Rapid Antibacterial Activity of Cannabichromenic Acid against Methicillin-Resistant Staphylococcus aureus

 ,
,  ,
, 
Abstract
1. Introduction
2. Results and Discussion
2.1. Phytocannabinoid Analogues Inhibit the Growth of Drug-Resistant and Pathogenic Bacteria
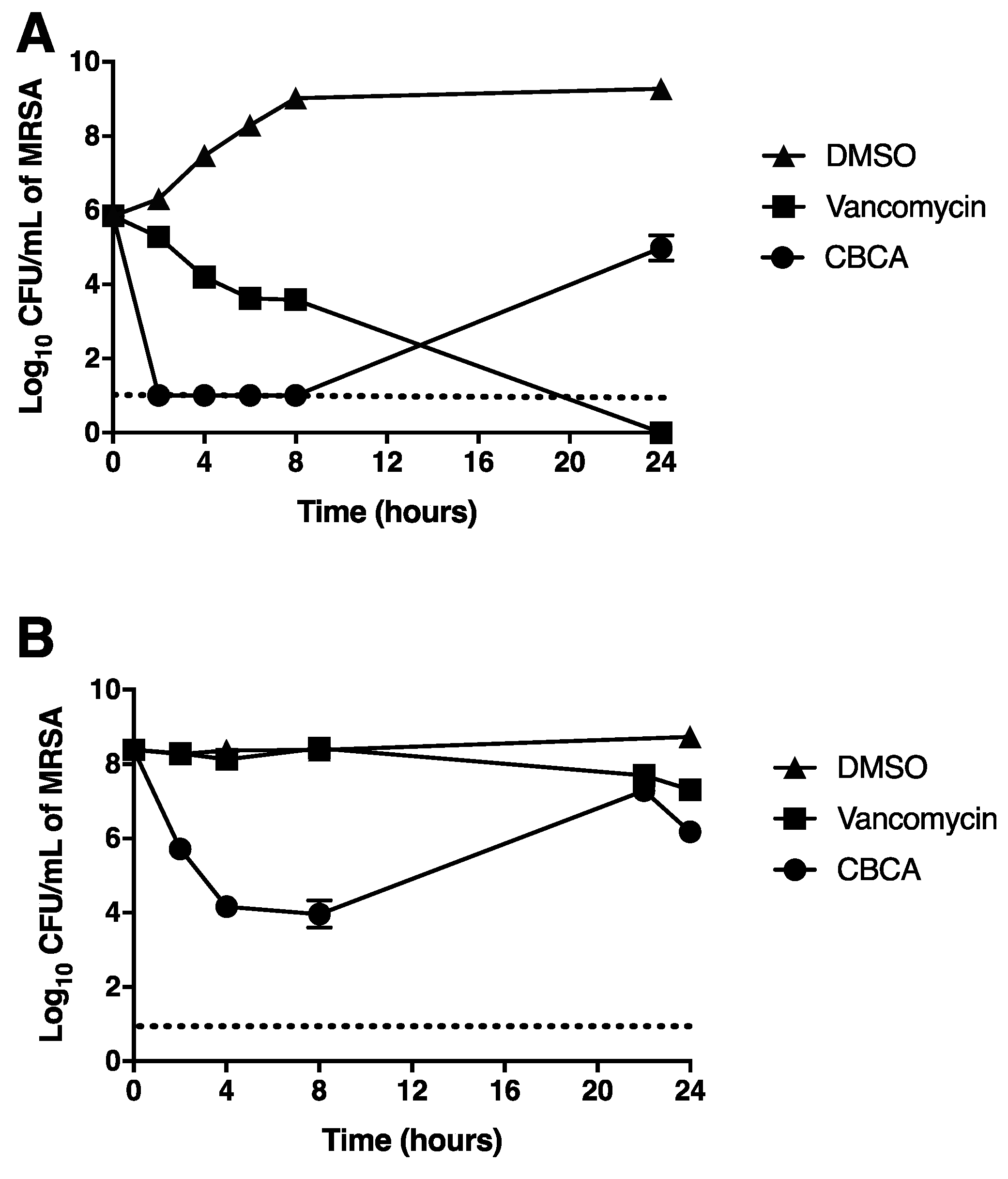
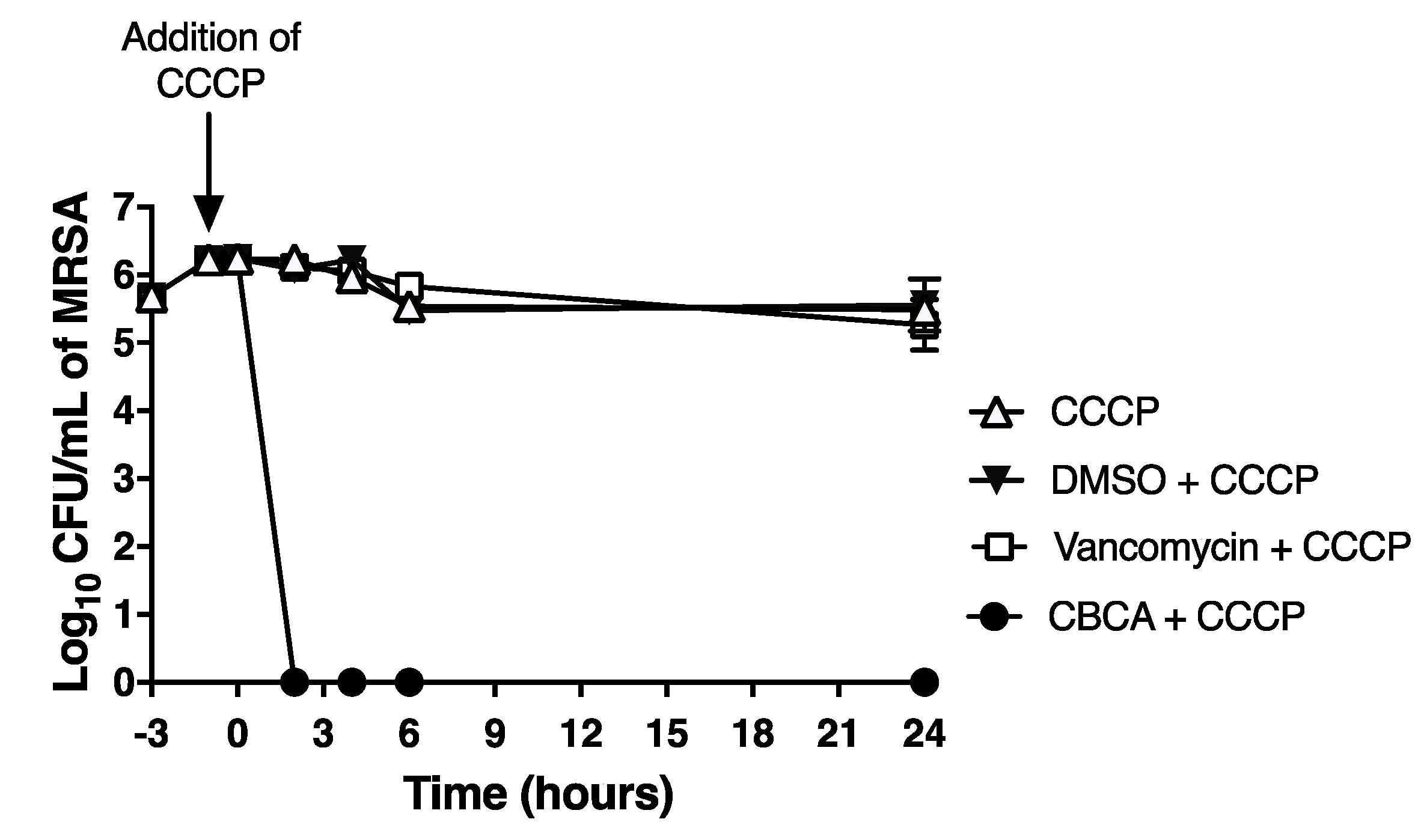
2.2. CBCA Exerts Rapid Bactericidal Activity that Is Independent of Bacterial Cell Density and Metabolism
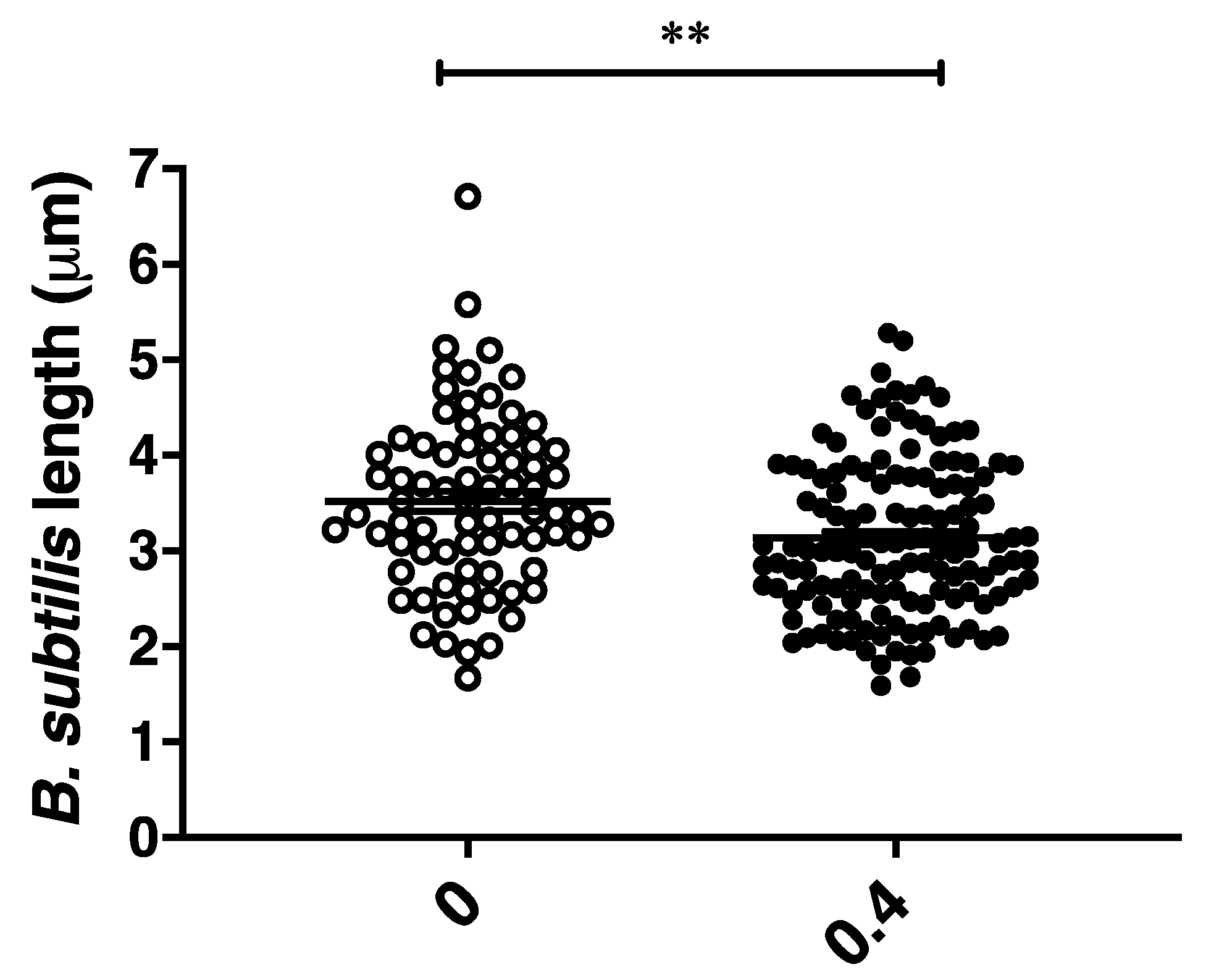
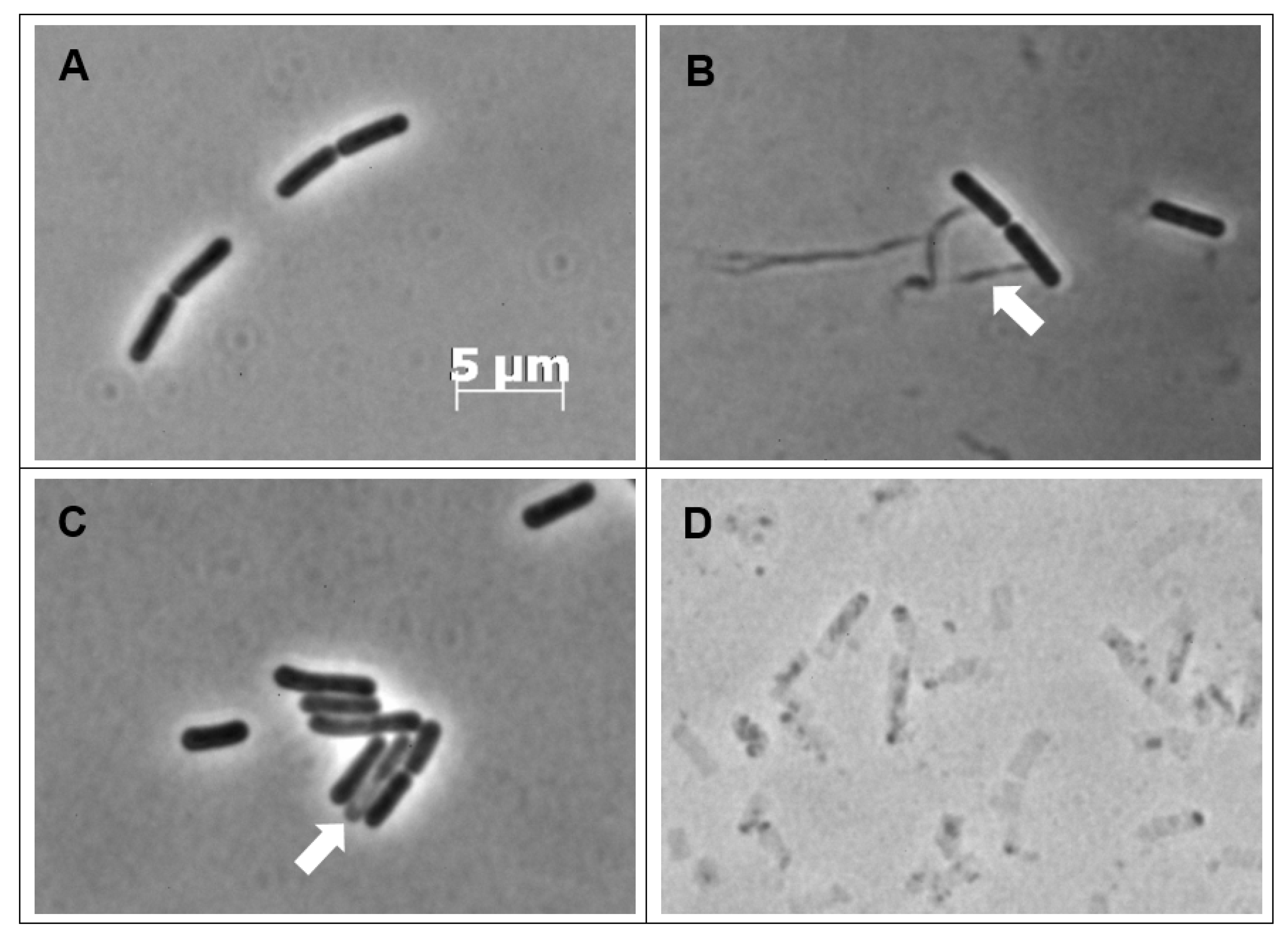
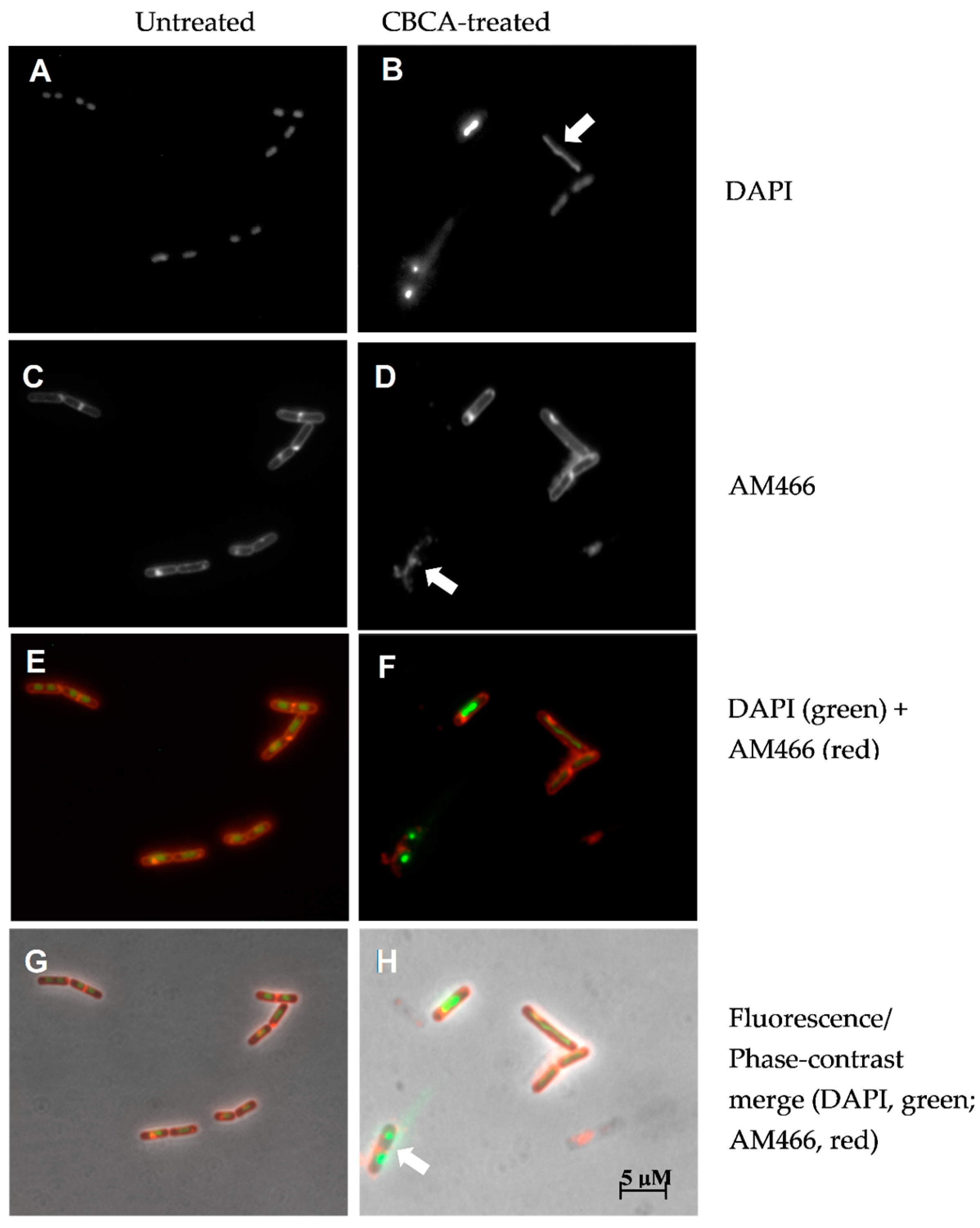
2.3. CBCA Induces Distinct and Degradative Morphological Changes Indicative of Bacterial Degeneration and Cell Lysis
3. Materials and Methods
3.1. Bacterial Strains
3.2. Cannabinoid Compounds
3.3. Bacterial Growth Conditions
3.4. Mammalian Cell Growth Conditions
3.5. Resazurin Reduction Assay for Minimum Inhibitory Concentration
3.6. Resazurin Reduction Assay for Minimum Toxicity Concentration
3.7. Time-Kill Analysis of Compound Inhibition
3.8. Phase-Contrast and Fluorescence Microscopy
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rhee, Y.; Aroutcheva, A.; Hota, B.; Weinstein, R.A.; Popovich, K.J. Evolving epidemiology of Staphylococcus aureus bacteremia. Infect. Control Hosp. Epidemiol. 