Efficacy and Gut Dysbiosis of Gentamicin-Intercalated Smectite as a New Therapeutic Agent against Helicobacter pylori in a Mouse Model

Abstract
1. Introduction
2. Results
2.1. CLO test and PCR Assay of H. pylori in Gastric Mucosa
2.2. Pro-Inflammatory Cytokines and Atrophy of Gastric Mucosa
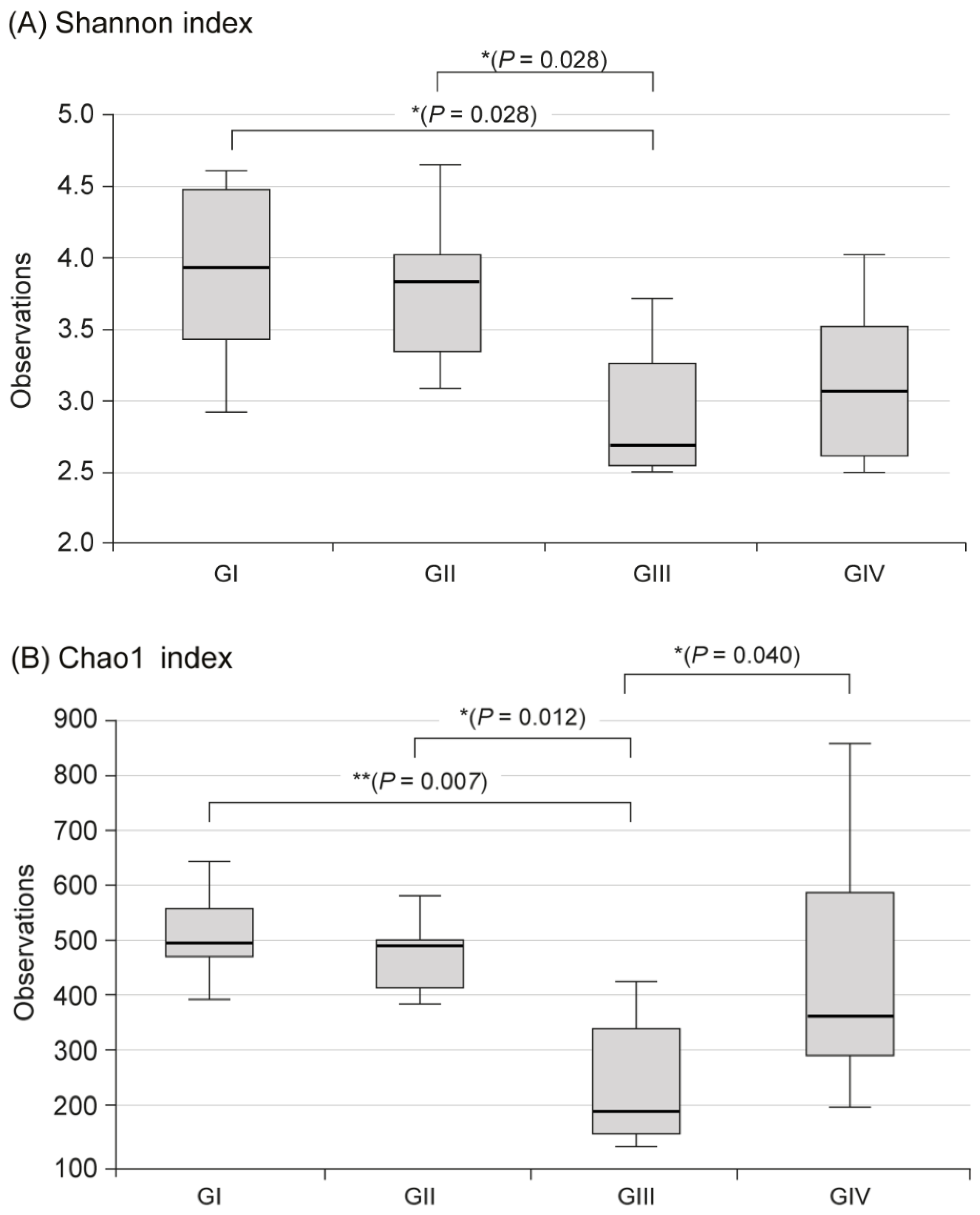
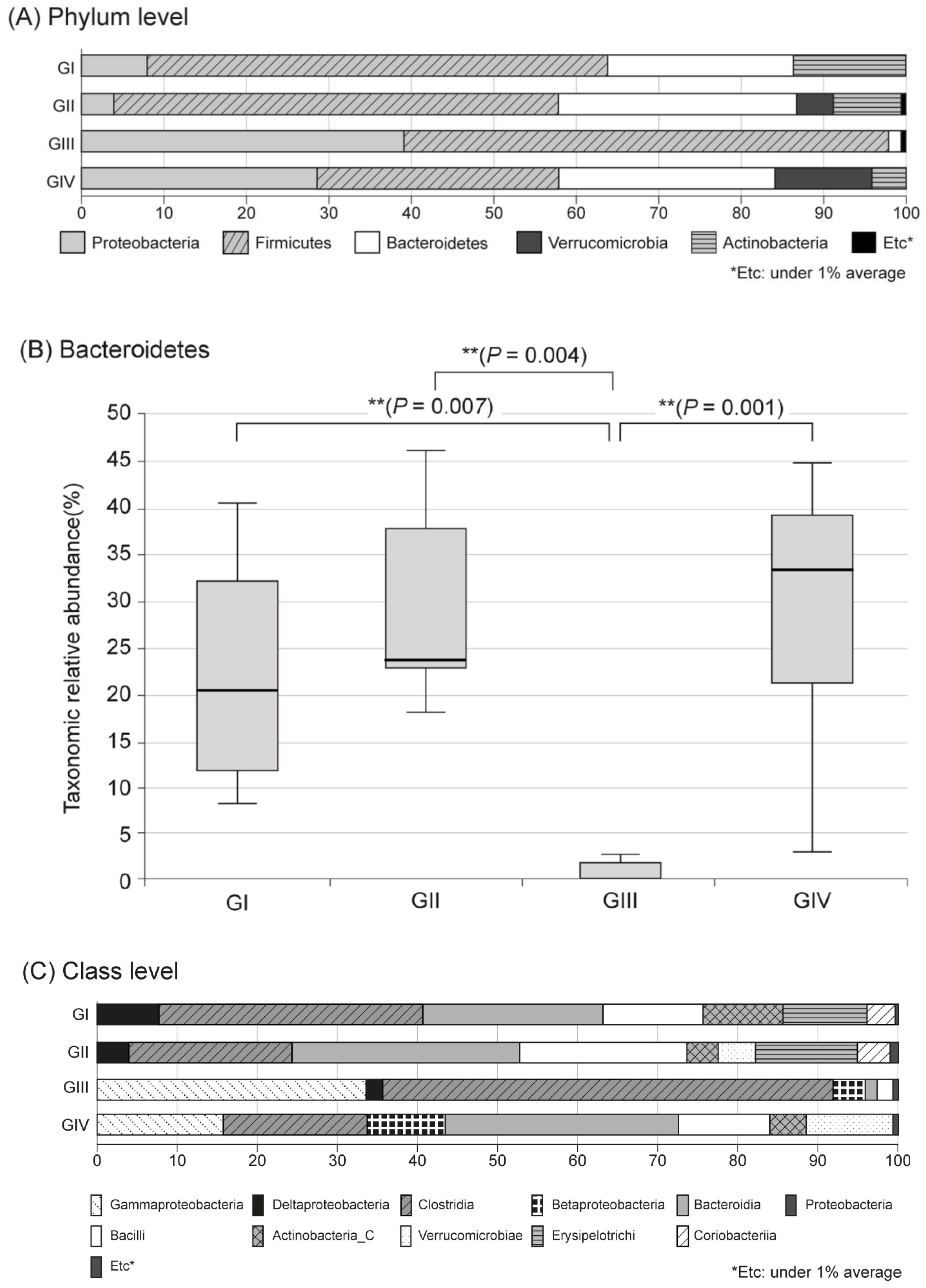
2.3. Changes in Fecal Microbiota
3. Discussion
4. Materials and Methods
4.1. Intercalation of GM
4.2. Animal Preparation
4.3. Anti-H. pylori Efficacy In Vivo
4.3.1. H. pylori Strains and Culture Conditions
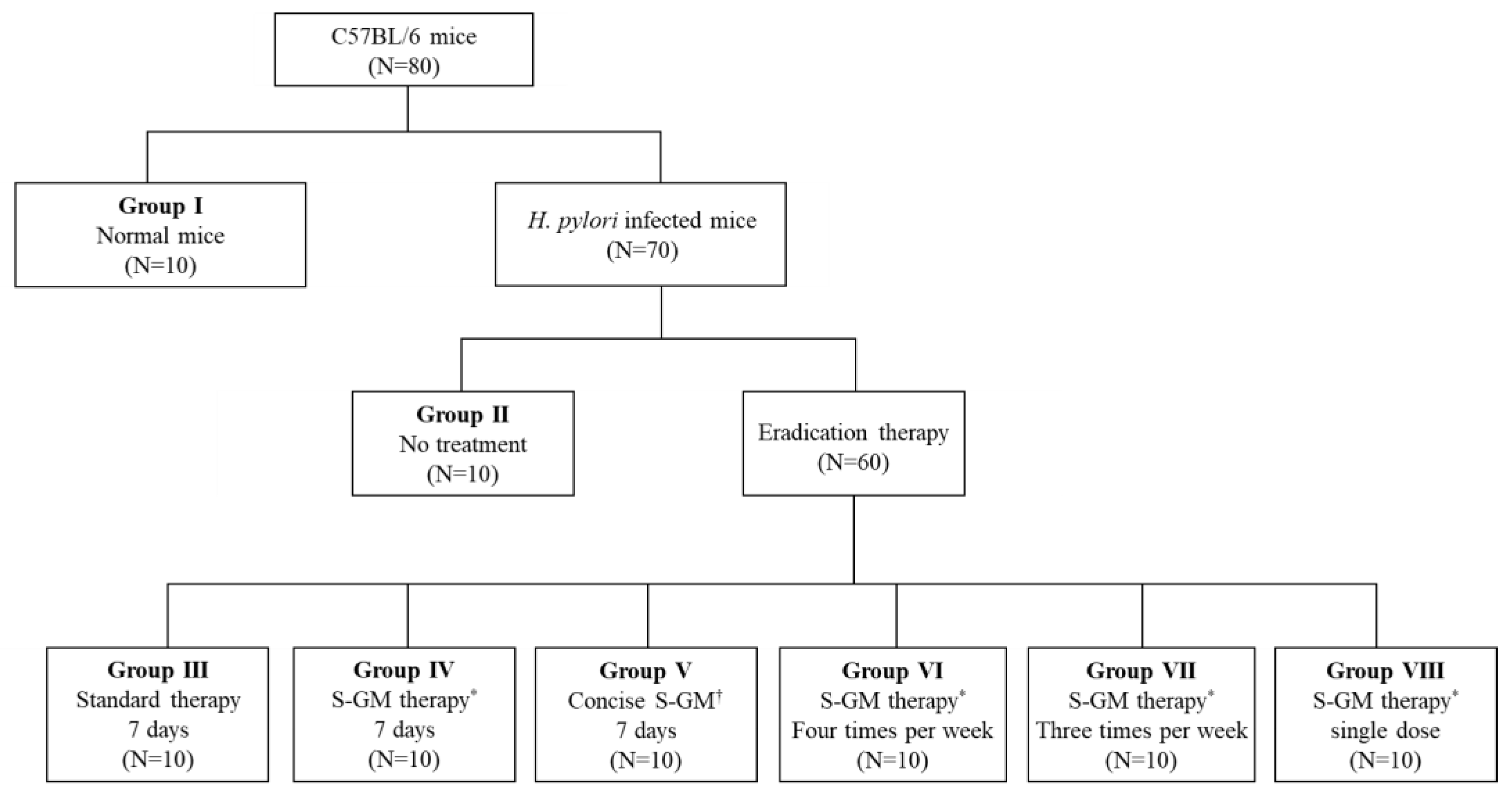
4.3.2. Inoculation of Experimental Animals
4.3.3. Distribution of Animals
4.3.4. CLO Test and PCR Assay of H. pylori in Gastric Mucosa
4.3.5. Pro-Inflammatory Cytokines and Atrophy of Gastric Mucosa
4.4. Fecal Microbiota
4.4.1. DNA Extraction from Fecal Materials
4.4.2. PCR Amplification and 16S rRNA Gene Sequencing
4.4.3. Bioinformatics for Microbiota Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| CLO | Campylobacter-like organism test |
| DNA | Deoxyribonucleic acid |
| ELISA | Enzyme-linked immunosorbent assay |
| GM | Gentamicin |
| H. pylori | Helicobacter pylori |
| IgG | Immunoglobulin G |
| IL-6 | Interleukin-6 |
| OUT | Operational taxonomic units |
| PCoA | Principal coordinates analysis |
| PCR | Polymerase chain reaction |
| PD | Pharmacodynamics |
| PK | Pharmacokinetics |
| PPI | Proton pump inhibitor |
| RNA | Ribonucleic acid |
| S-GM | Gentamicin-intercalated smectite hybrid |
| TNF-α | Tumor necrosis factor-α |
References
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Kusters, J.G.; van Vliet, A.H.; Kuipers, E.J. Pathogenesis of Helicobacter pylori infection. Clin. Microbiol. Rev. 2006, 19, 449–490. [Google Scholar] [CrossRef] [PubMed]
