
Attachment and Growth of Fibroblast Cells on Poly (2-Methoxyethyl Acrylate) Analog Polymers as Coating Materials

 , , ,
, , , 
Abstract
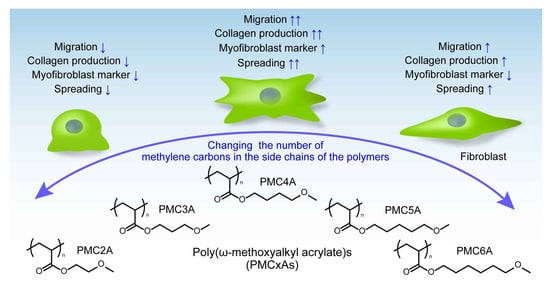
1. Introduction
2. Materials and Methods
2.1. Preparation of Polymer-Coated Substrates
2.2. Contact Angle Measurement
2.3. Cell Culture
2.4. Cell Attachment and Proliferation Assays
2.5. Quantification of Adsorbed Protein on Polymer Substrates
2.6. Immunocytochemical Analysis
2.7. Western Blotting
2.8. Cell Migration Test
2.9. Collagen Production Measurement
2.10. Statistical Analysis
3. Results and Discussion
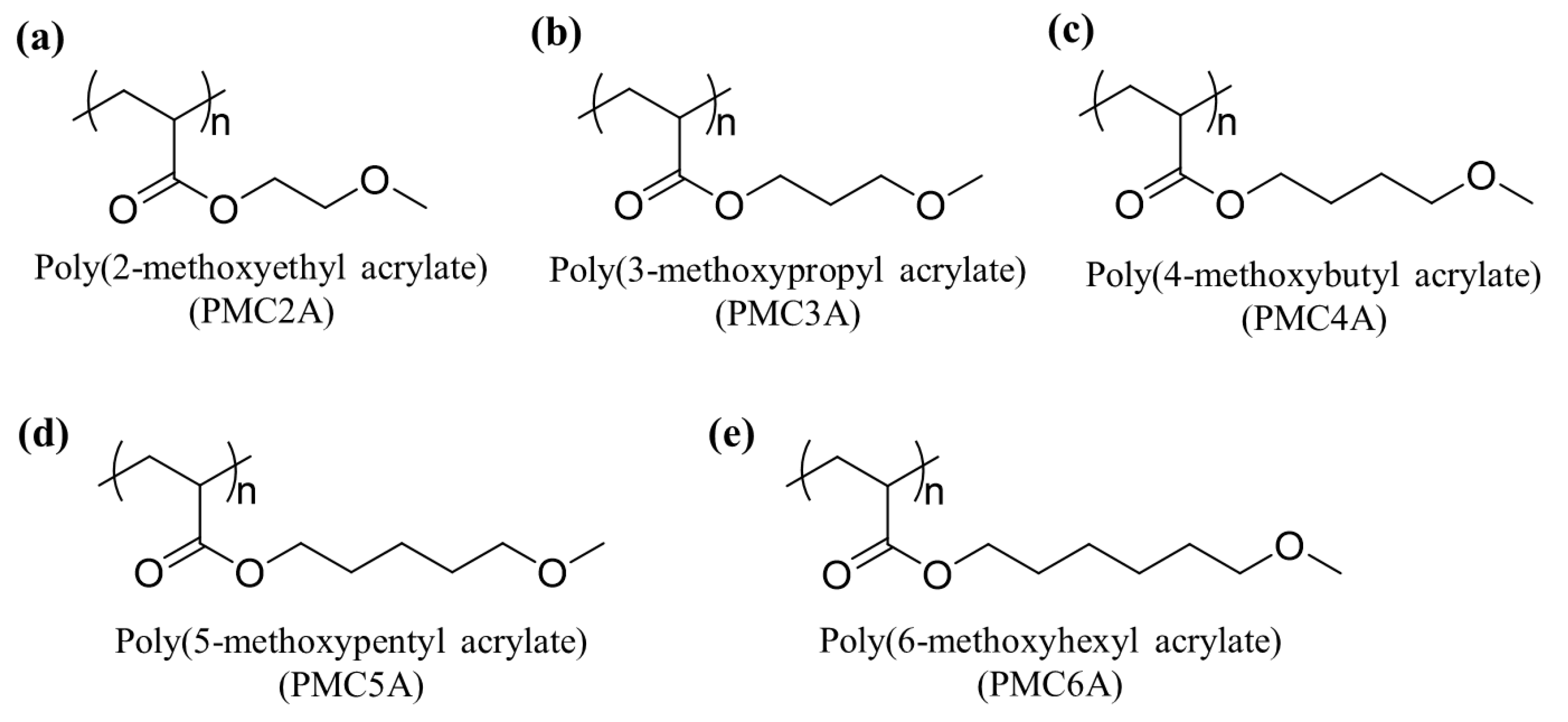
3.1. Physicochemical Properties of PMCxA-Coated Substrates
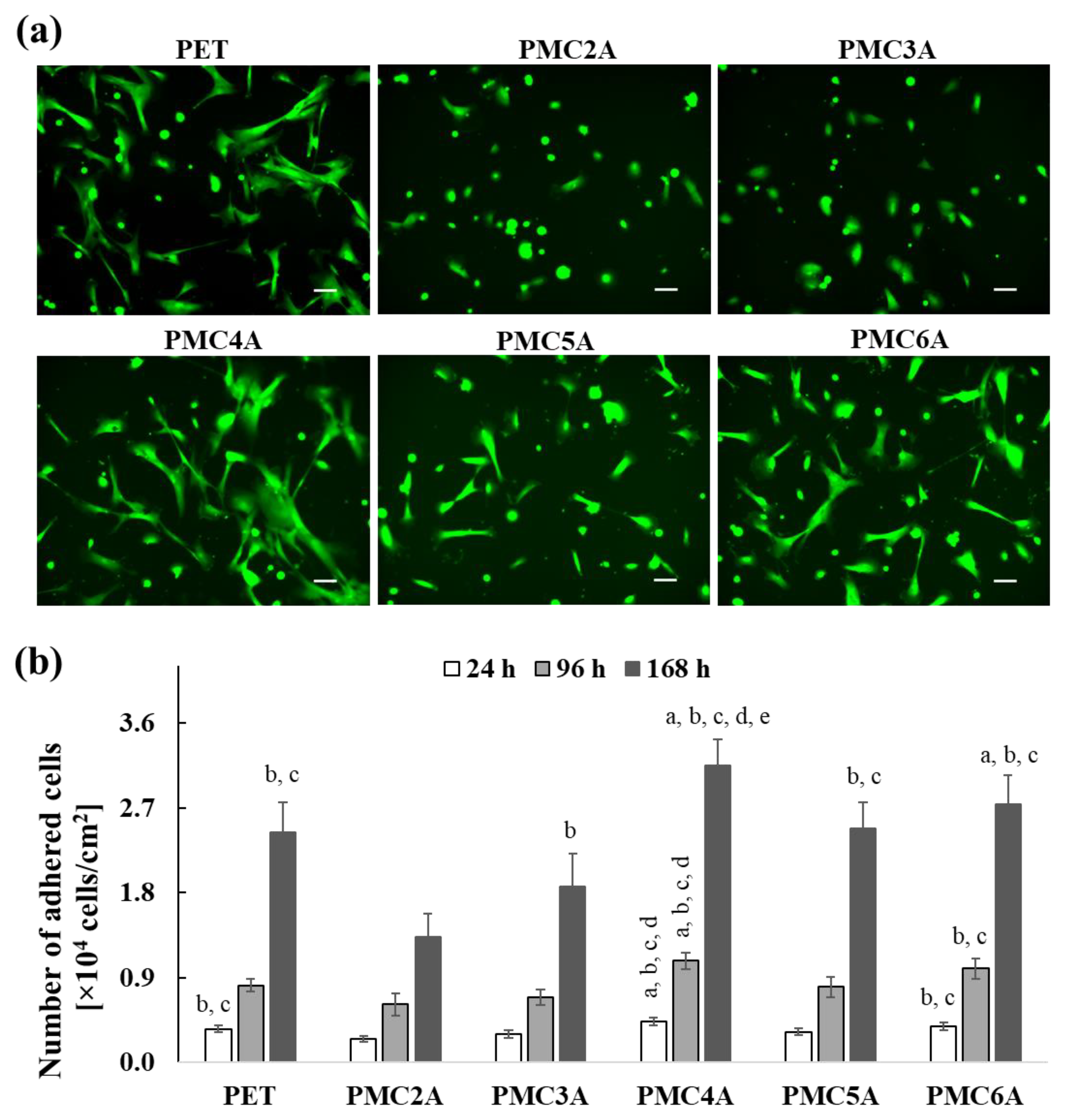
3.2. Cell Attachment and Proliferation Assay
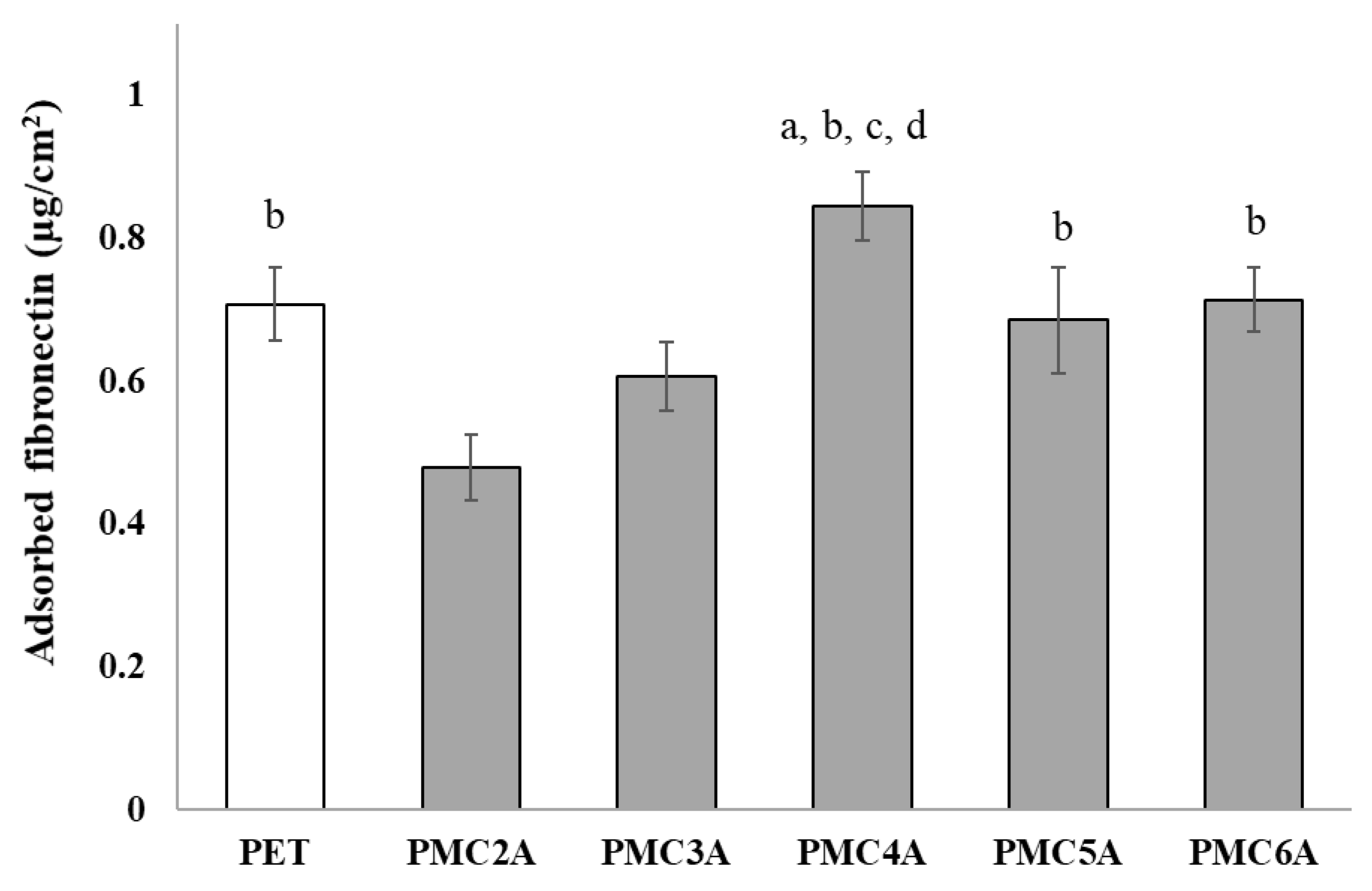
3.3. Quantification of Adsorbed Protein on Polymer Substrates
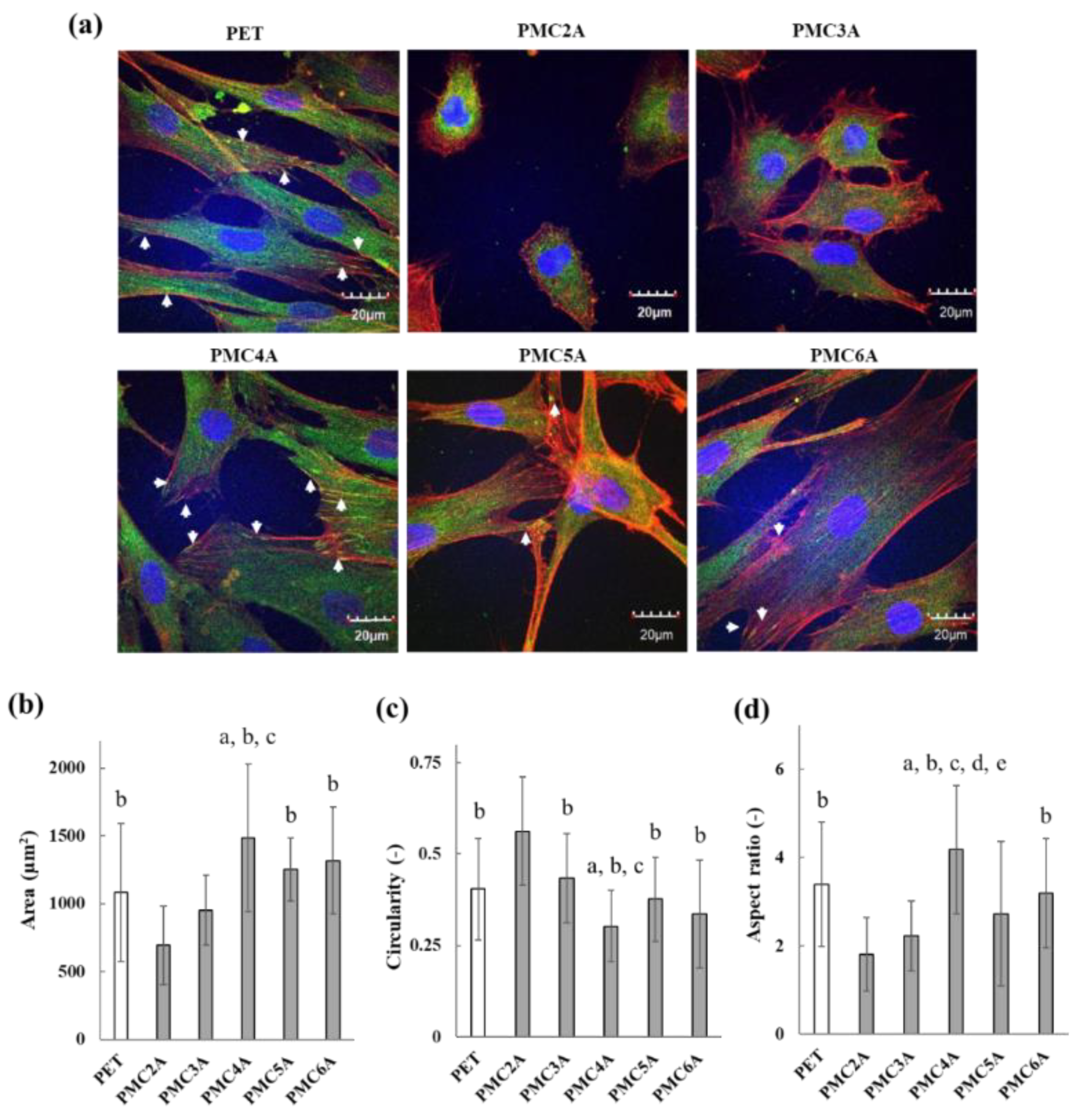
3.4. Immunocytochemical Assay
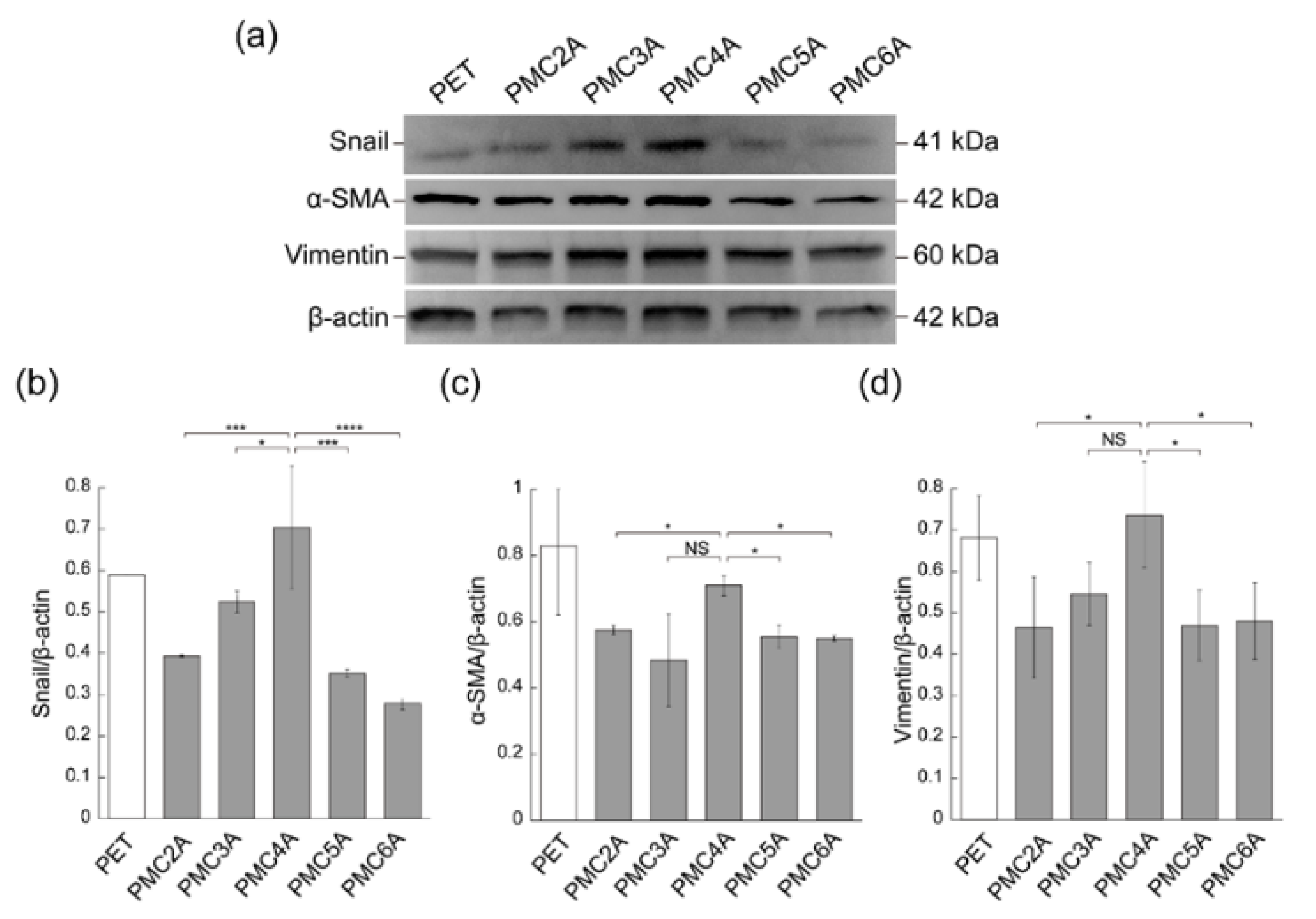
3.5. Immunoblotting Studies
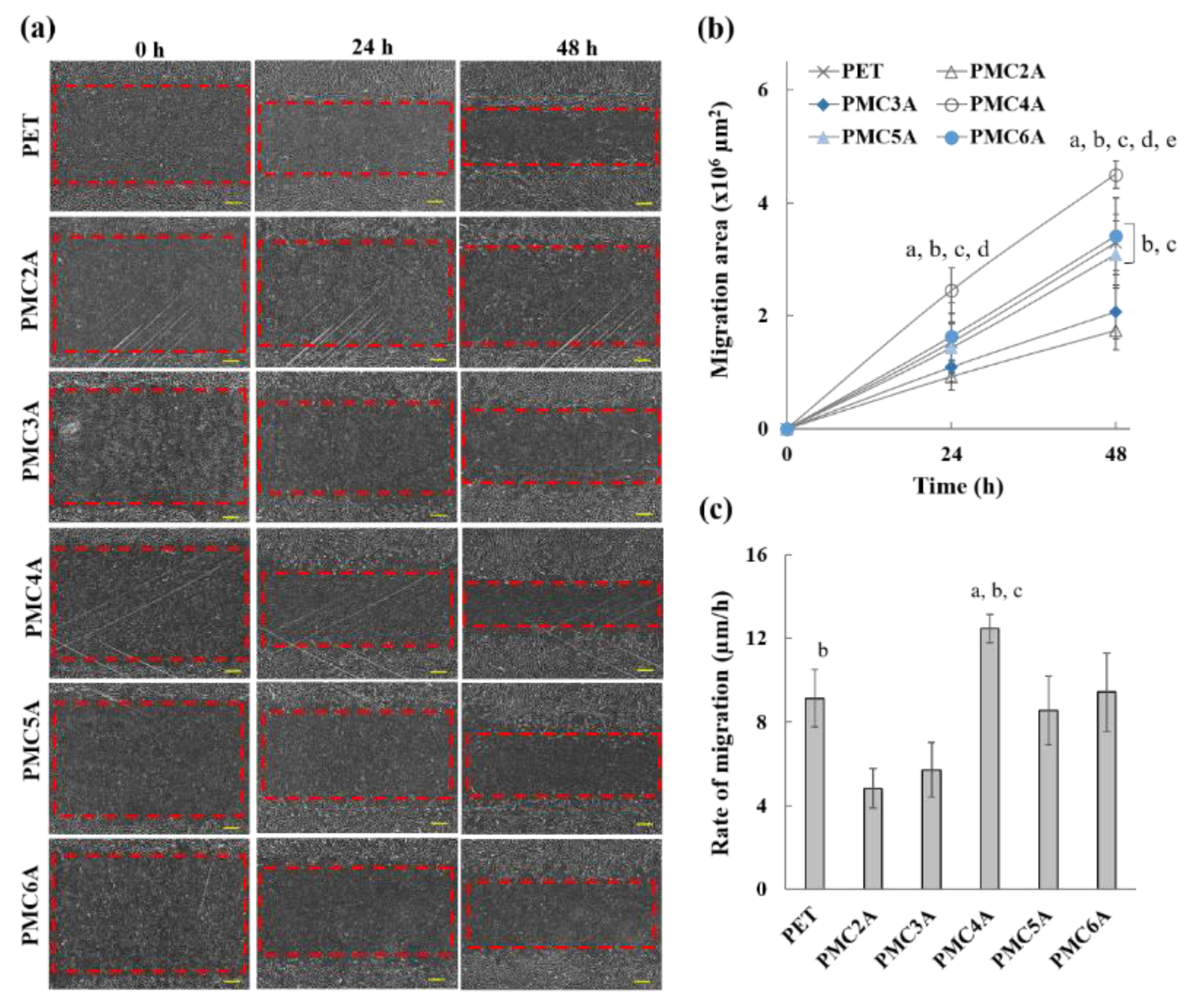
3.6. Cell Migration Test
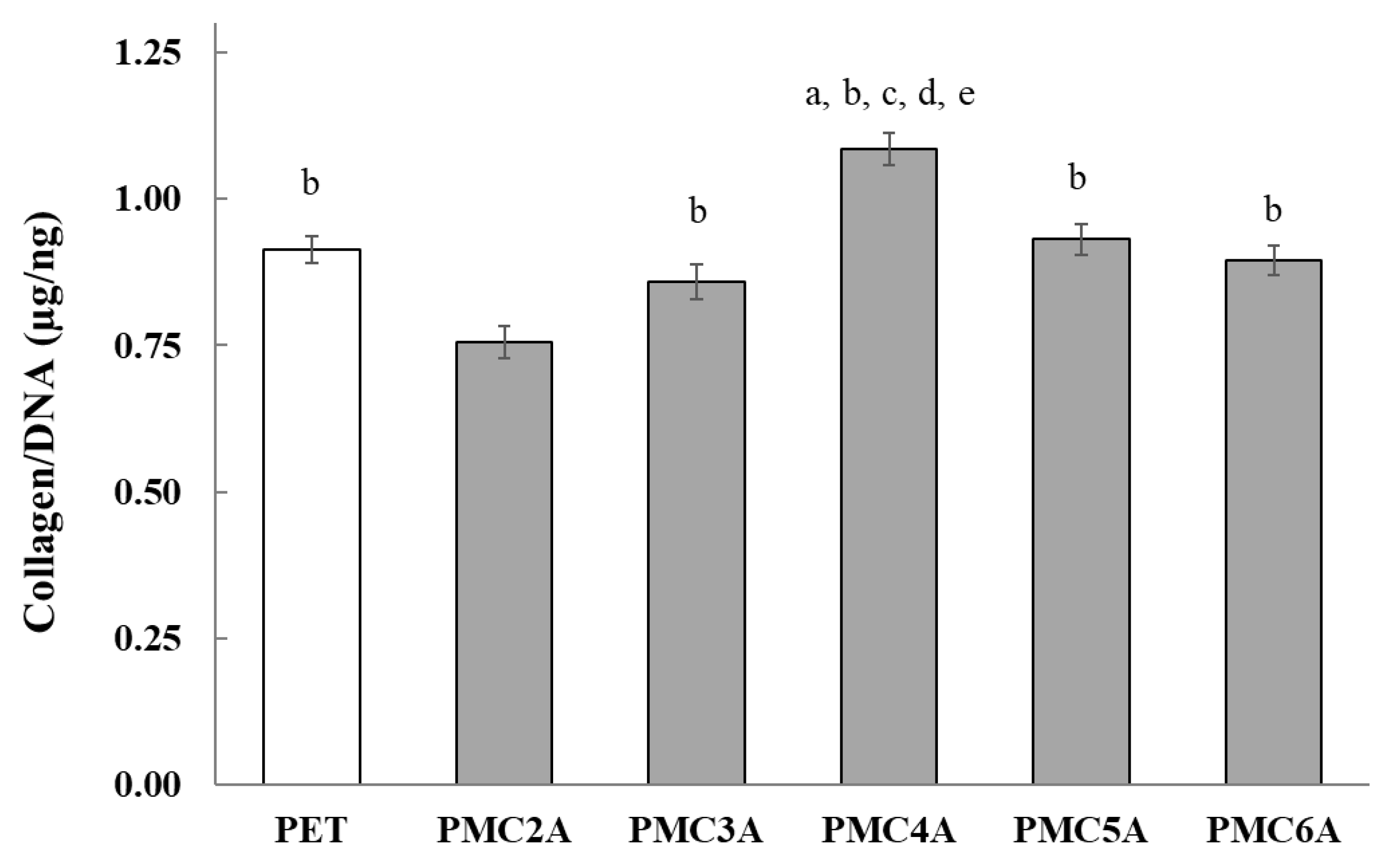
3.7. Total Collagen Measurement
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schindhelm, K.; Milthorpe, B.K. Overview of biomaterials. Australas. Phys. Eng. Sci. Med. 1986, 9, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Teo, A.J.; Mishra, A.; Park, I.; Kim, Y.J.; Park, W.T.; Yoon, Y.J. Polymeric biomaterials for medical implants and devices. ACS Biomater. Sci. Eng. 2016, 2, 454–472. [Google Scholar] [CrossRef]
- Saini, M. Implant biomaterials: A comprehensive review. World J. Clin. Cases 2015, 3, 52–57. [Google Scholar] [CrossRef]
- Ou, S.F.; Chen, C.S.; Hosseinkhani, H.; Yu, C.H.; Shen, Y.D.; Ou, K.L. Surface properties of nano-structural silicon-doped carbon films for biomedical applications. Int. J. Nanotechnol. 2013, 10, 945–958. [Google Scholar] [CrossRef]
- Chen, H.; Yuan, L.; Song, W.; Wu, Z.; Li, D. Biocompatible polymer materials: Role of protein-surface interactions. Prog. Polym. Sci. 2008, 33, 1059–1087. [Google Scholar] [CrossRef]