2015, 36, 1417–1422. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.Y.; Singh, A.; David, M.Z.; Bartsch, S.M.; Slayton, R.B.; Huang, S.S.; Zimmer, S.M.; Potter, M.A.; Macal, C.M.; Lauderdale, D.S.; et al. The economic burden of community-associated methicillin-resistant Staphylococcus aureus (CA-MRSA). Clin. Microbiol. Infect. 2013, 19, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Kourtis, A.P.; Hatfield, K.; Baggs, J.; Mu, Y.; See, I.; Epson, E.; Nadle, J.; Kainer, M.A.; Dumyati, G.; Petit, S.; et al. Vital Signs: Epidemiology and Recent Trends in Methicillin-Resistant and in Methicillin-Susceptible Staphylococcus aureus Bloodstream Infections—United States. Morb. Mortal. Wkly. Rep. 2019, 68, 214. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.G.; Perdreau-Remington, F.; Rieg, G.; Mehdi, S.; Perlroth, J.; Bayer, A.S.; Tang, A.W.; Phung, T.O.; Spellberg, B. Necrotizing fasciitis caused by community-associated methicillin-resistant Staphylococcus aureus in Los Angeles. N. Eng. J. Med. 2005, 352, 1445–1453. [Google Scholar] [CrossRef] [PubMed]
