Overview of Global Trends in Classification, Methods of Preparation and Application of Bacteriocins


 ,
,  ,
,  ,
,
Abstract
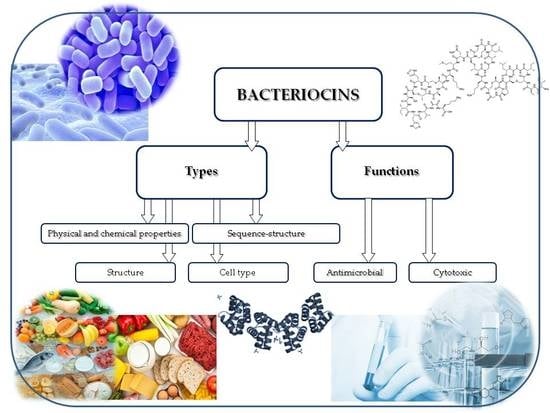
:
1. Introduction
2. Classification of Bacteriocins
2.1. Cell Type Classification
2.1.1. Bacteriocins from Gram-Negative Bacteria
2.1.2. Bacteriocins from Gram-Positive Bacteria
2.2. Structure-Based Classification
2.2.1. Cyclic Structure Classification
2.2.2. Linear Structure Classification
2.3. Sequence-Structure Based Classification
2.4. Classification by Physicochemical Properties of Peptides
3. Sources of Bacteriocins
4. Antimicrobial Activity of Bacteriocins
5. Bacteriocins in Cancer Treatment
6. Nanotechnologies in the Use of Bacteriocins
7. Conclusions
- -
- Search for new bacteriocin-producing strains;
- -
- Improve the productivity of strains (by selection and genetic engineering);
- -
- Search for strains with new properties;
- -
- Study the effect of production conditions on the content of bacteriocins;
- -
- Optimization of production processes (cost reduction and increased product yield);
- -
- Environmentalization of production (introduction of closed production cycles, reduction of waste, the most comprehensive use of components);
- -
- Assessment of environmental, economic, medical risks from scaling up production.
Author Contributions
Funding
Conflicts of Interest
References
- Sugrue, I.; O’Connor, P.M.; Hill, C.; Stanton, C.; Ross, R.P. Actinomyces produces defensin-like bacteriocins (actifensins) with a highly degenerate structure and broad antimicrobial activity. J. Bacteriol. 2020, 202, E00529-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bindiya, E.S.; Bhat, S.G. Marine bacteriocins: A review. J. Bacteriol. Mycol. 2016, 2, 140–147. [Google Scholar]
- Yongkiettrakul, S.; Maneerat, K.; Arechanajan, B.; Malila, Y.; Srimanote, P.; Gottschalk, M.; Visessanguan, W. Antimicrobial susceptibility of Streptococcus suis isolated from diseased pigs, asymptomatic pigs, and human patients in Thailand. BMC Vet. Res. 2019, 15, 5–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, S.N.; Rimmer, M.A.; Turner, K.B.; Phillips, D.A.; Caruana, J.C.; Hervey, W.J.; Leary, D.H.; Walper, S.A. Lactobacillus acidophilus Membrane Vesicles as a Vehicle of Bacteriocin Delivery. Front. Microbiol. 2020, 11, 710. [Google Scholar] [CrossRef]
- Noda, M.; Miyauchi, R.; Danshiitsoodol, N.; Matoba, Y.; Kumagai, T.; Sugiyama, M. Expression of Genes Involved in Bacteriocin Production and Self-Resistance in Lactobacillus Brevis 174A Is Mediated by Two Regulatory Proteins. Appl. Environ. Microbiol. 2018, 84, E02707-17. [Google Scholar] [CrossRef] [Green Version]
- Kumariya, R.; Garsa, A.K.; Rajput, Y.S.; Sood, S.K.; Akhtar, N.; Patel, S. Bacteriocins: Classification, synthesis, mechanism of action and resistance development in food spoilage causing bacteria. Microb. Pathog. 2019, 128, 171–177. [Google Scholar] [CrossRef]