- Rahmati, M.; Silva, E.A.; Reseland, J.E.; Heyward, C.A.; Haugen, H.J. Biological responses to physicochemical properties of biomaterial surface. Chem. Soc. Rev. 2020, 49, 5178–5224. [Google Scholar] [CrossRef]
- Cai, S.; Wu, C.; Yang, W.; Liang, W.; Yu, H.; Liu, L. Recent advance in surface modification for regulating cell adhesion and behaviors. Nanotechnol. Rev. 2020, 9, 971–989. [Google Scholar] [CrossRef]
- Wong, T.; McGrath, J.A.; Navsaria, H. The role of fibroblasts in tissue engineering and regeneration. Br. J. Dermatol. 2007, 156, 1149–1155. [Google Scholar] [CrossRef]
- Sriram, G.; Bigliardi, P.L.; Bigliardi-Qi, M. Fibroblast heterogeneity and its implications for engineering organotypic skin models in vitro. Eur. J. Cell Biol. 2015, 94, 483–512. [Google Scholar] [CrossRef] [PubMed]
- Ko, U.H.; Choi, J.; Choung, J.; Moon, S.; Shin, J.H. Physicochemically tuned myofibroblasts for wound healing strategy. Sci. Rep. 2019, 9, 16070. [Google Scholar] [CrossRef]
- Mori, L.; Bellini, A.; Stacey, M.A.; Schmidt, M.; Mattoli, S. Fibrocytes contribute to the myofibroblast population in wounded skin and originate from the bone marrow. Exp. Cell Res. 2005, 304, 81–90. [Google Scholar] [CrossRef]
- Bharath Rao, K.; Malathi, N.; Narashiman, S.; Rajan, S.T. Evaluation of myofibroblasts by expression of alpha smooth muscle actin: A marker in fibrosis, dysplasia and carcinoma. J. Clin. Diagn. Res. 2014, 8, 14–17. [Google Scholar] [CrossRef]
- Honda, E.; Park, A.M.; Yoshida, K.; Tabuchi, M.; Munakata, H. Myofibroblasts: Biochemical and proteomic approaches to fibrosis. Tohoku J. Exp. Med. 2013, 230, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Witherel, C.E.; Abebayehu, D.; Barker, T.H.; Spiller, K.L. Macrophage and fibroblast interactions in biomaterial-mediated fibrosis. Adv. Healthc. Mater. 2019, 8, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Kerch, G. Polymer hydration and stiffness at biointerfaces and related cellular processes. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yan, C.; Zheng, Z. Functional polymer surfaces for controlling cell behaviors. Mater. Today 2018, 21, 38–59. [Google Scholar] [CrossRef]