- Malfertheiner, P.; Megraud, F.; O’Morain, C.A.; Gisbert, J.P.; Kuipers, E.J.; Axon, A.T.; Bazzoli, F.; Gasbarrini, A.; Atherton, J.; Graham, D.Y.; et al. Management of Helicobacter pylori infection-the maastricht v/florence consensus report. Gut 2017, 66, 6–30. [Google Scholar] [CrossRef] [PubMed]
- O’Morain, N.R.; Dore, M.P.; O’Connor, A.J.; Gisbert, J.P.; O’Morain, C.A. Treatment of h elicobacter pylori infection in 2018. Helicobacter 2018, 23, e12519. [Google Scholar]
- Kaakoush, N.O.; Asencio, C.; Mégraud, F.; Mendz, G.L. A redox basis for metronidazole resistance in Helicobacter pylori. Antimicrob. Agents Chemother. 2009, 53, 1884–1891. [Google Scholar] [CrossRef]
- Song, Z.; Zhang, J.; He, L.; Chen, M.; Hou, X.; Li, Z.; Zhou, L. Prospective multi-region study on primary antibiotic resistance of Helicobacter pylori strains isolated from chinese patients. Dig. Liver Dis. 2014, 46, 1077–1081. [Google Scholar] [CrossRef]
- Lee, K.H.; Park, S.Y.; Jeong, S.J.; Jung, D.H.; Kim, J.-H.; Jeong, S.H.; Kang, I.-M.; Song, Y.G. Can aminoglycosides be used as a new treatment for Helicobacter pylori? In Vitro activity of recently isolated Helicobacter pylori. Infect. Chemother. 2019, 51, 10–20. [Google Scholar] [CrossRef]
- Cox, C.E. Gentamicin. Med. Clin. N. Am. 1970, 54, 1305–1315. [Google Scholar] [CrossRef]
- Recchia, J.; Lurantos, M.H.; Amsden, J.A.; Storey, J.; Kensil, C.R. A semisynthetic quillaja saponin as a drug delivery agent for aminoglycoside antibiotics. Pharm. Res. 1995, 12, 1917–1923. [Google Scholar] [CrossRef]
- Thomas, F.; Michot, L.J.; Vantelon, D.; Montarges, E.; Prelot, B.; Cruchaudet, M.; Delon, J.-F. Layer charge and electrophoretic mobility of smectites. Colloids Surf. A Physicochem. Eng. Asp. 1999, 159, 351–358. [Google Scholar] [CrossRef]