- Ahmad, V.; Khan, M.S.; Jamal, Q.M.S.; Alzohairy, M.A.; Karaawi, M.A.A.; Siddiqui, M.U. Antimicrobial Potential of Bacteriocins: In Therapy, Agriculture and Food Preservation. Int. J. Antimicrob. Agents 2017, 49, 1–11. [Google Scholar] [CrossRef]
- Mbandlwa, P.; Doyle, N.; Hill, C.; Stanton, C.; Ross, R.P. Bacteriocins: Novel Applications in Food, and Human and Animal Health. Ref. Modul. Food Sci. 2020. [Google Scholar] [CrossRef]
- Nishie, M.; Nagao, J.-I.; Sonomoto, K. Antibacterial Peptides “Bacteriocins”: An Overview of Their Diverse Characteristics and Applications. Biocontrol Sci. 2012, 17, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Todorov, S.D.; Chikindas, M.L. Lactic Acid Bacteria Bacteriocins and their Impact on Human Health. In Lactic Acid Bacteria. A Functional Approach; Taylor & Francis Group: Boca Raton, FL, USA, 2020; Volume 5, pp. 1–14. [Google Scholar]
- Munoz, M.; Jaramillo, D.; Melendez, A.; Almeciga-Diaz, C.J.; Sanchez, O.F. Native and Heterologous Production of Bacteriocins from Gram-Positive Microorganisms. Recent Pat. Biotechnol. 2011, 5, 275–277. [Google Scholar] [CrossRef]
- Montalban-Lopez, M.; Sanchez-Hidalgo, M.; Martinez-Bueno, M. Are Bacteriocins Underexploited? NOVEL Applications for OLD Antimicrobials. Curr. Pharm. Biotechnol. 2011, 12, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Zendo, T. Screening and characterization of novel bacteriocins from lactic acid bacteria. Biosci. Biotechnol. Biochem. 2013, 77, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Blinkova, L.P.; Al’tshuler, M.L.; Dorofeeva, E.S.; Gorobest, O.B. Molecular basis of production and action of bacteriocins (In Russian). J. Microbiol. Epidemiol. Immunobiol. 2007, 2, 97–104. [Google Scholar]
- Karpiński, T.M.; Szkaradkiewicz, A.K. Characteristic of bacteriocines and their application. Pol. J. Microbiol. 2013, 62, 223–235. [Google Scholar] [CrossRef]
- Egorov, N.S.; Baranova, I.P. Bacteriocins. Formation, properties, application. Antibiot. Chemother. 1999, 6, 33–40. [Google Scholar]
- Ge, J.; Kang, J.; Ping, W. Effect of Acetic Acid on Bacteriocin Production by Gram-Positive Bacteria. J. Microbiol. Biotechnol. 2019, 29, 1341–1348. [Google Scholar] [CrossRef]
- Rebuffat, S. Microcins and other bacteriocins: Bridging the gaps between killing stategies, ecology and applications. In The Bacteriocins: Current Knowledge and Future Prospects; Caister Academic Press: Wymondham, UK, 2016; pp. 11–34. [Google Scholar]
- Wencewicz, T.A.; Miller, M.J. Sideromycins as Pathogen-Targeted Antibiotics. In Antibacterials 2017. Topics in Medicinal Chemistry 26; Springer: Cham, Switzerland, 2017; pp. 151–183. [Google Scholar]
- Garcia-Gutierrez, E.; O’Connor, P.M.; Colquhoun, I.J.; Vior, N.M.; Rodríguez, J.M.; Mayer, M.J.; Cotter, P.D.; Narbad, A. Production of multiple bacteriocins, including the novel bacteriocin gassericin M, by Lactobacillus gasseri LM19, a strain isolated from human milk. Appl. Microbiol. Biotechnol. 2020, 104, 3869–3884. [Google Scholar] [CrossRef] [Green Version]
- Egan, K.; Ross, R.P.; Hill, C. Bacteriocins: Antibiotics in the age of the microbiome. Emerg. Top. Life Sci. 2017, 1, 55–63. [Google Scholar]