- Bacakova, L.; Filova, E.; Parizek, M.; Ruml, T.; Svorcik, V. Modulation of cell adhesion, proliferation and differentiation on materials designed for body implants. Biotechnol. Adv. 2011, 29, 739–767. [Google Scholar] [CrossRef]
- Wang, X.; Hu, X.; Dulinska-Molak, I.; Kawazoe, N.; Yang, Y.; Chen, G. Discriminating the independent influence of cell adhesion and spreading area on stem cell fate determination using micropatterned surfaces. Sci. Rep. 2016, 6, 28708. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, M.A.; Modzelewska, K.; Kwong, L.; Keely, P.J. Focal adhesion regulation of cell behavior. Biochim. Biophys. Acta Mol. Cell Res. 2004, 1692, 103–119. [Google Scholar] [CrossRef]
- Wysotzki, P.; Gimsa, J. Surface coatings modulate the differences in the adhesion forces of eukaryotic and prokaryotic cells as detected by single cell force microscopy. Int. J. Biomater. 2019, 2019, 7024259. [Google Scholar] [CrossRef]
- Nedela, O.; Slepicka, P.; Švorcík, V. Surface modification of polymer substrates for biomedical applications. Materials 2017, 10, 1115. [Google Scholar] [CrossRef]
- Ye, Z.; Hiroyasu, K.; Yuya, S.; Kano, M.; Tada-Aki, K.; Shimizu, Y. MPC polymer regulates fibrous tissue formation by modulating cell adhesion to the biomaterial surface. Dent. Mater. J. 2010, 29, 518–528. [Google Scholar] [CrossRef][Green Version]
- Tanaka, M.; Motomura, T.; Kawada, M.; Anzai, T.; Kasori, Y.; Shiroya, T.; Shimura, K.; Onishi, M.; Mochizuki, A. Blood compatible aspects of poly(2-methoxyethylacrylate) (PMEA)-relationship between protein adsorption and platelet adhesion on PMEA surface. Biomaterials 2000, 21, 1471–1481. [Google Scholar] [CrossRef]
- Hoshiba, T.; Nikaido, M.; Tanaka, M. Characterization of the attachment mechanisms of tissue-derived cell lines to blood-compatible polymers. Adv. Healthc. Mater. 2014, 3, 775–784. [Google Scholar] [CrossRef]