- Gillet, Y.; Issartel, B.; Vanhems, P.; Fournet, J.-C.; Lina, G.; Bes, M.; Vandenesch, F.; Piémont, Y.; Brousse, N.; Floret, D.; et al. Association between Staphylococcus aureus strains carrying gene for Panton-Valentine leukocidin and highly lethal necrotising pneumonia in young immunocompetent patients. Lancet 2002, 359, 753–759. [Google Scholar] [CrossRef]
- Chambers, H.F.; DeLeo, F.R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. Microbiol. 2009, 7, 629. [Google Scholar] [CrossRef]
- Kaur, D.C.; Chate, S.S. Study of antibiotic resistance pattern in methicillin resistant Staphylococcus aureus with special reference to newer antibiotic. J. Glob. Infect. Dis. 2015, 7, 78. [Google Scholar] [CrossRef]
- VanEperen, A.S.; Segreti, J. Empirical therapy in methicillin-resistant Staphylococcus aureus infections: An up-to-date approach. J. Infect. Chemother. 2016, 22, 351–359. [Google Scholar] [CrossRef]
- Weigel, L.M.; Chow, J.; Kopp, R.J.; Portney, P.R. Genetic analysis of a high-level vancomycin-resistant isolate of Staphylococcus aureus. Science 2003, 302, 1569–1571. [Google Scholar] [CrossRef]
- Butler, M.S.; Blaskovich, M.; Cooper, M.A. Antibiotics in the clinical pipeline at the end of 2015. J. Antibiot. 2017, 70, 3. [Google Scholar] [CrossRef]