- Acedo, J.Z.; Chiorean, S.; Vederas, J.C.; van Belkum, M.J. The expanding structural variety among bacteriocins from Gram-positive bacteria. FEMS Microbiol. Rev. 2018, 42, 805–828. [Google Scholar] [CrossRef]
- Ongey, E.L.; Yassi, H.; Pflugmacher, S.; Neubauer, P. Pharmacological and pharmacokinetic properties of lanthipeptides undergoing clinical studies. Biotechnol. Lett. 2017, 39, 473–482. [Google Scholar] [CrossRef]
- Wiebach, V.; Mainz, A.; Siegert, M.A.J.; Jungmann, N.A.; Lesquame, G.; Tirat, S.; Dreux-Zigha, A.; Aszodi, J.; Le Beller, D.; Süssmuth, R.D. The anti-staphylococcal lipolanthines are ribosomally synthesized lipopeptides. Nat. Chem. Biol. 2018, 14, 652–654. [Google Scholar] [CrossRef] [PubMed]
- Bennallack, P.R.; Griffitts, J.S. Elucidating and engineering thiopeptide biosynthesis. World J. Microbiol. Biotechnol. 2017, 33, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Garvey, M.; Rowan, N.J. Pulsed UV as a potential surface sanitizer in food production processes to ensure consumer safety. Curr. Opin. Food Sci. 2019, 26, 65–70. [Google Scholar] [CrossRef]
- Zou, J.; Jiang, H.; Cheng, H.; Fang, J.; Huang, G. Strategies for screening, purification and characterization of bacteriocins. Int. J. Biol. Macromol. 2018, 117, 781–789. [Google Scholar] [CrossRef]
- Lv, X.; Du, J.; Jie, Y.; Zhang, B.; Bai, F.; Zhao, H.; Li, J. Purification and antibacterial mechanism of fish-borne bacteriocin and its application in shrimp (Penaeus vannamei) for inhibiting Vibrio parahaemolyticus. World J. Microbiol. Biotechnol. 2017, 33, 156–159. [Google Scholar] [CrossRef]
- Meade, E.; Slattery, M.A.; Garvey, M. Bacteriocins, potent antimicrobial peptides and the fight against multi drug resistant species: Resistance is futile? Antibiotics 2020, 9, 32. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V.G.; Pandey, A. New and Future Developments in Microbial Biotechnology and Bioengineering: Microbial Secondary Metabolites Biochemistry and Applications; Elsevier Science: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Zhao, Z.; Orfe, L.H.; Liu, J.; Lu, S.Y.; Besser, T.E.; Call, D.R. Microcin PDI regulation and proteolytic cleavage are unique among known microcins. Sci. Rep. 2017, 16, 42529–42534. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Sieiro, P.; Montalbán-López, M.; Mu, D.; Kuipers, O.P. Bacteriocins of lactic acid bacteria: Extending the family. Appl. Microbiol. Biotechnol. 2016, 100, 2939–2951. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Wang, X.; Zhang, X.; Wu, H.; Zou, Y.; Li, P.; Sun, C.; Xu, W.; Liu, F.; Wang, D. Class III bacteriocin Helveticin-M causes sublethal damage on target cells through impairment of cell wall and membrane. J. Ind. Microbiol. Biotechnol. 2018, 45, 213–227. [Google Scholar] [CrossRef]
- Tracanna, V.; de Jong, A.; Medema, M.H.; Kuipers, O.P. Mining prokaryotes for antimicrobial compounds: From diversity to function. FEMS Microbiol. Rev. 2017, 41, 417–429. [Google Scholar] [CrossRef]