- Hoshiba, T.; Nemoto, E.; Sato, K.; Orui, T.; Otaki, T.; Yoshihiro, A.; Tanaka, M. Regulation of the contribution of integrin to Cell attachment on poly(2-methoxyethyl acrylate) (PMEA) analogous polymers for attachment-based cell enrichment. PLoS ONE 2015, 10, e0136066. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Hayashi, T.; Morita, S. The roles of water molecules at the biointerface of medical polymers. Polym. J. 2013, 45, 701–710. [Google Scholar] [CrossRef]
- Miwa, Y.; Ishida, H.; Saitô, H.; Tanaka, M.; Mochizuki, A. Network structures and dynamics of dry and swollen poly(acrylate)s. Characterization of high- and low-frequency motions as revealed by suppressed or recovered intensities (SRI) analysis of 13C NMR. Polymer 2009, 50, 6091–6099. [Google Scholar] [CrossRef]
- Morita, S.; Tanaka, M.; Ozaki, Y. Time-resolved in situ ATR-IR observations of the process of sorption of water into a poly(2-methoxyethyl acrylate) film. Langmuir 2007, 23, 3750–3761. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Mochizuki, A. Effect of water structure on blood compatibility—Thermal analysis of water in poly(meth)acrylate. J. Biomed. Mater. Res. Part A 2004, 68, 684–695. [Google Scholar] [CrossRef]
- Tanaka, M.; Sato, K.; Kitakami, E.; Kobayashi, S.; Hoshiba, T.; Fukushima, K. Design of biocompatible and biodegradable polymers based on intermediate water concept. Polym. J. 2015, 47, 114–121. [Google Scholar] [CrossRef]
- Tanaka, M.; Kobayashi, S.; Murakami, D.; Aratsu, F.; Kashiwazaki, A.; Hoshiba, T.; Fukushima, K. Design of polymeric biomaterials: The “Intermediate water concept.”. Bull. Chem. Soc. Jpn. 2019, 92, 2043–2057. [Google Scholar] [CrossRef]
- Hoshiba, T.; Nemoto, E.; Sato, K.; Maruyama, H.; Endo, C.; Tanaka, M. Promotion of adipogenesis of 3T3-L1 cells on protein adsorption-suppressing poly(2-methoxyethyl acrylate) analogs. Biomacromolecules 2016, 17, 3808–3815. [Google Scholar] [CrossRef]
