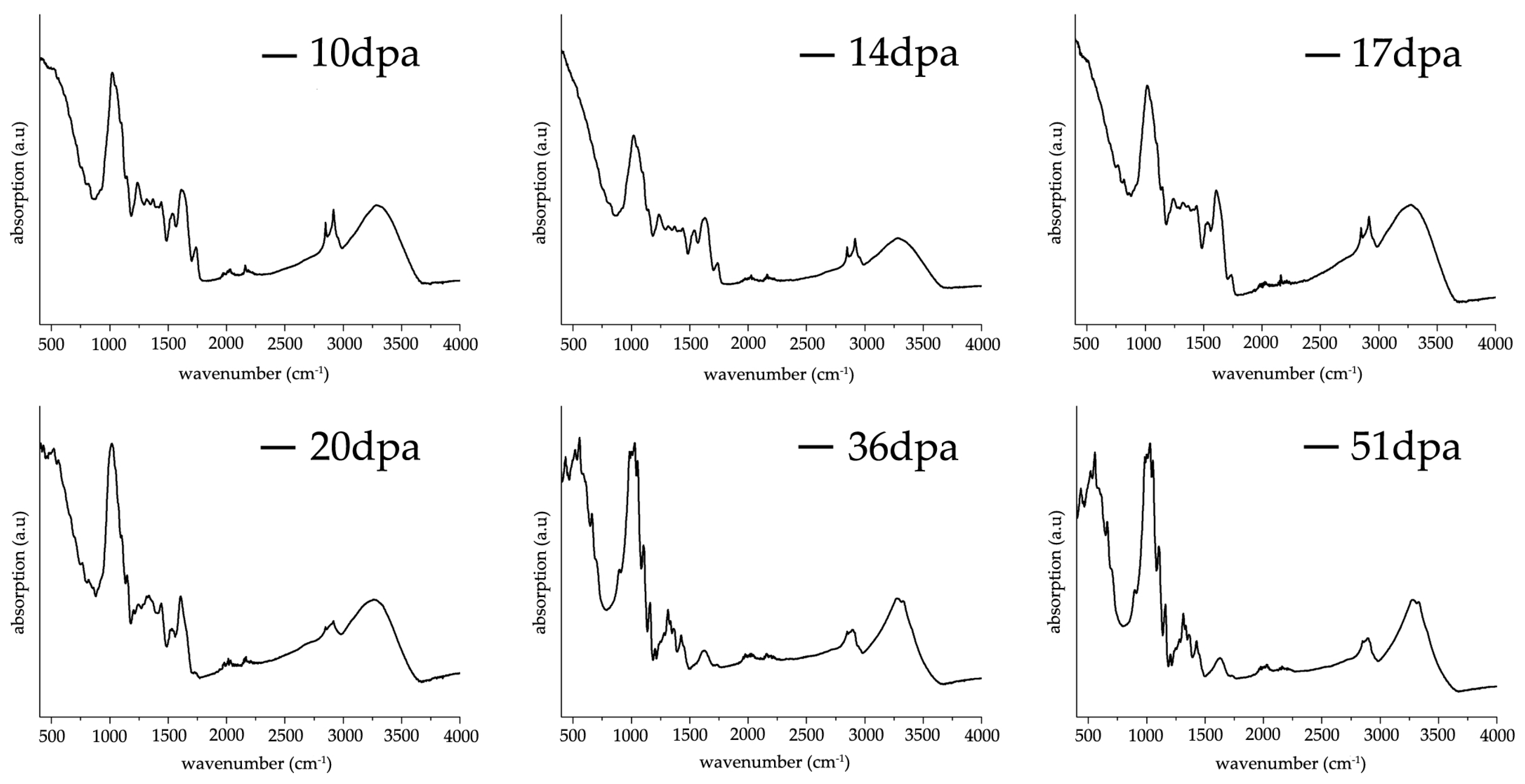
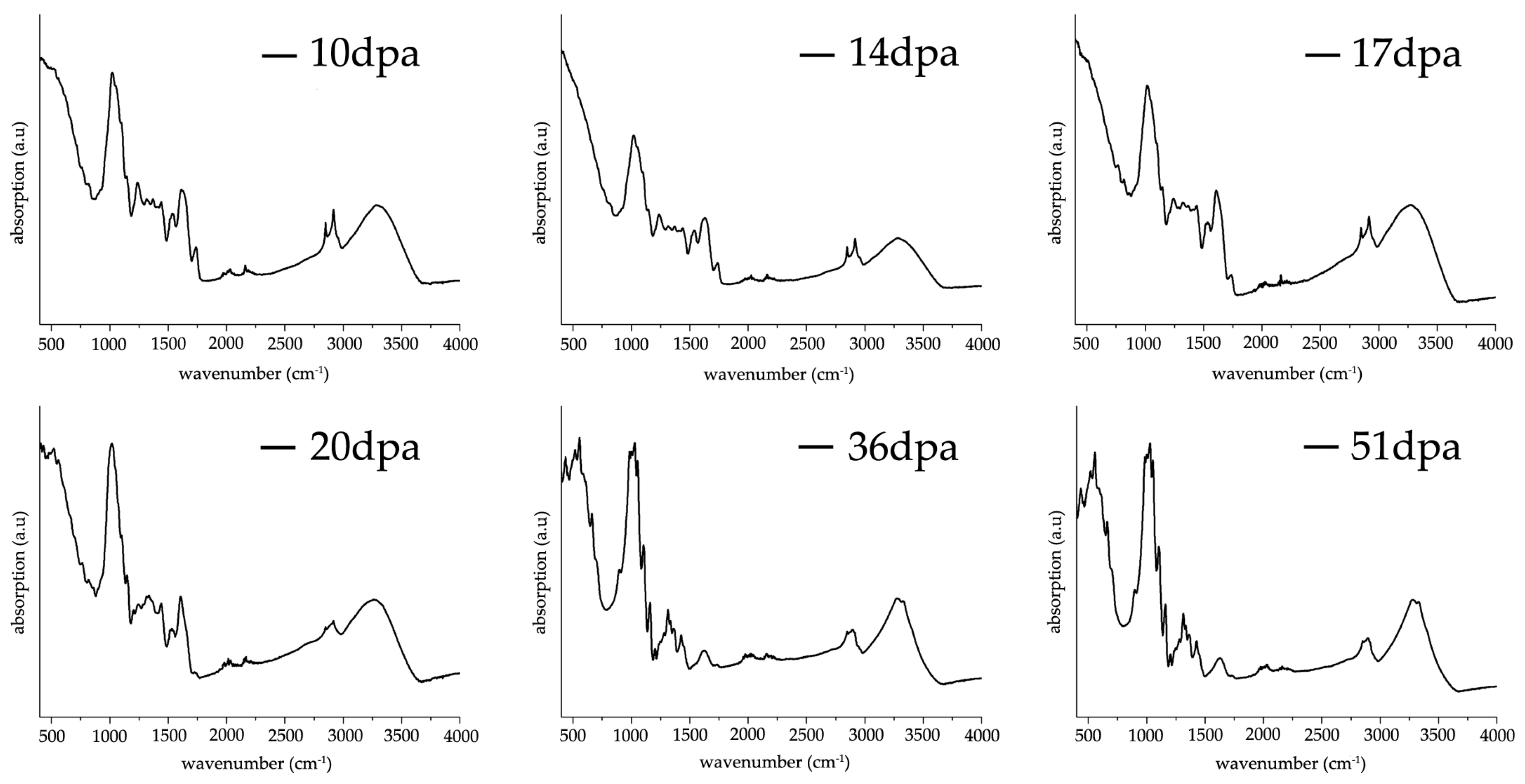
3.3. FT-IR ATR Characterization of G. hirsutum Fibers Development between 10–51 dpa
Figure 3 shows a sequence of representative FT-IR ATR spectra taken from washed and air-dried fibers harvested at different stages of development (10, 14, 17, 20, 36 and 51 dpa). The spectral regions between 3000–3600 cm
−1 corresponds to stretching vibrations of OH groups derived from both water molecules and intra/inter hydrogen bonding in cellulose [
7]. From 10 dpa until 20 dpa, one single broad band is observed whereas at 36 and 51 dpa a double band appears, displaying higher intensity at 51 dpa. Interestingly, the broad maximum band found between 10 dpa to 20 dpa shifts towards lower wavenumbers, i.e., 3297/3281/3277/3263 cm
−1 for 10/14/17/20 dpa respectively. Contrary, the maximum band height for the double bands (3277 and 3329 cm
−1) found at 36 and 51 dpa remains inalterable in terms of shift and intensity, suggesting a higher level of structural organization. At the current stage, it is not possible to attribute the double band and its significance to cotton fiber structural organization solely on the assumption of an increase of intra- and intermolecular hydrogen bonding [
14,
15] taking into consideration the contribution of water molecules OH stretching vibrations, their effect on the structural organization of the cellulose chains (hydrogen bonds) and in addition the contribution of homogalacturonans from pectins (OH stretching vibrations) that disappear during the second phase of cell wall synthesis.