- Lajis, A.F.B. Biomanufacturing process for the production of bacteriocins from Bacillaceae family. Bioresour. Bioprocess. 2020, 7, 8–13. [Google Scholar] [CrossRef]
- Crone, W.J.K.; Vior, N.M.; Santos-Aberturas, J.; Schmitz, L.G.; Leeper, F.J.; Truman, A.W. Dissecting Bottromycin Biosynthesis Using Comparative Untargeted Metabolomics. Angew. Chemie Int. Ed. 2016, 55, 9639–9643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.; Moore, G.; Nodwell, J. Put a bow on it: Knotted antibiotics take center stage. Antibiotics 2019, 8, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makarova, K.S.; Wolf, Y.I.; Karamycheva, S.; Zhang, D.; Aravind, L.; Koonin, E.V. Antimicrobial peptides, polymorphic toxins, and self-nonself recognition systems in archaea: An untapped armory for intermicrobial conflicts. mBio 2019, 10, E00715-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bédard, F.; Biron, E. Recent progress in the chemical synthesis of class II and S-glycosylated bacteriocins. Front. Microbiol. 2018, 9, 1048–1051. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Wang, S.; Yu, X. Cryptand-imidazolium supported total synthesis of the lasso peptide BI-32169 and its d-enantiomer. Commun. Chem. 2019, 55, 3323–3326. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [Green Version]
- Tymoszewska, A.; DIep, D.B.; Wirtek, P.; Aleksandrzak-Piekarczyk, T. The Non-Lantibiotic Bacteriocin Garvicin Q Targets Man-PTS in a Broad Spectrum of Sensitive Bacterial Genera. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Li, P.; Gu, Q. Heterologous expression and purification of plantaricin NC8, a two-peptide bacteriocin against Salmonella spp. from Lactobacillus plantarum ZJ316. Protein Expr. Purif. 2016, 127, 28–34. [Google Scholar] [CrossRef]
- Ovchinnikov, K.V.; Kristiansen, P.E.; Straume, D.; Jensen, M.S.; Aleksandrzak-Piekarczyk, T.; Nes, I.F.; Diep, D.B. The leaderless bacteriocin enterocin K1 is highly potent against Enterococcus faecium: A study on structure, target spectrum and receptor. Front. Microbiol. 2017, 8, 774–779. [Google Scholar] [CrossRef] [Green Version]
- Baquero, F.; Lanza, V.F.; Baquero, M.R.; del Campo, R.; Bravo-Vázquez, D.A. Microcins in Enterobacteriaceae: Peptide Antimicrobials in the Eco-Active Intestinal Chemosphere. Front. Microbiol. 2019, 10, 2261. [Google Scholar] [CrossRef] [PubMed]
- Chiumento, S.; Roblin, C.; Kieffer-Jaquinod, S.; Tachon, S.; Leprètre, C.; Basset, C.; Aditiyarini, D.; Olleik, H.; Nicoletti, C.; Bornet, O.; et al. Ruminococcin C, a promising antibiotic produced by a human gut symbiont. Sci. Adv. 2019, 5, eaaw9969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roblin, C.; Chiumento, S.; Bornet, O.; Nouailler, M.; Müller, C.S.; Jeannot, K.; Basset, C.; Kieffer-Jaquinod, S.; Couté, Y.; Torelli, S.; et al. The unusual structure of Ruminococcin C1 antimicrobial peptide confers clinical properties. Proc. Natl. Acad. Sci. USA 2020, 117, 19168–19177. [Google Scholar] [CrossRef] [PubMed]