- Kobayashi, S.; Wakui, M.; Iwata, Y.; Tanaka, M. Poly(ω-methoxyalkyl acrylate)s: Nonthrombogenic polymer family with tunable protein adsorption. Biomacromolecules 2017, 18, 4214–4223. [Google Scholar] [CrossRef] [PubMed]
- Jonkman, J.E.; Cathcart, J.A.; Xu, F.; Bartolini, M.E.; Amon, J.E.; Stevens, K.M.; Colarusso, P. An introduction to the wound healing assay using live-cell microscopy. Cell Adhes. Migr. 2014, 8, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Satyam, A.; Fan, X.; Collin, E.; Rochev, Y.; Rodriguez, B.J.; Gorelov, A.; Dillon, S.; Joshi, L.; Raghunath, M.; et al. Macromolecularly crowded in vitro microenvironments accelerate the production of extracellular matrix-rich supramolecular assemblies. Sci. Rep. 2015, 5, 8729. [Google Scholar] [CrossRef] [PubMed]
- Bachhuka, A.; Hayball, J.; Smith, L.E.; Vasilev, K. Effect of surface chemical functionalities on collagen deposition by primary human dermal fibroblasts. ACS Appl. Mater. Interfaces 2015, 7, 23767–23775. [Google Scholar] [CrossRef]
- Bathawab, F.; Bennett, M.; Cantini, M.; Reboud, J.; Dalby, M.J.; Salmerón-Sánchez, M. Lateral chain length in polyalkyl acrylates determines the mobility of fibronectin at the cell/material interface. Langmuir 2016, 32, 800–809. [Google Scholar] [CrossRef]
- Xing, F.; Li, L.; Zhou, C.; Long, C.; Wu, L.; Lei, H.; Kong, Q.; Fan, Y.; Xiang, Z.; Zhang, X. Regulation and directing stem cell fate by tissue engineering functional microenvironments: Scaffold physical and chemical cues. Stem Cells Int. 2019, 2019, 2180925. [Google Scholar] [CrossRef]
- Guerra, N.B.; González-García, C.; Llopis, V.; Rodríguez-Hernández, J.C.; Moratal, D.; Rico, P.; Salmerón-Sánchez, M. Subtle variations in polymer chemistry modulate substrate stiffness and fibronectin activity. Soft Matter 2010, 6, 4748–4755. [Google Scholar] [CrossRef]
- Wong, J.Y.; Leach, J.B.; Brown, X.Q. Balance of chemistry, topography, and mechanics at the cell-biomaterial interface: Issues and challenges for assessing the role of substrate mechanics on cell response. Surf. Sci. 2004, 570, 119–133. [Google Scholar] [CrossRef]
