
Study on the Structural Characteristics of Bird Necks and Their Static Motion Features in the Sagittal Plane

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects and Experimental Setup
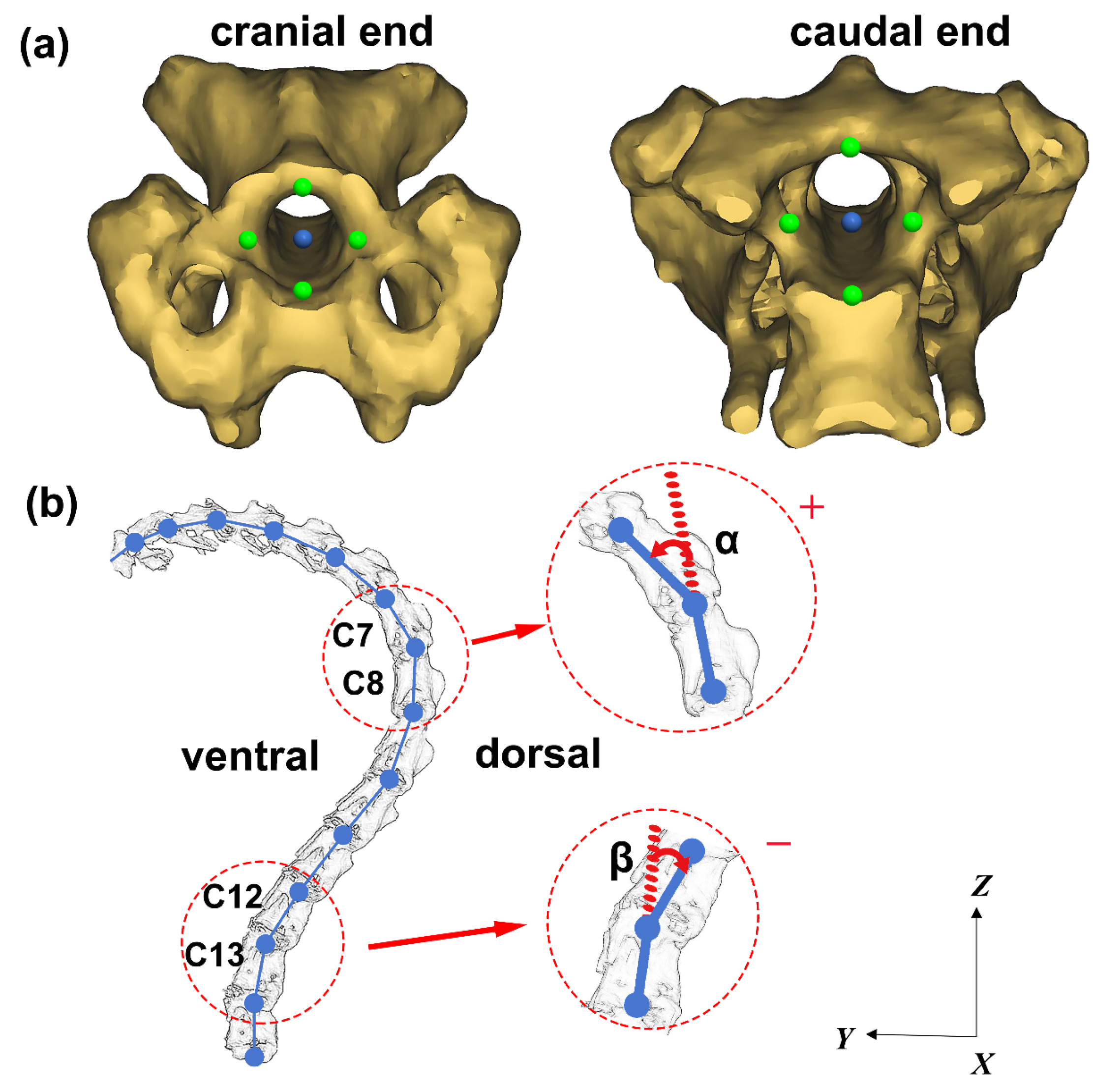
2.2. Structural Parameters Measurement
2.3. Motion Measurement of Articular Process Alignment
2.4. Measurement of Ventroflexion/Dorsiflexion Motion
3. Results
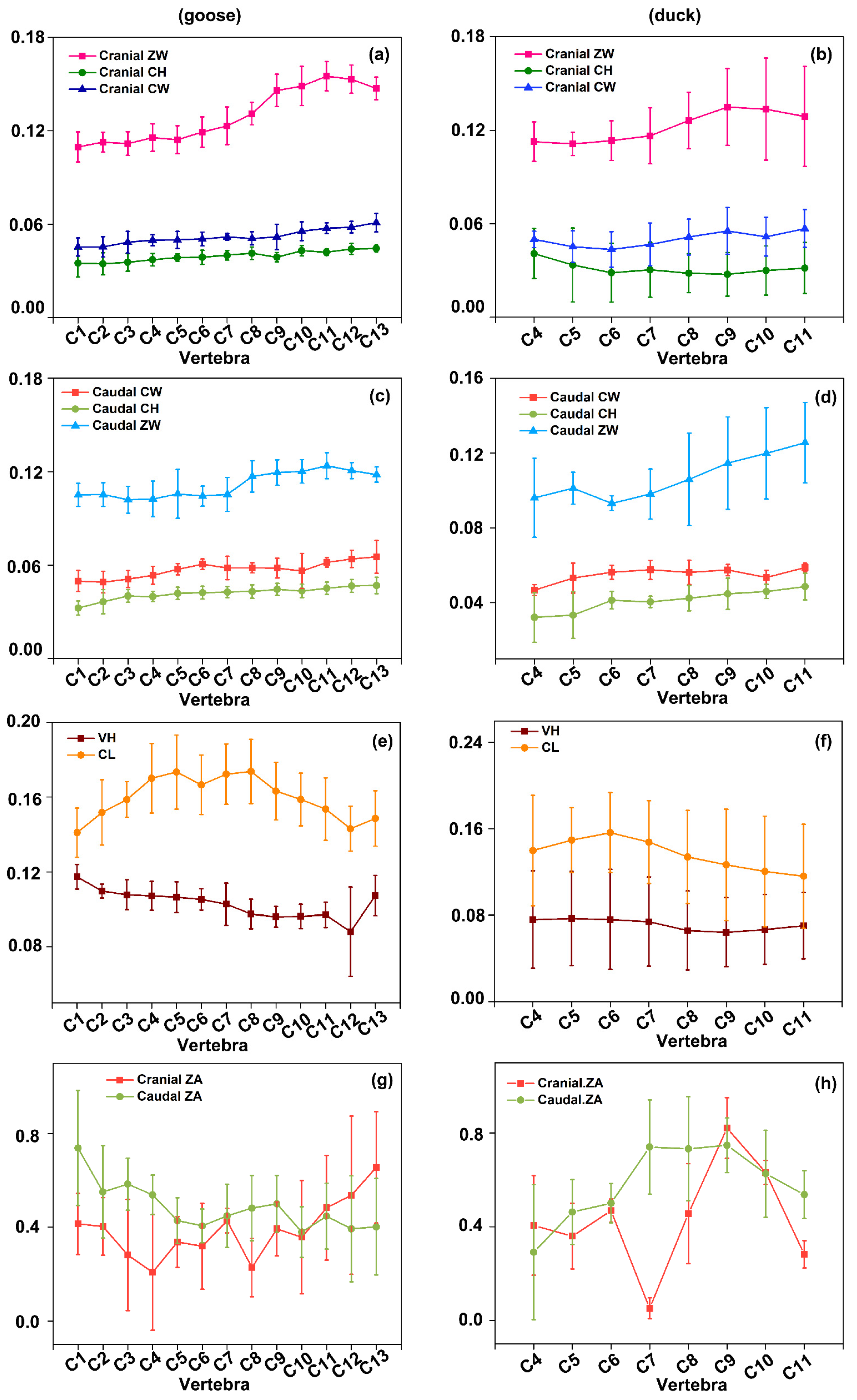
3.1. Structural Parameter Measurement
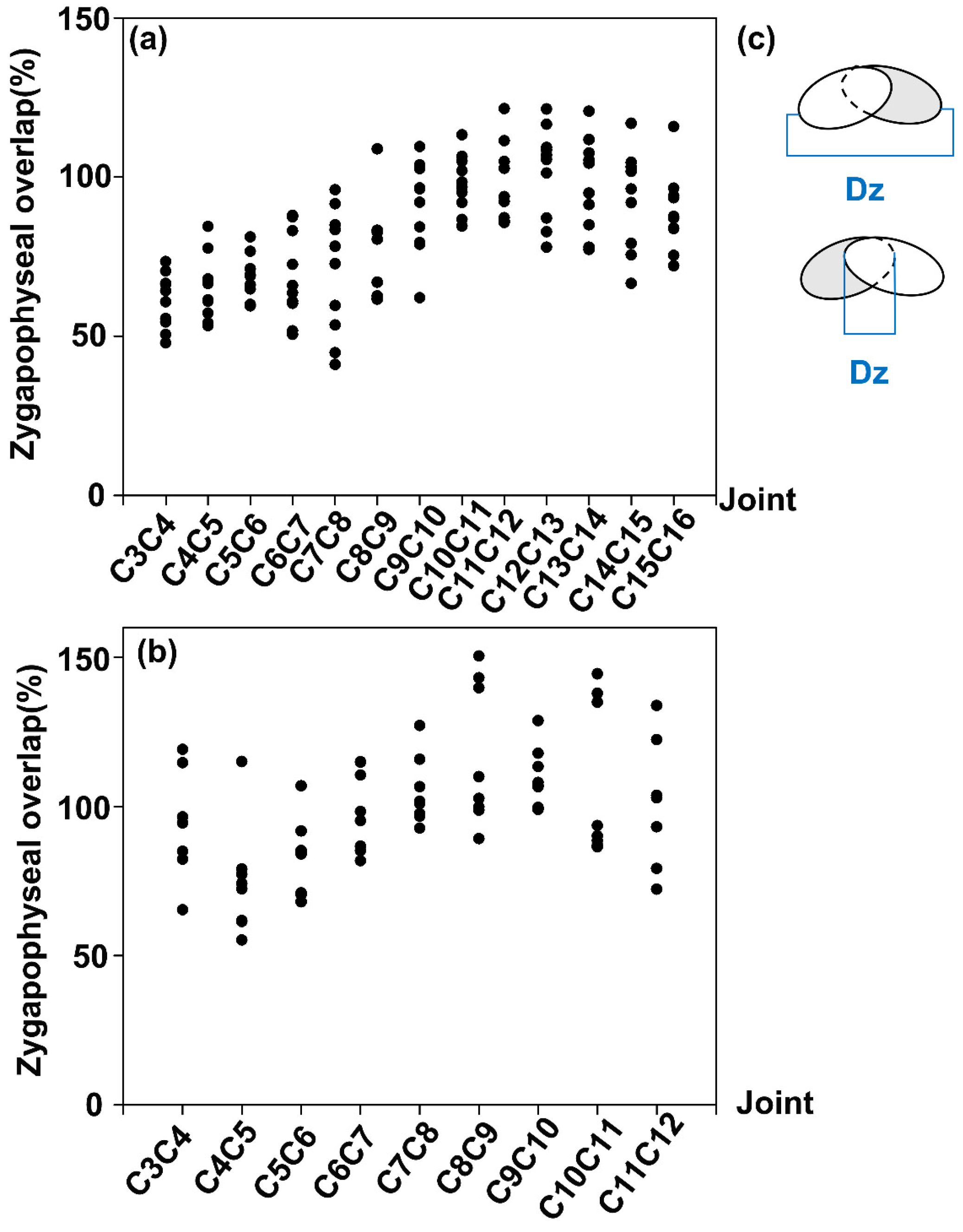
3.2. Analysis of the Zygapophyseal Overlaps of Vertebrae Joints
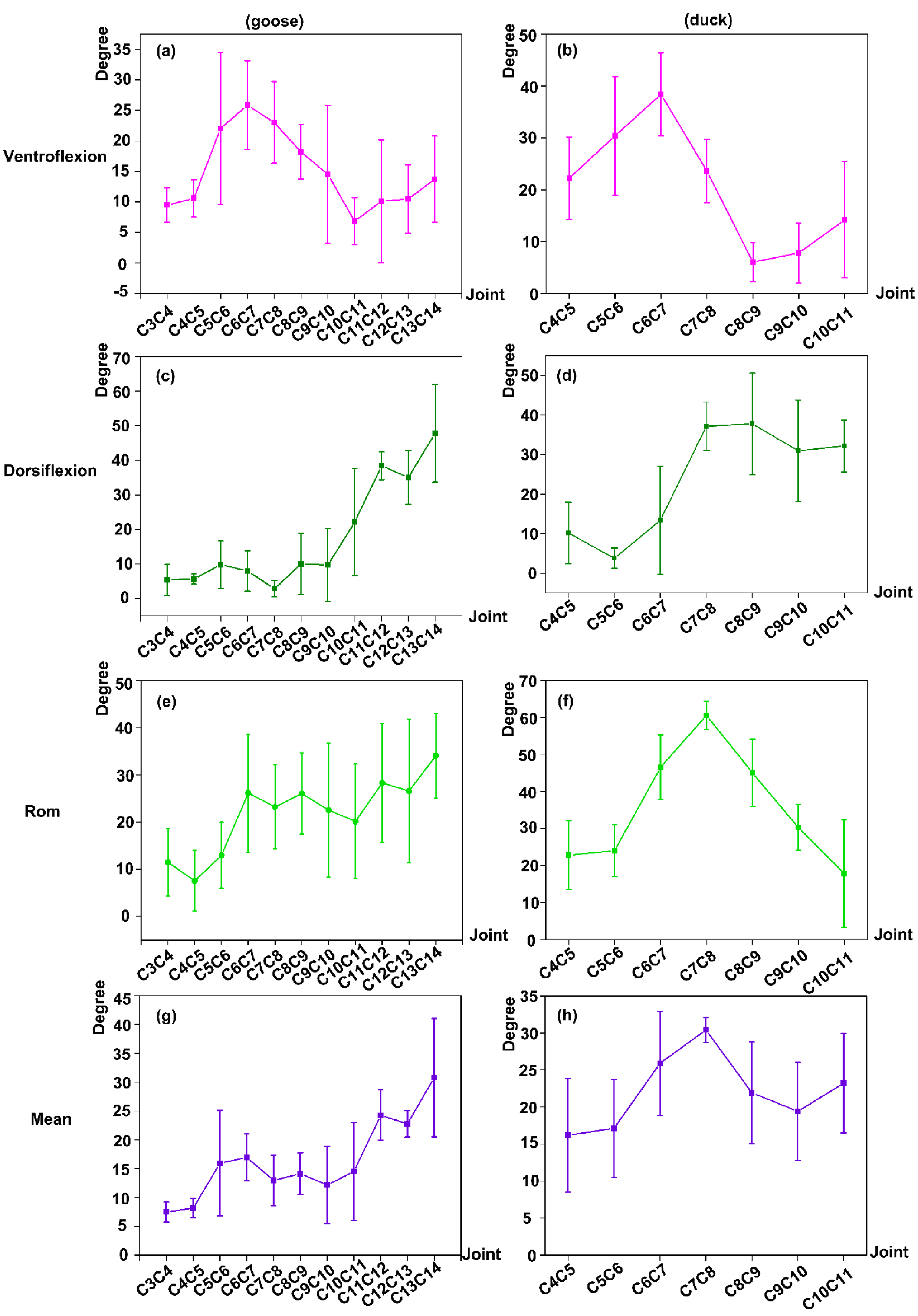
3.3. Ventroflexion/Dorsiflexion Movement Measurement
3.3.1. Ventroflexion RoM
3.3.2. Dorsiflexion Exercise
3.3.3. The Overall RoM in the Sagittal Plane
3.3.4. The Absolute Mean Value of Ventroflexion and Dorsiflexion Exercise (Mean)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Ethical Approval
References
- Heers, A.M.; Dial, K.P. From extant to extinct: Locomotor ontogeny and the evolution of avian flight. Trends Ecol. Evol. 2012, 27, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Rashid, D.J.; Chapman, S.C.; Larsson, H.C.E.; Organ, C.L.; Bebin, A.G.; Merzdorf, C.S.; Bradley, R.; Horner, J.R. From dinosaurs to birds: A tail of evolution. EvoDevo 2014, 5, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, P.R.; Grant, B.R. Unpredictable evolution in a 30-year study of Darwin’sfinches. Science 2002, 296, 707–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellairs, R.; Osmond, M. Atlas of Chick Development, 3rd ed.; Academic Press: Cambridge, UK, 2005. [Google Scholar]
- Kisia, S.M. Vertebrates: Structures and Functions; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Lovette, I.J.; Fitzpatrick, J.W. Handbook of Bird Biology (Cornell Lab of Ornithology), 3rd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2016; pp. 169–214. [Google Scholar]