- Adedire, O.M.; Odeniyi, O.A. Antimicrobial Activities of Bacteriocin-Like Extracellular Metabolites Produced by Soil Bacteria. UKJPB 2017, 5, 47–54. [Google Scholar] [CrossRef]
- DeCastro, M.E.; Escuder, J.J.R.; Becerra, M.; Rodríguez-Belmonte, E.; González, M.I.S. Archaeal biocommunication in Hot Springs revealed by metagenomics. In Biocommunication of Archaea; Springer: Cham, Switzerland, 2017; pp. 85–101. [Google Scholar]
- Nowakowski, M.; Jaremko, Ł.; Wladyka, B.; Dubin, G.; Ejchart, A.; Mak, P. Spatial attributes of the four-helix bundle group of bacteriocins—The high-resolution structure of BacSp222 in solution. Int. J. Biol. Macromol. 2018, 107, 2715–2724. [Google Scholar] [CrossRef] [Green Version]
- Kyriakou, P.K.; Ekblad, B.; Kristiansen, P.E.; Kaznessis, Y.N. Interactions of a class IIb bacteriocin with a model lipid bilayer, investigated through molecular dynamics simulations. Biochim. Biophys. Acta 2016, 1858, 824–835. [Google Scholar] [CrossRef]
- Chiorean, S.; Vederas, J.C.; van Belkum, M.J. Identification and Heterologous Expression of the sec-Dependent Bacteriocin Faerocin MK from Enterococcus faecium M3K31. Probiot. Antimicrob. Proteins 2018, 10, 142–147. [Google Scholar] [CrossRef]
- De Castro, I.; Mendo, S.; Caetano, T. Antibiotics from Haloarchaea: What Can We Learn from Comparative Genomics? Mar. Biotechnol. 2020, 22, 308–316. [Google Scholar] [CrossRef]
- Sharp, C.; Bray, J.; Housden, N.G.; Maiden, M.C.J.; Kleanthous, C. Diversity and distribution of nuclease bacteriocins in bacterial genomes revealed using Hidden Markov Models. PLoS Comput. Biol. 2017, 13, E1005652. [Google Scholar] [CrossRef] [Green Version]
- Do Nascimento, K.D.O.; Paes, S.D.N.D.; Augusta, I.M. A Review “Clean Labeling”: Applications of Natural Ingredients in Bakery Products. J. Food Nutr. Res. 2018, 6, 285–294. [Google Scholar] [CrossRef]
- Abdulkarim, I.H.; Mohammed, S.S.D.; Orukotan, A.A. Gene Identification for Bacteriocin Production by Lactic Acid Bacteria Isolated from Selected Fermented Foods. J. Biochem. Mol. Biol. Biophys. 2020, 3, 1–12. [Google Scholar]
- Bungenstock, L.; Abdulmawjood, A.; Reich, F. Evaluation of antibacterial properties of lactic acid bacteria from traditionally and industrially produced fermented sausages from Germany. PLoS ONE 2020, 15, E0230345. [Google Scholar] [CrossRef] [PubMed]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.V.; Copetti, M.V. Alternative methods for mould spoilage control in bread and bakery products. Int. Food Res. J. 2019, 26, 737–749. [Google Scholar]
- Ołdak, A.; Zielińska, D. Bacteriocins from lactic acid bacteria as an alternative to antibiotics. Postepy Hig. Med. Dosw. 2017, 71, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.M.; Jung, D.Y.G.; Jin, D.Y.Y.; Jayabalan, D.R.; Yang, D.S.H.; Suh, J.W. Bacteriocins as food preservatives: Challenges and emerging horizons. Crit. Rev. Food Sci. Nutr. 2018, 58, 2743–2767. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Vuai, M.S.; Zhong, M. The role of bacteria in cancer therapy—Enemies in the past, but allies at present. Infect Agent Cancer 2018, 13, 9–16. [Google Scholar] [CrossRef]