- Ngandu Mpoyi, E.; Cantini, M.; Reynolds, P.M.; Gadegaard, N.; Dalby, M.J.; Salmerón-Sánchez, M. Protein adsorption as a key mediator in the nanotopographical control of cell behavior. ACS Nano 2016, 10, 6638–6647. [Google Scholar] [CrossRef] [PubMed]
- Parisi, L.; Toffoli, A.; Ghezzi, B.; Mozzoni, B.; Lumetti, S.; Macaluso, G.M. A glance on the role of fibronectin in controlling cell response at biomaterial interface. Jpn. Dent. Sci. Rev. 2020, 56, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Khalili, A.A.; Ahmad, M.R. A Review of cell adhesion studies for biomedical and biological applications. Int. J. Mol. Sci. 2015, 16, 18149–18184. [Google Scholar] [CrossRef]
- Kanchanawong, P.; Shtengel, G.; Pasapera, A.M.; Ramko, E.B.; Davidson, M.W.; Hess, H.F.; Waterman, C.M. Nanoscale architecture of integrin-based cell adhesions. Nature 2010, 468, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, W.H.; Liddington, R.C.; Critchley, D.R. The structure and regulation of vinculin. Trends Cell Biol. 2006, 16, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Sumida, Y.; Sanzen, N.; Sekiguchi, K. Laminin-10/11 and fibronectin differentially regulate integrin-dependent Rho and Rac activation via p130Cas-CrkII-DOCK180 pathway. J. Biol. Chem. 2001, 276, 27090–27097. [Google Scholar] [CrossRef]
- Hinz, B.; Celetta, G.; Tomasek, J.J.; Gabbiani, G.; Chaponnier, C. Alpha-smooth muscle actin expression upregulates fibroblast contractile activity. Mol. Biol. Cell 2001, 12, 2730–2741. [Google Scholar] [CrossRef]
- Shinde, A.V.; Humeres, C.; Frangogiannis, N.G. The role of α-smooth muscle actin in fibroblast-mediated matrix contraction and remodeling. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 298–309. [Google Scholar] [CrossRef]
- Franz, M.; Spiegel, K.; Umbreit, C.; Richter, P.; Codina-Canet, C.; Berndt, A.; Altendorf-Hofmann, A.; Koscielny, S.; Hyckel, P.; Kosmehl, H.; et al. Expression of Snail is associated with myofibroblast phenotype development in oral squamous cell carcinoma. Histochem. Cell Biol. 2009, 131, 651–660. [Google Scholar] [CrossRef]