- Tambussi, C.P.; de Mendoza, R.; Degrange, F.J.; Picasso, M.B. Flexibility along the neck of the neogene terror bird Andalgalornis steulleti (Aves Phorusrhacidae). PLoS ONE 2012, 7, e37701. [Google Scholar] [CrossRef] [Green Version]
- Marek, R.D.; Falkingham, P.L.; Benson, R.B.J.; Gardiner, J.D.; Maddox, T.W.; Bates, K.T. Evolutionary versatility of the avian neck. P Roy Soc. B-Biol. Sci. 2021, 288, 20203150. [Google Scholar]
- Krings, M.; Nyakatura, J.A.; Fischer, M.S.; Wagner, H. The cervical spine of the american barn owl (tyto furcata pratincola): I. anatomy of the vertebrae and regionalization in their s-shaped arrangement. PLoS ONE 2014, 9, e91653. [Google Scholar] [CrossRef] [Green Version]
- Terray, L.; Plateau, O.; Abourachid, A.; Bohmer, C.; Delapre, A.; de la Bernardie, X.; Cornette, R. Modularity of the neck in birds (Aves). Evol. Biol. 2020, 47, 97–110. [Google Scholar] [CrossRef]
- Kambic, R.E.; Biewener, A.A.; Pierce, S.E. Experimental determination of three-dimensional cervical joint mobility in the avian neck. Front. Zool. 2017, 14, 37. [Google Scholar] [CrossRef] [Green Version]
- Katzir, G.; Schechtman, E.; Carmi, N.; Weihs, D. Head stabilization in herons. J. Comp. Physiol. A 2001, 187, 423–432. [Google Scholar]
- Angelique, H.J.; Leeuw, V.D.; Bout, R.G.; Zweer, G.A. Control of the Cranio-Cervical System During Feeding in Birds. Am. Zool. 2001, 41, 1352–1363. [Google Scholar]
- Krings, M.; Nyakatura, J.A.; Boumans, M.L.L.M.; Fischer, M.S.; Wagner, H. Barn owls maximize head rotations by a combination of yawing and rolling in functionally diverse regions of the neck. J. Anat. 2017, 231, 12–22. [Google Scholar] [CrossRef] [Green Version]