- Chien, J.W.; Kucia, M.L.; Salata, R.A. Use of linezolid, an oxazolidinone, in the treatment of multidrug-resistant gram-positive bacterial infections. Clin. Infect. Dis. 2000, 30, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Tally, F.P.; Zeckel, M.; Wasilewski, M.M.; Carini, C.; Berman, C.L.; Drusano, G.L.; Oleson, F.B., Jr. Daptomycin: A novel agent for Gram-positive infections. Exp. Opin. Investig. Drugs 1999, 8, 1223–1238. [Google Scholar] [CrossRef] [PubMed]
- Flamm, R.K.; Mendes, R.E.; Hogan, P.A.; Streit, J.M.; Ross, J.E.; Jones, R.N. Linezolid surveillance results for the United States (LEADER surveillance program 2014). Antimicrob. Agents Chemother. 2016, 60, 2273–2280. [Google Scholar] [CrossRef] [PubMed]
- Reekie, T.A.; Scott, M.P.; Kassiou, M. The evolving science of phytocannabinoids. Nat. Rev. Chem. 2018, 2, 0101. [Google Scholar] [CrossRef]
- Reddy, P.M.; Maurya, N.; Velmurugan, B.K. Medicinal Use of Synthetic Cannabinoids—A Mini Review. Curr. Pharmacol. Rep. 2019, 5, 1–13. [Google Scholar] [CrossRef]
- Kumar, K.K.; Shalev-Benami, M.; Robertson, M.J.; Hu, H.; Banister, S.; Hollingsworth, S.A.; Latorraca, N.R.; Kato, H.E.; Hilger, D.; Maeda, S.; et al. Structure of a signaling cannabinoid receptor 1-G protein complex. Cell 2019, 176, 448–458. [Google Scholar] [CrossRef]
- Klahn, P. Cannabinoids-Promising Antimicrobial Drugs orIntoxicants with Benefits? Antibiotics 2020, 9, 297. [Google Scholar] [CrossRef]