Double sharp bands found at 2848 and 2916 cm−1 attributed to CH2 symmetric stretching are present until 17 dpa, where no band shift is observed. After 17 dpa, both bands intensities decrease to form an undistinguished broad band. Both broadening and band intensity decrease infers a transition from an elongation (primary cell wall synthesis) to a maturation (secondary wall synthesis) process as the ratio between cellulose and non-cellulosic components (e.g., pectins) become higher.
The band located at 1738 cm
−1 decreases in intensity until 17 dpa without varying its maximum wavenumber. After 17 dpa, this band remains extremely broad and with almost negligible intensity, following a similar trend as found for the bands located at 2848 and 2916 cm
−1 corresponding to CH
2 symmetric stretching. This band is attributed to C=O stretching of saturated alkyl esters associated to pectins [
16], and/or waxes [
17] and it is strongly influenced by hydrogen bonds [
18]. When 51 dpa
G. hirsutum fibers grown under the same conditions are treated with NaOH/ ethylenediaminetetraacetic acid/sodium dodecyl sulfate (EDTA/SDS) for 1 h at 80 °C (scouring), this band completely disappears [
7] supporting the hypothesis of a hydrophobic nature of the components (waxes or alkylated pectins). Consequently, these findings support the idea that this band can serve to quickly demonstrate the transition between the different stages of cell wall synthesis in
G. hirsutum.
In the spectral region between 1611 and 1628 cm
−1 a double broad band at 10 dpa is observed. At 14 dpa, it becomes one single broad band centered at 1630 cm
−1 displaying a decreased intensity. Interestingly, between 17 and 20 dpa, this single band sharpens and shifts towards lower wavenumbers (1606 cm
−1) but retains its intensity at 17 dpa with a slight increase at 20 dpa. Typically, the vibrations found in this range are associated with the asymmetric stretching of carboxylic acids in the ionic form [
18]. At 36 dpa, the band broadens, decreases in intensity and shifts to higher wavenumbers (1623 cm
−1). Finally, at 51 dpa an increase in band intensity and shifting to 1630 cm
−1 is observed in comparison with the 36 dpa band. This wavenumber fluctuation is in opposition to previous findings, where one single band remains at a fixed wavenumber (1633 cm
−1) through all developmental stages of
G. hirsutum L. (soil-based culture) and in which band intensity decrease correlates with the dpa increasing [
14,
15]. However, this wavenumber variability throughout the different developmental stages, particularly when considering the period between 17 and 20 dpa (1606 cm
−1), which represents the metabolic transition between primary and secondary cell wall synthesis, makes the attribution to vibrations corresponding to water molecules (1620 cm
−1) and/or amide I (ca. 1620 cm
−1) difficult. In fact, this oscillatory process infers a more complex underlying process during this transitory period and provides additional evidence that hydroponic culture impacts the development of
G. hirsutum when compared with normal soil-based cultures [
14,
15].