- Russell, A.P.; Bentley, A.D. Opisthotonic head displacement in the domestic chicken and its bearing on the ‘dead bird’ posture of non-avialan dinosaurs. J. Zool. 2016, 298, 20–29. [Google Scholar] [CrossRef]
- Kambic, R.E.; Roberts, T.J.; Gatesy, S.M. Guineafowl with a twist: Asymmetric limb control in steady bipedal locomotion. J. Exp. Biol. 2015, 218, 3836–3844. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.Y.; Liu, X.; Du, Y.Y.; Wang, B.; Zhao, N.; Geng, Z.Y. Green forage and fattening duration differentially modulate cecal microbiome of Wanxi white geese. PLoS ONE 2018, 13, e0204210. [Google Scholar] [CrossRef]
- German, D.P.; Horn, M.H. Gut length and mass in herbivorous and carnivorous prickleback fishes (Teleostei: Stichaeidae): Ontogenetic, dietary, and phylogenetic effects. Mar. Biol. 2006, 148, 1123–1134. [Google Scholar] [CrossRef]
- Cobley, M.J.; Rayfield, E.J.; Barrett, P.M. Inter-vertebral flexibility of the ostrich neck: Implications for estimating sauropod neck flexibility. PLoS ONE 2013, 8, e72187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boszczyk, B.M.; Boszczyk, A.A.; Putz, R. Comparative and functional anatomy of the mammalian lumbar spine. Anat. Rec. 2001, 264, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.E. Preliminary data on the effect of osseous anatomy on ex vivo joint mobility in the equine thoracolumbar region. Equine Vet. J. 2015, 48, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Buchholtz, E.A. Vertebral osteology and swimming style in living and fossil whales (Order: Cetacea). J. Zool. 2001, 253, 175–190. [Google Scholar] [CrossRef]