- Ekblad, B.; Kyriakou, P.K.; Oppegård, C.; Nissen-Meyer, J.; Kaznessis, Y.N.; Kristiansen, P.E. Structure-Function Analysis of the Two-Peptide Bacteriocin Plantaricin EF. Biochemistry 2016, 55, 5106–5116. [Google Scholar] [CrossRef]
- Romero-Calle, D.; Benevides, R.G.; Góes-Neto, A.; Billington, C. Bacteriophages as alternatives to antibiotics in clinical care. Antibiotics 2019, 8, 138. [Google Scholar] [CrossRef] [Green Version]
- Ovchinnikov, K.V.; Chi, H.; Mehmeti, I.; Holo, H.; Nes, I.F.; Diep, D.B. Novel group of leaderless multipeptide bacteriocins from Gram-positive bacteria. Appl. Environ. Microbiol. 2016, 82, 5216–5224. [Google Scholar] [CrossRef] [Green Version]
- Woraprayote, W.; Malila, Y.; Sorapukdee, S.; Swetwiwathana, A.; Benjakul, S.; Visessanguan, W. Bacteriocins from lactic acid bacteria and their applications in meat and meat products. Meat Sci. 2016, 120, 118–132. [Google Scholar] [CrossRef] [PubMed]
- Andryukov, B.G.; Mikhaylov, V.V.; Besednova, N.N.; Zaporozhets, T.S.; Bynina, M.P.; Matosova, E.V. The Bacteriocinogenic Potential of Marine Microorganisms. Russ. J. Mar. Biol. 2018, 44, 433–441. [Google Scholar] [CrossRef]
- Rather, I.A.; Galope, R.; Bajpai, V.K.; Lim, J.; Paek, W.K.; Park, Y.H. Diversity of Marine Bacteria and Their Bacteriocins: Applications in Aquaculture. Fish. Sci. Aquac. 2017, 25, 257–269. [Google Scholar] [CrossRef]
- Lee, H.-T.; Lee, C.-C.; Yang, J.-R.; Lai, J.Z.C.; Chang, K.Y.A. Large-Scale Structural Classification of Antimicrobial Peptides. BioMed. Res. Int. 2015, 2015, 475062. [Google Scholar] [CrossRef]
- Jenssen, H.; Hamill, P.; Hancock, R.E.W. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.T.; Haney, E.F.; Vogel, H.J. The expanding scope of antimicrobial peptide structures and their modes of action. Trends Biotechnol. 2011, 29, 464–472. [Google Scholar] [CrossRef]
- Fjell, C.D.J.; Hiss, A.; Hancock, R.E.W.; Schneider, G. Designing antimicrobial peptides: Form follows function. Nat. Rev. Drug Discov. 2012, 11, 37–51. [Google Scholar] [CrossRef]
- Boone, K.; Camarda, K.; Spencer, P.; Tamerler, C. Antimicrobial peptide similarity and classification through rough set theory using physicochemical boundaries. BMC Bioinform. 2018, 19, 469–473. [Google Scholar] [CrossRef]
- Ahmadi, S.; Ghollasi, M.; Hosseini, H.M. The apoptotic impact of nisin as a potent bacteriocin on the colon cancer cells. Microb. Pathog. 2017, 111, 193–197. [Google Scholar] [CrossRef]
- Klaessig, F.; Marrapese, M.; Abe, S. Current Perspectives in Nanotechnology Terminology and Nomenclature. In Nanotechnology Standards; Murashov, V., Howard, J., Eds.; Springer: New York, NY, USA, 2011; pp. 21–52. [Google Scholar]
- Chou, L.Y.T.; Ming, K.; Chan, W.C.W. Strategies for the intracellular delivery of nanoparticles. Chem. Soc. Rev. 2011, 40, 233–245. [Google Scholar] [CrossRef]