In the region between 1491 and 1568 cm−1, a broad band centered at 1535 cm−1 with a shoulder at 1518 cm−1 is found at 10 dpa. A band with similar shape but with less intensity is observed at 14 dpa. At 17 and 20 dpa, the bands becomes broader due to the fact that the shoulder observed at 10 and 14 dpa is merged following a similar trend of that found for the bands located at 1606 cm−1, i.e., the 17 dpa band displays a lower intensity than the 20 dpa. At 36 and 51 dpa, no bands in this region are present. The interpretation of this band is straightforward when assuming that the primary and secondary cell wall synthesis transition occurs between 17 and 20 dpa. This vibration is attributed to the NH2 deformation of amide II, in particular, is a combination of N-H deformation and C-N stretching. Due to fiber thickening the amide II signal becomes obscured and, consequently, it is no longer possible to detect this vibration after the maturation process has evolved (e.g., cellulose shielding).
The vibration found at 1440 cm
−1 is primarily attributed to CH
2 vibrations (scissoring) of non-cellulosic components. It remains with a fixed maximum wavenumber (1440 cm
−1) with a linear intensity decrease until 36 dpa, where the band shifts towards lower wavenumbers (1427 cm
−1) with a progressive increase in intensity, attributed to CH
2 symmetric deformation and bending [
19], similar to soil-based cultures [
14,
20].
In addition, the presence of this band at 1427 cm
−1 has been attributed to an increase in the structural organization of cellulose (e.g., crystalline domains) and used in combination with the vibration correspondent to [β(1-4)glucopyranose] linkage to determine the “crystalline index”, later referred to as the “later order index” (LOI) [
2,
20,
21]. A similar trend is found for the double band located at 1313 and 1332 cm
−1. Between 10 and 20 dpa, the intensity of this single wide band centered at 1315 cm
−1 (CH
2 rocking) [
19] decreases, whereas after 20 dpa a double band appears with progressively increasing intensity. There seems to be a direct relation between these two vibrations. Apart from the respective CH
2 rocking vibration attributed to this band, there is, however, no significant wavenumber shift (the observed 2 cm
−1 difference is attributed instrumental error) and no relation to the crystallinity index and/or quantification [
15,
22]. Interestingly, an intense and broad band found at 1236 cm
−1 in 10 dpa fibers decreases in intensity until 17 dpa, where a neighboring sharp band located at 1203 cm
−1 rises until 51 dpa. While the existence of a band at 1245 cm
−1 has been reported to be present in soil-based cultures and assigned to C=O stretching or NH
2-deformation, it is difficult to assign the same type of vibration to the band located at 1236 cm
−1 despite the fact that it follows the same trend as found at spectral region between 1611 and 1628 cm
−1. On the other side, the band at 1203 cm
−1 can be, eventually, attributed to C-O in-plane bending or stretching of [β(1-4)glucopyranose] rings and correlated with fiber maturation/development. However this vibration is not fully enough understood to draw conclusions.
At 1145, 1105 and 1052 cm
−1, a small shoulder can be identified following a similar behavior as found for soil-based cultures [
14,
15], i.e., a decrease of intensity until 17 dpa and band shift towards higher wavenumbers (more pronounced from 1145 cm
−1 at 10 dpa to 1160 cm
−1 in 51 dpa) and band sharpening from 36 dpa until 51 dpa. These vibrations are assigned to C-O-C stretching and asymmetrical stretching of non-cellulose molecules and [β(1-4)glucopyranose] rings associated with primary and secondary cell wall development.
A broad asymmetric band centered at 1020 cm−1 decreases intensity until 20 dpa. At 36 dpa and 51 dpa, this band displays three peaks at 1029, 1000 and 983 cm−1. Despite the fact that this band is extremely complex for any attempt to attribute any possible vibrations, this is where the major differences between the hydroponic/greenhouse and soil-based G. hirsutum cultivars are found. The band typically assigned to the [β(1-4)glucopyranose] linkage is located at 897 cm−1, but only from 36 dpa onwards, indicating that at 36 dpa the development stage is already in the secondary cell wall growth regime.
At 760 cm
−1 a low-intensity wider band decreases until 17 dpa and disappears afterwards. No band is found at 710 cm
−1 and thus an assignment to one of the possible cellulose allomorphs using FT-IR ATR is not possible. This is in opposition to the previous assumption that these bands are characteristic of I
β (monoclinic, 710 cm
−1) and I
α (triclinic, 750–760 cm
−1) and cannot be used for this purpose [
23,
24,
25]. Using other characterization techniques than FT-IR ATR, the fibers from cotton grown under hydroponic/greenhouse conditions have been classified as cellulose I
β (monoclinic) [
7].