- Pierce, S.E.; Clack, J.A.; Hutchinson, J.R. Comparative axial morphology in pinnipeds and its correlation with aquatic locomotory behaviour. J. Anat. 2011, 219, 502–514. [Google Scholar] [CrossRef]
- Oliver, J.D.; Jones, K.E.; Hautier, L.; Loughry, W.J.; Pierce, S.E. Vertebral bending mechanics and xenarthrous morphology in the nine-banded armadillo (Dasypus novemcinctus). J. Exp. Biol. 2016, 219, 2991–3002. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n | Direction of the Vertebrae | Cranial | Caudal | VH | CL | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Measuring Structure | CH | CW | ZW | CH | CW | ZW | |||||||||||
| Vertebra | Cn | Cn + 1 | Cn | Cn + 1 | Cn | Cn + 1 | Cn | Cn + 1 | Cn | Cn + 1 | Cn | Cn + 1 | Cn | Cn + 1 | Cn | Cn + 1 | |
| 3 | Ventroflexion | − | − | − | + | − | − | − | + | − | − | − | − | − | − | − | − |
| Dorsiflexion | + | − | − | + | − | − | − | + | − | − | − | − | − | − | − | − | |
| Mean | + | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | |
| Rom | + | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | |
| 4 | Ventroflexion | − | − | − | − | − | − | − | − | − | − | + | + | − | + | − | − |
| Dorsiflexion | − | − | − | − | − | − | + | − | − | − | − | + | − | + | − | − | |
| Mean | − | − | − | − | − | − | − | − | − | − | − | + | − | + | − | − | |
| Rom | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + | − | |
| 6 | Ventroflexion | − | − | − | − | − | − | − | + | − | − | − | − | − | − | − | − |
| Dorsiflexion | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | |
| Mean | − | − | − | − | − | − | − | + | − | − | − | − | − | − | − | − | |
| Rom | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | |
| 7 | Ventroflexion | − | − | − | − | − | − | − | − | − | − | + | − | − | − | + | − |
| Dorsiflexion | − | − | − | − | − | − | − | − | + | − | − | − | − | − | − | − | |