- Jeong, S.J.; Kim, J.; Jung, D.H.; Lee, K.H.; Park, S.Y.; Kang, I.; Song, Y.G. Gentamicin-intercalated smectite as a new therapeutic option for Helicobacter pylori eradication. J. Antimicrob. Chemother. 2018, 73, 1324–1329. [Google Scholar]
- Nicholson, J.K.; Holmes, E.; Kinross, J.; Burcelin, R.; Gibson, G.; Jia, W.; Pettersson, S. Host-gut microbiota metabolic interactions. Science 2012, 336, 1262–1267. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.L.; Vieira Silva, S.; Liston, A.; Raes, J. How informative is the mouse for human gut microbiota research? Dis. Models Mech. 2015, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Moodley, Y.; Linz, B.; Bond, R.P.; Nieuwoudt, M.; Soodyall, H.; Schlebusch, C.M.; Bernhöft, S.; Hale, J.; Suerbaum, S.; Mugisha, L.; et al. Age of the association between Helicobacter pylori and man. PLoS Pathog. 2012, 8, e1002693. [Google Scholar] [CrossRef] [PubMed]
- Warren, J.R.; Marshall, B. Unidentified curved bacilli on gastric epithelium in active chronic gastritis. Lancet 1983, 1, 1273–1275. [Google Scholar] [PubMed]
- Miftahussurur, M.; Syam, A.F.; Nusi, I.A.; Makmun, D.; Waskito, L.A.; Zein, L.H.; Akil, F.; Uwan, W.B.; Simanjuntak, D.; Wibawa, I.D.N.; et al. Surveillance of Helicobacter pylori antibiotic susceptibility in indonesia: Different resistance types among regions and with novel genetic mutations. PLoS ONE 2016, 11, e0166199. [Google Scholar] [CrossRef]
- Ontsira Ngoyi, E.N.; Atipo Ibara, B.I.; Moyen, R.; Ahoui Apendi, P.C.; Ibara, J.R.; Obengui, O.; Ossibi Ibara, R.B.; Nguimbi, E.; Niama, R.F.; Ouamba, J.M.; et al. Molecular detection of Helicobacter pylori and its antimicrobial resistance in brazzaville, congo. Helicobacter 2015, 20, 316–320. [Google Scholar] [CrossRef]