In the region below 700 cm−1, four additional bands at 653, 557 (sharp), 518 (shoulder) and 435 cm−1 are found, but only for 36 dpa and 51 dpa. Due to their existence only at the later stages of fruit development, these bands might be eventually related with secondary cell wall activity. The FT-IR ATR analysis shows that, despite several marked spectroscopic differences likely attributable to the growing conditions, the transition between primary and secondary cell wall development occurs around the 17–20 dpa similarly to the soil-based cultures.
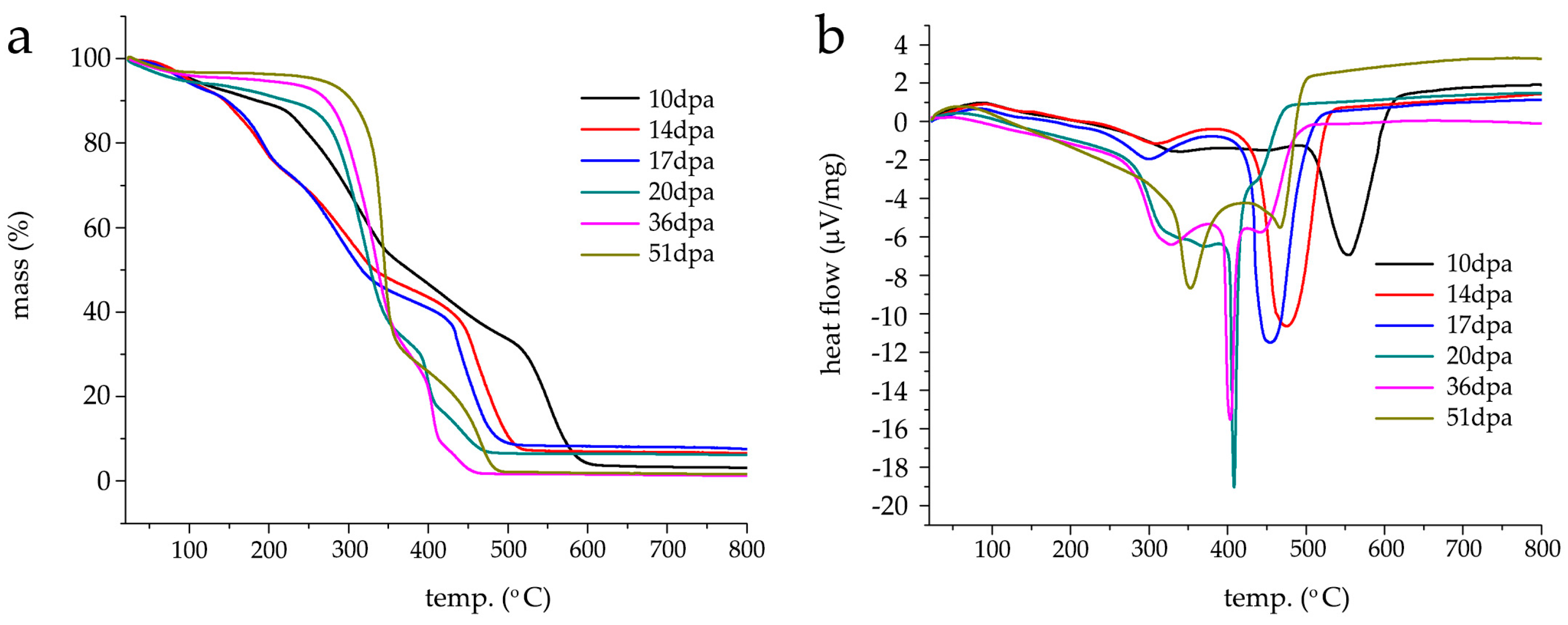
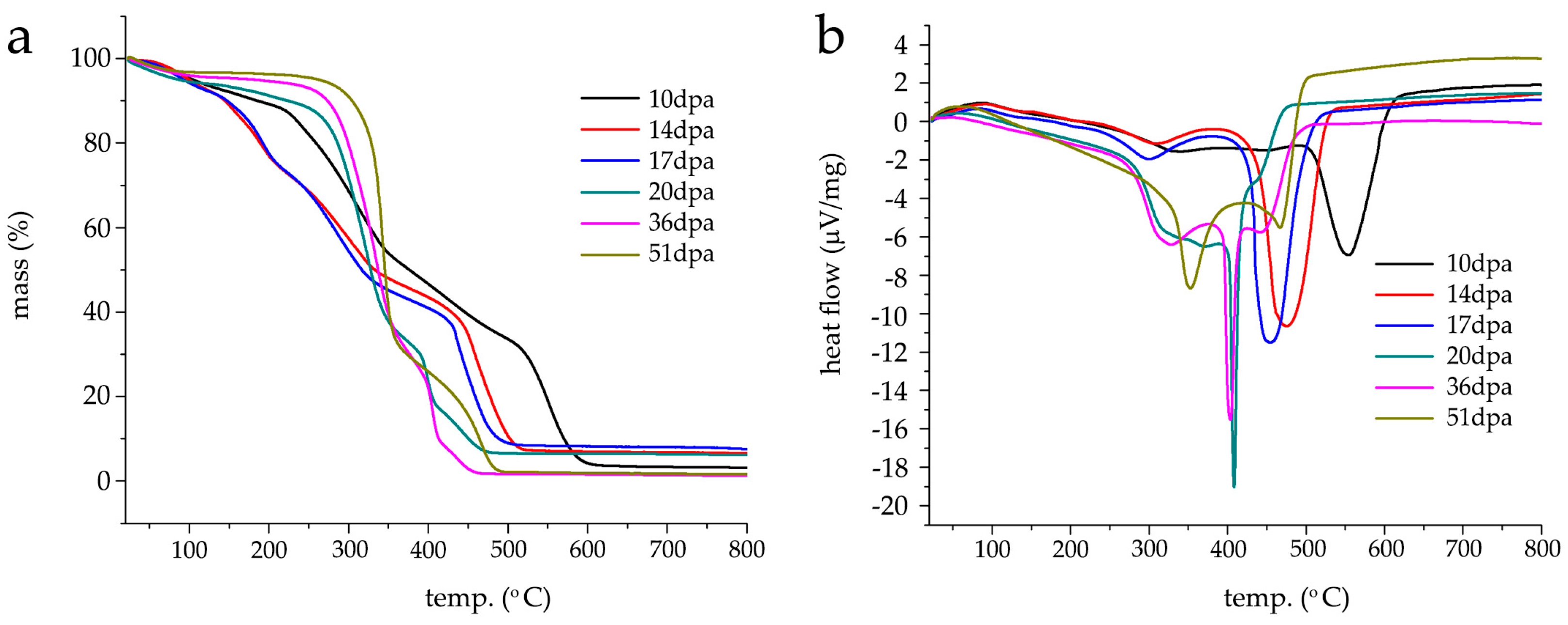
3.4. TGA/DSC Characterization of G. hirsutum Fibers Development between 10–51 dpa
Thermogravimetric (TGA) and differential scanning calorimetric (DSC) analysis were performed on washed/air-dried fibers kept at 52% RH and retrieved at different time points of fruit development, namely, 10, 14, 17, 20, 36 and 51 dpa.
Figure 4a shows an assembly of averaged thermograms obtained for each set of fibers (under air). A cotton cellulose thermogram is, normally, characterized by three weight loss regions ranging between 60–150 °C, 150–250 °C and 250–500 °C, corresponding to water loss and volatile non-cellulosic components (e.g., waxes, fatty acids, esters), and the decomposition of non-cellulosic (e.g., proteins, pectins and hemicellulose) and cellulosic components/char decomposition, respectively. As previously reported, water quantification in non-scoured raw fibers is difficult to attribute due to a masking effect by the coexistence of water molecules and primary cell wall volatile non-cellulosic components [
7,
17] and even in scoured samples it is almost virtually impossible to distinguish between freezing-bound and non-freezing water fractions.