| Mean | − | − | − | − | − | − | − | − | − | − | + | − | − | − | + | − | |
| Rom | − | − | − | − | − | − | − | − | − | − | + | − | − | − | + | − | |
| 8 | Ventroflexion | − | − | − | − | − | − | − | − | − | − | − | − | − | + | − | − |
| Dorsiflexion | − | − | − | − | − | − | − | − | − | − | − | − | − | + | − | − | |
| Mean | − | − | − | − | − | − | − | − | − | − | − | − | − | + | − | − | |
| Rom | − | − | − | − | − | − | − | − | − | − | − | − | − | + | − | − | |
| 9 | Ventroflexion | − | − | − | − | − | − | − | − | − | − | − | − | − | + | − | − |
| Dorsiflexion | − | − | − | − | − | − | − | − | − | − | + | − | − | − | − | − | |
| Mean | − | − | − | − | − | − | − | − | − | − | − | − | − | + | − | − | |
| Rom | − | − | − | − | − | − | − | − | − | − | − | − | − | + | − | − | |
| 10 | Ventroflexion | − | − | − | − | − | − | − | − | − | − | + | − | − | − | − | − |
| Dorsiflexion | − | − | − | − | − | − | − | + | − | − | − | − | − | − | − | − | |
| Mean | − | − | − | − | − | − | − | + | − | − | − | − | − | − | − | − | |
| Rom | − | − | − | − | − | − | − | + | − | − | − | − | − | − | − | − | |
| 11 | Ventroflexion | − | − | − | − | − | − | − | − | − | − | − | + | − | − | − | + |
| Dorsiflexion | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | |
| Mean | − | − | − | − | − | − | − | − | − | − | − | + | − | − | − | + | |
| Rom | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | |
| 12 | Ventroflexion | − | − | − | − | − | − | − | − | − | − | − | − | + | − | − | − |
| Dorsiflexion | − | − | − | − | − | − | − | − | + | − | − | − | − | − | − | − | |
| Mean | − | − | − | − | + | − | − | + | − | − | + | + | − | − | + | − | |
| Rom | − | − | − | − | − | − | − | − | + | − | − | − | − | − | − | − | |