- Farha, M.A.; El-Halfawy, O.M.; Gale, R.T.; Macnair, C.R.; Carfrae, L.A.; Zhang, X.; Jentsch, N.G.; Magolan, J.; Brown, E.D. Uncovering the Hidden Antibiotic Potential of Cannabis. ACS Infect. Dis. 2020, 6, 338–346. [Google Scholar] [CrossRef]
- Reekie, T.; Scott, M.; Kassiou, M. Synthesis of Phytocannabinoids Including a Decarboxylation Step. U.S. Patent Application WO/2019/033168, 4 June 2020. [Google Scholar]
- Reekie, T.; Scott, M.; Kassiou, M. Synthesis of Phytocannabinoids Including a Demethylation Step. U.S. Patent Application WO/2019/033164, 4 June 2020. [Google Scholar]
- Silver, L.L.; Bostian, K. Screening of natural products for antimicrobial agents. Eur. J. Clin. Microbiol. Infect. Dis. 1990, 9, 455–461. [Google Scholar] [CrossRef]
- Udekwu, K.I.; Parrish, N.F.; Ankomah, P.; Baquero, F.; Levin, B.R. Functional relationship between bacterial cell density and the efficacy of antibiotics. J. Antimicrob. Chemother. 2009, 63, 745–757. [Google Scholar] [CrossRef]
- Mascio, C.T.M.; Alder, J.D.; Silverman, J.A. Bactericidal action of daptomycin against stationary-phase and nondividing Staphylococcus aureus cells. Antimicrob. Agents Chemother. 2007, 51, 4255–4260. [Google Scholar] [CrossRef] [PubMed]