In region I, the fibers from 10 dpa lose 1.58 ± 0.15% of their initial mass (
Figure 4a black line), with a slightly lesser mass loss at 14 dpa (1.02 ± 0.08%) (
Figure 4a red line). The percentage mass loss is higher for 17 (
Figure 4a light blue line), 20 (
Figure 4a dark green line), 36 (
Figure 4a pink line) and 51 dpa (
Figure 4a light green line) where values of 1.58 ± 0.07%, 3.55 ± 0.04%, 2.42 ± 0.14% and 2.07 ± 0.05% are respectively found. At 150 °C, 10 and 14 dpa shows a percentage mass loss of 7.86 ± 0.6% and 11.04 ± 0.45%, respectively, where 14 dpa represents a certain turning point. From this point on, the percentage mass loss values decreases as the fruit development increases. Values of 10.40 ± 0.08%, 6.62 ± 0.08%, 4.59 ± 0.06% and 3.34 ± 0.05% are found and correspond to 17, 20, 36 and 51 dpa. Interestingly, the thermogram profiles obtained from 10, 14 and 17 dpa show an inflexion point around 100 °C and 20 dpa at 120 °C, whereas the profiles of 36 and 51 dpa show a continuous logarithmic decay from room temperature until 150 °C, indicating a difference in overall composition (first derivative of TGA is found in
Figure S1). It seems that 20 dpa is the threshold between primary and secondary cell wall growth supporting the results obtained by FT-IR ATR.
In the region II (150–250 °C), 10 and 20 dpa have a unique thermographic profile and both 14/17 dpa and 36/51 dpa display similar profiles. At 10 dpa, the profile is linearly descending until 212 °C (percentage mass loss of 11.54 ± 0.06%). After this temperature, a pronounced decrease is observed, reaching a percentage mass loss of 17.86 ± 0.08% at 250 °C. For 14 and 17 dpa, inflexion points are observed at 189 and 191 °C with a corresponding percentage mass loss of 20 ± 0.08% and 19.6 ± 0.2%, respectively. From this temperature onwards, both profiles overlap until 250 °C with a corresponding percentage mass loss of 31.40 ± 0.06% and 31.73 ± 0.04% for 14 and 17 dpa. Compared to the percentage mass loss found for 10 dpa at 250 °C, these values are almost two-fold higher. The logarithmic decay-like behavior (i.e., no inflections points) for 20, 36 and 51 dpa continues throughout this temperature range with percentage mass losses of 11.67 ± 0.07%, 6.93 ± 0.07% and 4.49 ± 0.05% at 250 °C, respectively. The behavior found for 14, 17, 20, 36 and 51 dpa thermographic profiles correlates well with the attribution of this region to decomposition of non-cellulosic compounds and the primary/secondary wall switching, i.e., at 14 and 17 dpa the maximum content of non-cellulosic compounds in the fibers is found in opposition to the lower content observed for 20, 36 and 51 dpa.
In region three—attributed to the decomposition of cellulosic compounds ranging from 250–500 °C—the profiles differ more significantly between them. The 10 dpa profile shows a smooth inflexion point at 344 °C (44.86 ± 0.2% mass loss) with a linearly decreasing profile until 500 °C where a residual mass loss of 33.61 ± 0.07% assigned to charcoal formation is observed. The 14 and 17 dpa profiles are similar, i.e., they both decrease linearly until approximately 424 °C with major inflexion points appearing at 458 and 435 °C with correspondent percentage mass losses of 69.75 ± 0.45% and 69.95 ± 0.05%. At 500 °C, there is a decrease of mass loss of 89.36 ± 0.25% (14 dpa) and 90.99 ± 0.19% (17 dpa) assigned to the formation of charcoal. These values are three-fold smaller than the one found for the 10 dpa profile suggesting a significantly different chemical composition.