- Farokhzad, O.C.; Langer, R. Impact of nanotechnology on drug delivery. ASC Nano 2009, 3, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Brandelli, A. Nanostructures as promising tools for delivery of antimicrobial peptides. Mini Rev. Med. Chem. 2012, 12, 731–741. [Google Scholar] [CrossRef] [PubMed]
- Arthur, T.D.; Cavera, V.L.; Chikindas, M.L. On bacteriocin delivery systems and potential applications. Future Microbiol. 2014, 9, 235–248. [Google Scholar] [CrossRef]
- Mossallam, S.F.; Amer, E.I.; Diab, R.G. Potentiated anti-microsporidial activity of Lactobacillus acidophilus CH1 bacteriocin using gold nanoparticles. Exp. Parasitol. 2014, 144, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Zacharof, M.-P.; Coss, G.M.; Mandale, S.J.; Lovitt, R.W. Separation of lactobacilli bacteriocins from fermented broths using membranes. Process. Biochem. 2013, 48, 1252–1261. [Google Scholar] [CrossRef]
- Dicks, L.M.T.; van Staden, A.D.P.; Klumperman, B. Bacteriocins and Nanotechnology. In Micro and Nano Technologies; Elsevier: Amsterdam, The Netherlands, 2017; Volume 13, pp. 267–288. [Google Scholar]

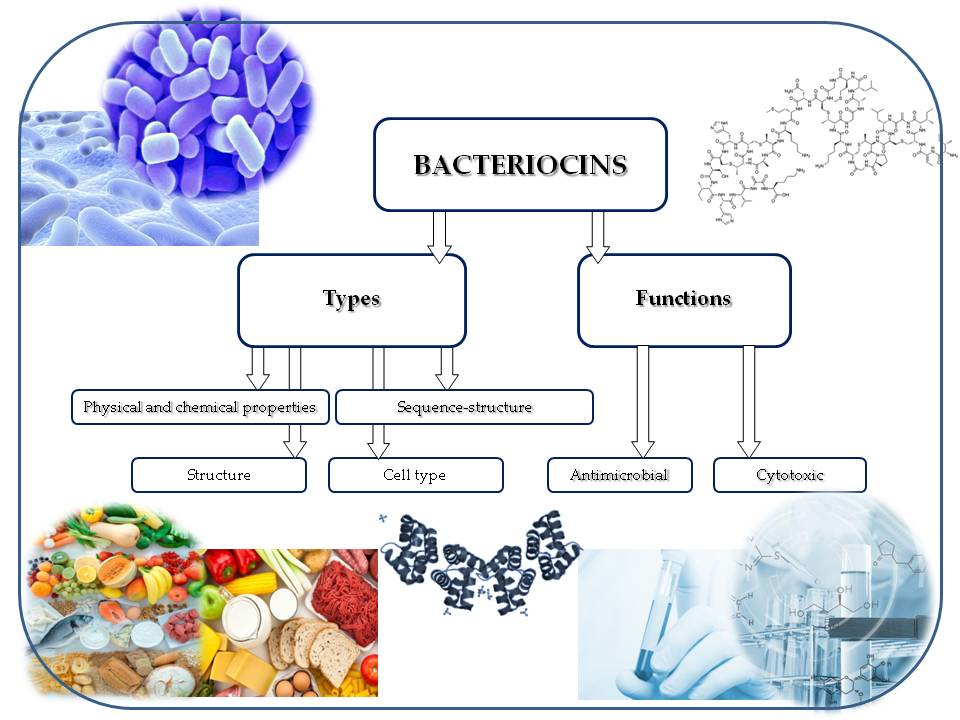{kind=link}
| Bacteria Name | Class | Size (kDa) | Examples | Source |
|---|---|---|---|---|
| Gram-negative bacteria | Colicins | 30–80 | Colicins A, B, E2, E3 | [17,18,19] |
| Colicin-like bacteriocins | 30–80 | S-piocins, klebicins | [17,19,20] | |
| Bacteriocins, phage-tail like | 20–100 | R- and F-piocins | [17,19,21] | |
| Microcins | <10 | Microcin C7, microcin B17, colicin V | [17,19,22] | |
| Gram-positive bacteria | Class I | <5 | Nisin, mersadicin, lacticin 3147 | [19,23] |
| Class II | <10 | Pediocin RA1, carnobacteriocin B2 | [19,24] | |
| Class II | >10 | Helvecin, enterocin AS-48 | [19,25,26] | |
| Archaea | Halocins | >5 | Halocin A4, C8, H1, H4 | [19,27] |
| Sulfolobaceae | ~20 | Sulfolobaceae | [19,28] |
| Class | Group | Distinctive Characteristic | Source of Information |
|---|---|---|---|
| Class I | Lantibiotics | Residues (methyl)lanthionine | [17,38,39] |
| Lipolantins | N-terminal fatty acid and avionin fragment | [26,27,29] | |
| Thiopeptides | 6-membered nitrogen heterocycle, azole rings | [26,28,40] | |
| Botromycins | Macrocyclic amidine, decarboxylated C-terminal thiazole, β-methylated residues | [26,28,39,41] | |
| Linear azole-containing peptides | thiazole and (methyl)oxazole rings, linear back bone | [19,42,43] | |
| Sactibiotics (sactipeptides) | Disulfide α-carbon bridges | [19,44,45,46,47] | |
| Lasso peptides | Cyclization of an N-terminal amine into a γ-acid | [17,27,40] | |
| Cyclic bacteriocins with a “head-to-tail” connection | Cycling from N-terminus to C-terminus | [26,27,48] | |
| Glycocins | Glycosylated residues | [17,37] | |
| Class II | YGNG-motif containing bacteriocins | Consensus YGNG-motif, at least one disulfide bridge | [19,48,49] |
| Linear two-peptide bacteriocins | Synergy of two peptides | [19,28,49] | |
| Leaderless bacteriocins | Lack of a leader peptide | [26,43,50] | |
| Other linear bacteriocins | Non-YGNG-like linear peptides | [19,42,51] | |
| Class III | Bacteriolysins | Large lytic polypeptides, | [27,52,53] |