- De Bortoli, N.; Leonardi, G.; Ciancia, E.; Merlo, A.; Bellini, M.; Costa, F.; Mumolo, M.G.; Ricchiuti, A.; Cristiani, F.; Santi, S.; et al. Helicobacter pylori eradication: A randomized prospective study of triple therapy versus triple therapy plus lactoferrin and probiotics. Am. J. Gastroenterol. 2007, 102, 951–956. [Google Scholar] [CrossRef]
- Yang, J.; Lin, C.; Wang, H.; Chen, J.; Kao, J.Y.; Shun, C.; Lu, C.; Lin, B.; Shieh, M.; Chang, M.; et al. High-dose dual therapy is superior to standard first-line or rescue therapy for Helicobacter pylori infection. Clin. Gastroenterol. Hepatol. 2015, 13, 895–905. [Google Scholar] [CrossRef]
- Oh, B.; Kim, J.S.; Koh, S.; Kim, B.G.; Lee, K.L.; Chun, J. The effect of probiotics on gut microbiota during the Helicobacter pylori eradication: Randomized controlled trial. Helicobacter 2016, 21, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Tønnesen, H.H.; Karlsen, J. Alginate in drug delivery systems. Drug Dev. Ind. Pharm. 2002, 28, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.M.; Pereira Marques, J.; Pinto Ribeiro, I.; Costa, J.L.; Carneiro, F.; Machado, J.C.; Figueiredo, C. Gastric microbial community profiling reveals a dysbiotic cancer-associated microbiota. Gut 2018, 67, 226–236. [Google Scholar] [CrossRef]
- Jo, H.J.; Kim, N.; Park, J.H.; Nam, R.H.; Seok, Y.; Kim, Y.; Kim, J.M.; Lee, D.H.; Jung, H.C. Analysis of gastric microbiota by pyrosequencing: Minor role of bacteria other than Helicobacter pylori in the gastric carcinogenesis. Helicobacter 2016, 21, 364–374. [Google Scholar] [CrossRef]