The 20, 36 and 51 dpa profiles show similar behavior between them, i.e., a larger percentage mass loss is clearly observed in the temperature range between 250–375 °C and a smaller percentage mass loss beyond 400 °C and three plateaus. At 20 dpa, two major inflexion points are found at 324 and 399 °C with correspondent percentage mass losses of 47.12 ± 0.25% and 75.07 ± 0.74%. At 500 °C, the mass loss is of 93.4 ± 0.5%. Interestingly, the 36 dpa profile is very similar to that of 20 dpa. However, the two major inflexion points are found at 334 °C (9 °C higher than 20 dpa) and 405 °C (6 °C higher than 20 dpa) with a correspondent percentage mass loss of 47.3 ± 0.47% and 82.96 ± 0.45%. At 500 °C, the mass loss is of 98.32 ± 0.34%. The difference in these values suggests that the fibers are in a transitory chemical composition. The 51 dpa fibers show one major inflexion point at 343 °C and one less pronounced inflexion point at 466 °C and a mass loss of 43.4 ± 0.55% and 90.9 ± 0.37%, respectively. At 500 °C, the mass loss is of 97.98 ± 1.54%. In the cases of 36 and 51 dpa, the values of residual mass are far smaller than the values found for the other fibers at different development stages leading to the conclusion that at these (later) stage fibers are composed, in the majority, by cellulosic compounds. Despite the FT-IR ATR growth profile similarity, it is in thermogravimetric profiles from hydroponic/greenhouse fibers collected at different stages that the differences with soil-based cultures are more pronounced [
14,
15,
17]. The thermogravimetric analysis shows that the fibers at different stages of development have different chemical compositions when grown under hydroponic/greenhouse conditions.
To confirm the assumption laid by thermogravimetric analysis and the FT-IR ATR dataset, the DSC analysis of the raw/washed fibers collected at different time points was also carried out, namely, 10 (
Figure 4b black line), 14 (
Figure 4b red line), 17 (
Figure 4b blue line), 20 (
Figure 4b dark green line), 36 (
Figure 4b pink line) and 51 dpa (
Figure 4b light green). Despite the different profiles, an underlying trend is observed, i.e., as fruit development advances the major exothermic event appears at lower temperatures similarly to soil-based cultures [
17]. Despite this similar trend, the profiles for the fibers grown under hydroponic/greenhouse cultures differ significantly from soil-based cultures, reinforcing the idea that hydroponic/greenhouse conditions significantly impact the cotton development [
17]. It is worth noting that the DSC analysis was carried out in washed/raw fibers. It is known that the washing step removes substances and reduces the thermogram complexity by displaying fewer thermal transitions in the lower temperature region (room temperature −150 °C). However, the differences observed for the profiles obtained by hydroponic/greenhouse-grown fibers and soil-based fibers are pronouncedly different and cannot be attributed to this “simplifying” step. The thermogram profiles for 10, 14 and 17 dpa show major symmetric endothermic bands at 553, 474 and 454 °C assigned to the crystallization temperature (
Tc), a smaller endothermic band at 339, 309 and 300 °C and an exothermic band at 86 and 84 °C, respectively. These higher temperature values overlap with the major percentage mass losses observed on the thermogravimetric analysis (
Figure 4a). Interestingly, the thermogram profile found at 20 and 36 dpa differs significantly from all the other previous profiles. It shows a very sharp symmetric band with its lowest point at 408 °C (20 dpa) and 403 °C (36 dpa) assigned to the crystallization temperature (
Tc), respectively (
Figure 4b dark green and pink). However, this sharp band found at 36 dpa is combined with two other endothermic wider and less intense bands centered at 327 °C and 441 °C. According to the thermogravimetric analysis (see
Table 1 for summary), the highest percentage mass loss found in the 36 dpa fibers coincides with the temperature corresponding to this wider and less intense endothermic band located at 327 and can be assigned to cellulose thermal decomposition [
17]. The major symmetric and sharp endothermic band is, in turn, assigned to charcoal formation. In 20 dpa and 36 dpa profiles, exothermic bands are found at 56 and 47 °C, respectively and eventually attributed to waxes [
17]. At 51 dpa, the DSC profile shows two pronounced endothermic bands (
Figure 4b light green line), one larger and symmetric band centered at 353 °C and the second smaller, asymmetric and centered at 466 °C. The band located at 353 °C corresponds to the crystallization temperature (
Tc) and assigned to cellulose thermal decomposition infers the complete absence of primary cell wall with a well-defined chemical composition. As in the previous profiles, a small exothermic band is observed at 59 °C. A summary of the thermal behavior of
Gossypium hirsutum fibers is presented in
Table 1.





{kind=link}
{kind=link}
{kind=link}
{kind=link}