- Richter, K.; Thomas, N.; Zhang, G.; Prestidge, C.A.; Coenye, T.; Wormald, P.-J.; Vreugde, S. Deferiprone and gallium-protoporphyrin have the capacity to potentiate the activity of antibiotics in Staphylococcus aureus small colony variants. Front. Cell. Infect. Microbial. 2017, 7, 280. [Google Scholar] [CrossRef] [PubMed]
- Ji, G.; Beavis, R.C.; Novick, R.P. Cell density control of staphylococcal virulence mediated by an octapeptide pheromone. Proc. Natl. Acad. Sci. USA 1995, 92, 12055–12059. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.L.; Schneider, T.; Peoples, A.J.; Spoering, A.L.; Engels, I.; Conlon, B.P.; Müller, A.; Schäberle, T.F.; Hughes, D.E.; Epstein, S.; et al. A new antibiotic kills pathogens without detectable resistance. Nature 2015, 517, 455. [Google Scholar] [CrossRef]
- Walsh, C. Molecular mechanisms that confer antibacterial drug resistance. Nature 2000, 406, 775. [Google Scholar] [CrossRef]
- Germain, E.; Roghanian, M.; Gerdes, K.; Maisonneuve, E. Stochastic induction of persister cells by HipA through (p) ppGpp-mediated activation of mRNA endonucleases. Proc. Natl. Acad. Sci. USA 2015, 112, 5171–5176. [Google Scholar] [CrossRef]