| 13 | Ventroflexion | − | − | − | − | − | − | − | − | + | + | − | − | − | − | − | − |
| Dorsiflexion | − | − | − | − | − | − | − | − | + | + | − | − | − | − | − | − | |
| Mean | − | − | − | − | − | − | − | − | + | + | − | − | − | − | − | − | |
| Rom | − | − | − | − | − | − | − | − | + | + | − | − | − | − | − | − | |
| n | Direction of the Vertebrae | Cranial | Caudal | VH | CL | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Measuring structure | CH | CW | ZW | CH | CW | ZW | |||||||||||
| Vertebra | Cn | Cn + 1 | Cn | Cn + 1 | Cn | Cn + 1 | Cn | Cn + 1 | Cn | Cn + 1 | Cn | Cn + 1 | Cn | Cn + 1 | Cn | Cn + 1 | |
| 4 | Ventroflexion | − | + | − | − | − | − | − | − | − | − | + | − | − | − | − | − |
| Dorsiflexion | − | + | − | − | − | − | − | − | − | − | + | − | − | − | − | − | |
| Mean | − | + | − | − | − | − | − | − | − | − | + | − | − | − | − | − | |
| Rom | − | + | − | − | − | − | − | − | − | − | + | − | − | − | − | − | |
| 5 | Ventroflexion | + | − | − | + | − | − | − | − | − | − | − | − | − | − | + | + |
| Dorsiflexion | + | − | − | + | − | − | − | − | − | − | − | − | − | − | + | + | |
| Mean | + | − | − | + | − | − | − | − | − | − | − | − | − | − | + | + | |
| Rom | + | − | − | + | − | − | − | − | − | − | − | − | − | − | + | + | |
| 6 | Ventroflexion | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| Dorsiflexion | − | − | − | + | − | − | − | − | − | − | − | + | + | − | − | − | |
| Mean | − | − | − | + | − | − | − | − | − | − | − | + | + | − | − | − | |
| Rom | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | |
| 7 | Ventroflexion | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| Dorsiflexion | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | |
| Mean | − | − | − | − | − | − | + | − | − | − | − | − | − | − | − | − | |