- Jakobsson, H.E.; Jernberg, C.; Andersson, A.F.; Sjölund Karlsson, M.; Jansson, J.K.; Engstrand, L. Short-term antibiotic treatment has differing long-term impacts on the human throat and gut microbiome. PLoS ONE 2010, 5, e9836. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Zou, J.; Zhang, Z.; Zhao, X.; Noriega, J.; Zhang, B.; Zhao, C.; Ingle, H.; Bittinger, K.; Mattei, L.M.; et al. Segmented filamentous bacteria prevent and cure rotavirus infection. Cell 2019, 179, 644–658. [Google Scholar] [CrossRef]
- Robertson, S.J.; Lemire, P.; Maughan, H.; Goethel, A.; Turpin, W.; Bedrani, L.; Guttman, D.S.; Croitoru, K.; Girardin, S.E.; Philpott, D.J. Comparison of co-housing and littermate methods for microbiota standardization in mouse models. Cell Rep. 2019, 27, 1910–1919. [Google Scholar] [CrossRef]
- Kim, B.-J.; Lee, S.-H.; Lyu, M.-A.; Kim, S.-J.; Bai, G.-H.; Kim, S.-J.; Chae, G.-T.; Kim, E.-C.; Cha, C.-Y.; Kook, Y.-H. Identification of mycobacterial species by comparative sequence analysis of the rna polymerase gene (rpob). J. Clin. Microbiol. 1999, 37, 1714–1720. [Google Scholar] [CrossRef]
- Kim, Y.B.; Kim, S.T.; Lee, S.W.; Jeen, Y.T.; Chun, H.J.; Lee, H.S.; Song, C.W.; Um, S.H.; Choi, J.H.; Kim, C.D. The influence of number of gastroscopic biopsy specimens on follow-up campylobacter-like organism (clo) test. Korean J. Gastroenterol. 2000, 35, 422–428. [Google Scholar]
- Kundu, P.; Mukhopadhyay, A.K.; Patra, R.; Banerjee, A.; Berg, D.E.; Swarnakar, S. Cag pathogenicity island-independent up-regulation of matrix metalloproteinases-9 and -2 secretion and expression in mice by Helicobacter pylori infection. J. Biol. Chem. 2006, 281, 34651–34662. [Google Scholar] [CrossRef]
- Fox, J.G.; Beck, P.; Dangler, C.A.; Whary, M.T.; Wang, T.C.; Shi, H.N.; Nagler Anderson, C. Concurrent enteric helminth infection modulates inflammation and gastric immune responses and reduces helicobacter-induced gastric atrophy. Nat. Med. 2000, 6, 536–542. [Google Scholar] [CrossRef]