- Fischer-Parton, S.; Parton, R.M.; Hickey, P.C.; Dijksterhuis, J.; Atkinson, H.A.; Read, N.D. Confocal microscopy of FM4-64 as a tool for analysing endocytosis and vesicle trafficking in living fungal hyphae. J. Microsc. 2000, 198, 246–259. [Google Scholar] [CrossRef]
- Porter, K.G.; Feig, Y.S. The use of DAPI for identifying and counting aquatic microflora 1. Limnol. Oceanogr. 1980, 25, 943–948. [Google Scholar] [CrossRef]
- Galvez, A.; Maqueda, M.; Martínez-Bueno, M.; Valdivia, E. Permeation of bacterial cells, permeation of cytoplasmic and artificial membrane vesicles, and channel formation on lipid bilayers by peptide antibiotic AS-48. J. Bacteriol. 1991, 173, 886–892. [Google Scholar] [CrossRef]
- Hancock, R.E. Peptide antibiotics. Antimicrob. Agents Chemother. 1999, 43, 1317–1323. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Burkholder, P.R.; Giles, N.H., Jr. Induced biochemical mutations in Bacillus subtilis. Am. J. Bot. 1947, 34, 345–348. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibacterial Activity (MIC; μM) a | Cytotoxicity (MTC; µM) b | ||||
|---|---|---|---|---|---|
| Compound | S. aureus (MRSA) | S. aureus (MSSA) | E. faecalis (VRE) | A549 | HepG2 |
 (±)-CBCA | 3.9 | 7.8 | 7.8 | 250 | 125 |
 (±)-CBCTFA | >250 | >250 | >250 | n.d. | n.d. |
 (±)-CBLM | >250 | >250 | >250 | n.d. | n.d. |
 (±)-CBCM | >250 | >250 | >250 | n.d. | n.d. |