| Rom | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | |
| 8 | Ventroflexion | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| Dorsiflexion | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | |
| Mean | − | − | − | − | − | − | − | − | + | − | − | − | − | − | − | − | |
| Rom | − | − | − | − | − | − | − | − | + | − | − | − | − | − | − | − | |
| 9 | Ventroflexion | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| Dorsiflexion | + | + | − | − | + | − | − | − | − | − | − | − | + | + | + | − | |
| Mean | + | + | − | − | + | − | − | − | − | − | − | − | + | + | + | − | |
| Rom | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Jia, W.; Zhang, F.; Ma, X.; Qiu, Z.; Qian, Z.; Ren, L.; Guo, Z.; Zhang, Y. Study on the Structural Characteristics of Bird Necks and Their Static Motion Features in the Sagittal Plane. Coatings 2021, 11, 1228. https://doi.org/10.3390/coatings11101228
Wang J, Jia W, Zhang F, Ma X, Qiu Z, Qian Z, Ren L, Guo Z, Zhang Y. Study on the Structural Characteristics of Bird Necks and Their Static Motion Features in the Sagittal Plane. Coatings. 2021; 11(10):1228. https://doi.org/10.3390/coatings11101228
Chicago/Turabian StyleWang, Jiajia, Wenfeng Jia, Fu Zhang, Xiqiang Ma, Zhaomei Qiu, Zhihui Qian, Luquan Ren, Zhijun Guo, and Yakun Zhang. 2021. "Study on the Structural Characteristics of Bird Necks and Their Static Motion Features in the Sagittal Plane" Coatings 11, no. 10: 1228. https://doi.org/10.3390/coatings11101228
APA StyleWang, J., Jia, W., Zhang, F., Ma, X., Qiu, Z., Qian, Z., Ren, L., Guo, Z., & Zhang, Y. (2021). Study on the Structural Characteristics of Bird Necks and Their Static Motion Features in the Sagittal Plane. Coatings, 11(10), 1228. https://doi.org/10.3390/coatings11101228