- Cheng, F.; Shen, Y.; Mohanasundaram, P.; Lindström, M.; Ivaska, J.; Ny, T.; Erikss, J.E. Vimentin coordinates fibroblast proliferation and keratinocyte differentiation in wound healing via TGF-β-Slug signaling. Proc. Natl. Acad. Sci. USA 2016, 113, E4320–E4327. [Google Scholar] [CrossRef]
- Tottoli, E.M.; Dorati, R.; Genta, I.; Chiesa, E.; Pisani, S.; Conti, B. Skin wound healing process and new emerging technologies for skin wound care and regeneration. Pharmaceutics 2020, 12, 735. [Google Scholar] [CrossRef]
- Rangaraj, A.; Harding, K.; Leaper, D. Role of collagen in wound management. Drug Invent. Today 2020, 13, 55–57. [Google Scholar]
- Kennedy, K.M.; Bhaw-Luximon, A.; Jhurry, D. Cell-matrix mechanical interaction in electrospun polymeric scaffolds for tissue engineering: Implications for scaffold design and performance. Acta Biomater. 2017, 50, 41–55. [Google Scholar] [CrossRef]
- Tse, J.R.; Engler, A.J. Stiffness gradients mimicking in vivo tissue variation regulate mesenchymal stem cell fate. PLoS ONE 2011, 6, e15978. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Mao, Z.; Wu, J.; Guo, Y.; Ren, T.; Gao, C. Directional cell migration through cell-cell interaction on polyelectrolyte multilayers with swelling gradients. Biomaterials 2013, 34, 975–984. [Google Scholar] [CrossRef] [PubMed]
- Tusan, C.G.; Man, Y.H.; Zarkoob, H.; Johnston, D.A.; Andriotis, O.G.; Thurner, P.J.; Yang, S.; Sander, E.A.; Gentleman, E.; Sengers, B.G.; et al. Collective cell behavior in mechanosensing of substrate thickness. Biophys. J. 2018, 114, 2743–2755. [Google Scholar] [CrossRef] [PubMed]
- Hoshiba, T.; Orui, T.; Endo, C.; Sato, K.; Yoshihiro, A.; Minagawa, Y.; Tanaka, M. Adhesion-based simple capture and recovery of circulating tumor cells using a blood-compatible and thermo-responsive polymer-coated substrate. RSC Adv. 2016, 6, 89103–89112. [Google Scholar] [CrossRef]