- Layton, A.; McKay, L.; Williams, D.; Garrett, V.; Gentry, R.; Sayler, G. Development of bacteroides 16s rrna gene taqman-based real-time pcr assays for estimation of total, human, and bovine fecal pollution in water. Appl. Environ. Microbiol. 2006, 72, 4214–4224. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. Uchime improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Willis, A.D. Rarefaction, alpha diversity, and statistics. Front. Microbiol. 2019, 10, 2407. [Google Scholar] [CrossRef]
- Lin, J. Divergence measures based on the shannon entropy. IEEE Trans. Inf. Theory 1991, 37, 145–151. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Inoculation | Percentage of Animals with Positive a CLO Test Result, % | CLO Scores | Relative Fold Change in HP/GAPDH b | ||||
|---|---|---|---|---|---|---|---|---|
| HP Infection | Treatment | Duration of Treatment | Positive | Partially Positive | Negative | |||
| I | No | DW | D1–D7 | 0 | 0 | 100 | 0.0 ± 0.0 | 1.06 ± 0.39 |
| II | Yes | DW | D1–D7 | 80 | 20 | 0 | 2.8 ± 0.4 | 25,258.79 ± 6804.29 |
| III | Yes | AMX + CLR + PPI | D1–D7 | 0 | 10 | 90 | 0.1 ± 0.3 * | 4.23 ± 5.97* |
| IV | Yes | AMX + S-GM + PPI | D1–D7 | 0 | 10 | 90 | 0.1 ± 0.3 * | 3.74 ± 4.71 * |
| V | Yes | S-GM + PPI | D1–D7 | 0 | 20 | 80 | 0.2 ± 0.4 * | 522.90 ± 934.89 * |
| VI | Yes | AMX + S-GM + PPI | D1, D3, D5, D7 | 0 | 20 | 80 | 0.2 ± 0.4 * | 4.33 ± 4.07 * |
| VII | Yes | AMX + S-GM + PPI | D1, D4, D7 | 0 | 30 | 70 | 0.3 ± 0.5 * | 3.20 ± 4.24 * |
| VIII | Yes | AMX + S-GM + PPI | D1 | 60 | 30 | 10 | 2.3 ± 1.1 | 3860.59 ± 3425.28 * |
| Group | HP Infection | IL-8 (pg/mL) | TNF-α (pg/mL) | Atrophy | Fecal Microbiome | |||