| Non-lytic bacteriocins | Large non-lytic polypeptides | [41,49,52] | |
| Tailocins | Multiprotein complex, a structure similar to a phage tail | [21,53,54] |
| Target | Bacteriocin | Group | Source of Information |
|---|---|---|---|
| Lipid II | Nisin, microbisporicin, bovicin HJ50, mersacidin, lacticin 3147, haloduracin | Lantibiotics | [52,55] |
| Lactococcin 972 | Other linear bacteriocins | [52,56] | |
| Phosphatidylethanolamine | Cinnamycin | Lantibiotics | [52,57] |
| 50S ribosomal subunit | Thiostrepton, nosiheptide, micopoccine | Thiopeptides | [52,58] |
| Botromycin A2 | Botromycins | [52,56] | |
| Extension factor TU | Thiopeptide GE2270A | Thiopeptides | [52,56,58] |
| Regulator of cellular response WalR | Streptomonomicin | Lasso peptides | [57,59] |
| ClpC1 ATPase | Lassomycin | Lasso peptides | [35,60] |
| Glucose-phosphotransferase system | Sublancin 168, glycocin F | Glycocins | [35,60] |
| MscL mechanosensitive channel | Sublancin 168 | Glycocins | [55,61] |
| Maltose ABC-transporter | Garvicin ML | Cyclic bacteriocins with a “head-to-tail” connection | [52,60,61] |
| Mannose-phosphotransferase system | Pediocin PA-1, leucocin A, carnobacteriocin B2, sacacin P, curvacin A, enterocin HF | YGNG-motif containing bacteriocins | [56,60] |
| Lactococcin A, garviacin Q | Other linear bacteriocins | [58,60,61] | |
| Undecaprenyl-pyrophosphate-phosphatase | Lactococcin G, enterocin 1071 | Two-peptide bacteriocins | [39,58] |
| Amino acid-polyamine-organocation transporter | Plantaricin JK | Two-peptide bacteriocins | [35,39,55] |
| Zn-dependent membrane-bound metallopeptidase | LsbB, enterocin K1 | Leaderless bacteriocins | [58,62] |
| Peptidoglycan | Lysostaphin, zoocin A, millericin B, enterolisin A | Bacteriolysins | [52,56,58] |
| Lipopolysaccharides | Diffocin, monocin | Tailocins | [35,39,52,56] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zimina, M.; Babich, O.; Prosekov, A.; Sukhikh, S.; Ivanova, S.; Shevchenko, M.; Noskova, S. Overview of Global Trends in Classification, Methods of Preparation and Application of Bacteriocins. Antibiotics 2020, 9, 553. https://doi.org/10.3390/antibiotics9090553
Zimina M, Babich O, Prosekov A, Sukhikh S, Ivanova S, Shevchenko M, Noskova S. Overview of Global Trends in Classification, Methods of Preparation and Application of Bacteriocins. Antibiotics. 2020; 9(9):553. https://doi.org/10.3390/antibiotics9090553
Chicago/Turabian StyleZimina, Maria, Olga Babich, Alexander Prosekov, Stanislav Sukhikh, Svetlana Ivanova, Margarita Shevchenko, and Svetlana Noskova. 2020. "Overview of Global Trends in Classification, Methods of Preparation and Application of Bacteriocins" Antibiotics 9, no. 9: 553. https://doi.org/10.3390/antibiotics9090553
APA StyleZimina, M., Babich, O., Prosekov, A., Sukhikh, S., Ivanova, S., Shevchenko, M., & Noskova, S. (2020). Overview of Global Trends in Classification, Methods of Preparation and Application of Bacteriocins. Antibiotics, 9(9), 553. https://doi.org/10.3390/antibiotics9090553