- Okur, M.E.; Karantas, I.D.; Şenyiğit, Z.; Üstündağ Okur, N.; Siafaka, P.I. Recent trends on wound management: New therapeutic choices based on polymeric carriers. Asian J. Pharm. Sci. 2020, 15, 661–684. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scheme | Mn | Mw/ Mn | Tg drya | Tg weta | IW b | NFW c |
|---|---|---|---|---|---|---|
| (kg/mol) | (°C) | (°C) | (wt.%) | (wt.%) | ||
| PMC2A | 32 | 1.3 | −35 | −51 | 3.7 | 2.5 |
| PMC3A | 33 | 2.7 | −48 | −58 | 2.8 | 3.1 |
| PMC4A | 24 | 2.4 | −65 | −67 | 1.7 | 1.3 |
| PMC5A | 31 | 2.8 | −74 | −78 | 1.0 | 1.5 |
| PMC6A | 42 | 2.7 | −77 | −78 | 0.8 | 1.3 |
| Samples | Contact Angle (°) | |
|---|---|---|
| Sessile Water Drops a | Captive Air Bubble a | |
| PET | 86.4 (± 1.1) | 129.2 (± 1.0) |
| PMC2A | 44.2 (± 0.6) | 133.6 (± 1.0) |
| PMC3A | 51.1 (± 0.4) | 129.5 (± 1.7) |
| PMC4A | 54.4 (± 0.7) | 125.7 (± 1.7) |
| PMC5A | 57.3 (± 0.2) | 120.3 (± 0.6) |
| PMC6A | 61.0 (± 0.6) | 119.4 (± 3.7) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anjum, R.; Nishida, K.; Matsumoto, H.; Murakami, D.; Kobayashi, S.; Anada, T.; Tanaka, M. Attachment and Growth of Fibroblast Cells on Poly (2-Methoxyethyl Acrylate) Analog Polymers as Coating Materials. Coatings 2021, 11, 461. https://doi.org/10.3390/coatings11040461
Anjum R, Nishida K, Matsumoto H, Murakami D, Kobayashi S, Anada T, Tanaka M. Attachment and Growth of Fibroblast Cells on Poly (2-Methoxyethyl Acrylate) Analog Polymers as Coating Materials. Coatings. 2021; 11(4):461. https://doi.org/10.3390/coatings11040461
Chicago/Turabian StyleAnjum, Rubaiya, Kei Nishida, Haruka Matsumoto, Daiki Murakami, Shingo Kobayashi, Takahisa Anada, and Masaru Tanaka. 2021. "Attachment and Growth of Fibroblast Cells on Poly (2-Methoxyethyl Acrylate) Analog Polymers as Coating Materials" Coatings 11, no. 4: 461. https://doi.org/10.3390/coatings11040461
APA StyleAnjum, R., Nishida, K., Matsumoto, H., Murakami, D., Kobayashi, S., Anada, T., & Tanaka, M. (2021). Attachment and Growth of Fibroblast Cells on Poly (2-Methoxyethyl Acrylate) Analog Polymers as Coating Materials. Coatings, 11(4), 461. https://doi.org/10.3390/coatings11040461