|---|---|---|---|---|---|---|---|---|
| Alpha Diversity | Abundance (%) | |||||||
| Shannon | Chao 1 | Bacteroidetes | Firmicutes | |||||
| I | No | 17.13 ± 5.66 * | 225.00 ± 253.55 | 0.00 ± 0.00 | 3.86 ± 0.71 | 523.58 ± 96.92 | 22.47 | 55.81 |
| II | Yes | 31.48 ± 6.37 | 745.00 ± 485.64 | 1.60 ± 0.52 | 3.78 ± 0.60 | 483.81 ± 81.67 | 28.84 | 53.87 |
| III | Yes | 18.60 ± 9.06 * | 564.50 ± 549.98 | 1.50 ± 0.85 | 2.92 ± 0.53 ** | 245.71 ± 121.23 ** | 1.59 ** | 58.67 |
| IV | Yes | 14.70 ± 6.70 * | 442.50 ± 328.69 | 1.20 ± 0.79 | 3.13 ± 0.55 | 440.45 ± 213.56 † | 29.36 † | 29.72 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, S.J.; Lee, K.H.; Kim, J.-H.; Park, S.Y.; Song, Y.G. Efficacy and Gut Dysbiosis of Gentamicin-Intercalated Smectite as a New Therapeutic Agent against Helicobacter pylori in a Mouse Model. Antibiotics 2020, 9, 502. https://doi.org/10.3390/antibiotics9080502
Jeong SJ, Lee KH, Kim J-H, Park SY, Song YG. Efficacy and Gut Dysbiosis of Gentamicin-Intercalated Smectite as a New Therapeutic Agent against Helicobacter pylori in a Mouse Model. Antibiotics. 2020; 9(8):502. https://doi.org/10.3390/antibiotics9080502
Chicago/Turabian StyleJeong, Su Jin, Kyoung Hwa Lee, Jie-Hyun Kim, Soon Young Park, and Young Goo Song. 2020. "Efficacy and Gut Dysbiosis of Gentamicin-Intercalated Smectite as a New Therapeutic Agent against Helicobacter pylori in a Mouse Model" Antibiotics 9, no. 8: 502. https://doi.org/10.3390/antibiotics9080502
APA StyleJeong, S. J., Lee, K. H., Kim, J.-H., Park, S. Y., & Song, Y. G. (2020). Efficacy and Gut Dysbiosis of Gentamicin-Intercalated Smectite as a New Therapeutic Agent against Helicobacter pylori in a Mouse Model. Antibiotics, 9(8), 502. https://doi.org/10.3390/antibiotics9080502