 CBDVM | 15.6 | >250 | >250 | 125 | 250 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galletta, M.; Reekie, T.A.; Nagalingam, G.; Bottomley, A.L.; Harry, E.J.; Kassiou, M.; Triccas, J.A. Rapid Antibacterial Activity of Cannabichromenic Acid against Methicillin-Resistant Staphylococcus aureus. Antibiotics 2020, 9, 523. https://doi.org/10.3390/antibiotics9080523
Galletta M, Reekie TA, Nagalingam G, Bottomley AL, Harry EJ, Kassiou M, Triccas JA. Rapid Antibacterial Activity of Cannabichromenic Acid against Methicillin-Resistant Staphylococcus aureus. Antibiotics. 2020; 9(8):523. https://doi.org/10.3390/antibiotics9080523
Chicago/Turabian StyleGalletta, Maria, Tristan A. Reekie, Gayathri Nagalingam, Amy L. Bottomley, Elizabeth J. Harry, Michael Kassiou, and James A. Triccas. 2020. "Rapid Antibacterial Activity of Cannabichromenic Acid against Methicillin-Resistant Staphylococcus aureus" Antibiotics 9, no. 8: 523. https://doi.org/10.3390/antibiotics9080523
APA StyleGalletta, M., Reekie, T. A., Nagalingam, G., Bottomley, A. L., Harry, E. J., Kassiou, M., & Triccas, J. A. (2020). Rapid Antibacterial Activity of Cannabichromenic Acid against Methicillin-Resistant Staphylococcus aureus. Antibiotics, 9(8), 523. https://doi.org/10.3390/antibiotics9080523