Genetic Determinants for Metal Tolerance and Antimicrobial Resistance Detected in Bacteria Isolated from Soils of Olive Tree Farms


Abstract
1. Introduction
2. Results
2.1. Genetic Determinants Coding for Heavy Metal Tolerance and Antibiotic Resistance
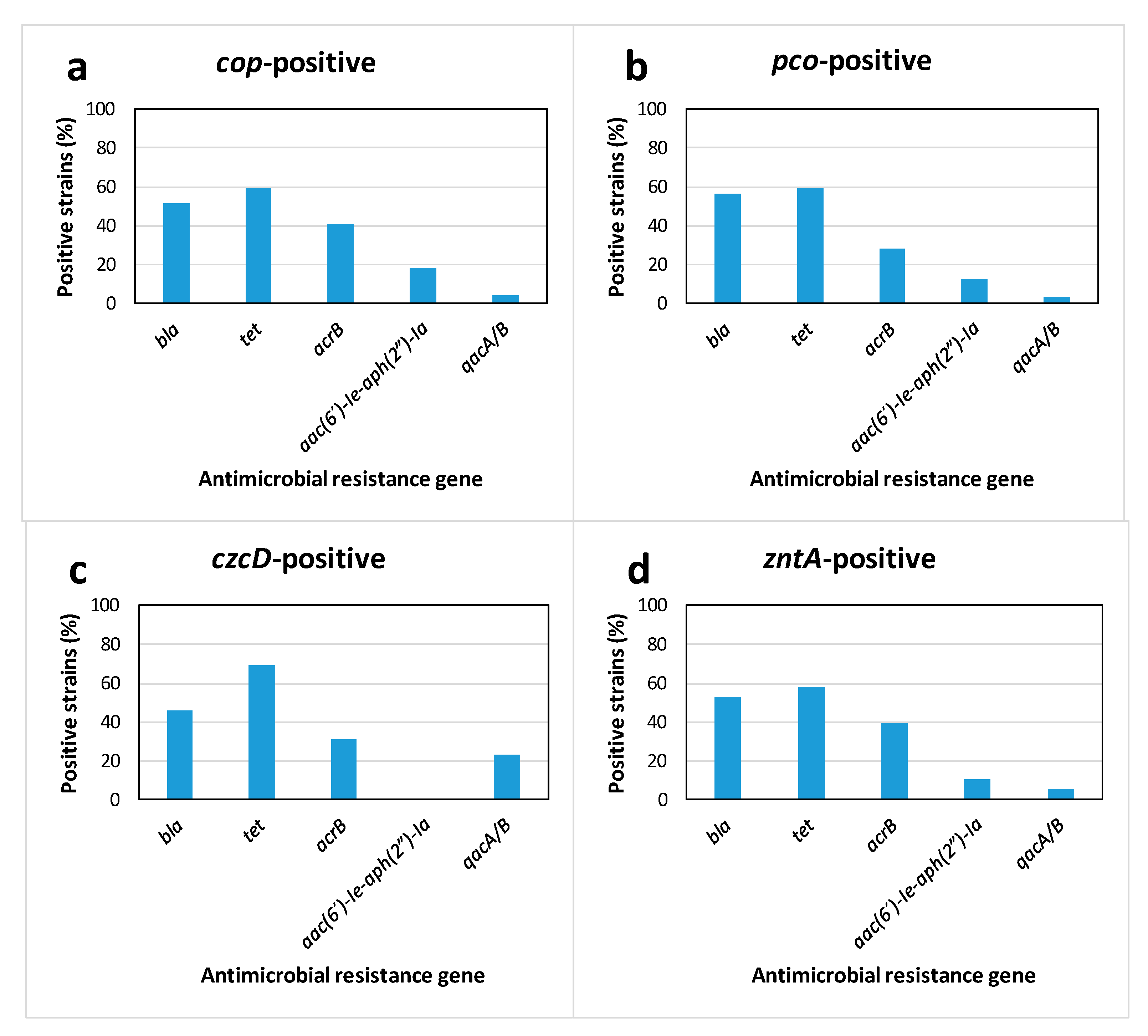
2.2. Association between Heavy Metal Tolerance and Genetic Determinants for Antibiotic Resistance
3. Discussion
4. Materials and Methods
4.1. Strain Selection
4.2. PCR Detection of Antimicrobial Resistance Genes
4.3. Heavy Metals Tolerance Genes
4.4. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ashbolt, N.J.; Amezquita, A.; Backhaus, T.; Borriello, P.; Bandt, K.K.; Collignon, P.; Coors, A.; Finley, R.; Gaze, W.H.; Heberer, T.; et al. Human health risk assessment (HHRA) for environmental development and transfer of antibiotic resistance. Environ. Health Perspect. 2013, 121, 993–1001. [Google Scholar] [CrossRef]
- Marti, E.; Variatza, E.; Balcazar, J.L. The role of aquatic ecosystems as reservoirs of antibiotic resistance. Trends Microbiol. 2014, 22, 36–41. [Google Scholar] [CrossRef]
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Bürgmann, H.; Sørum, H.; Norström, M.; Pons, M.N.; et al. Tackling antibiotic resistance: The environmental framework. Nat. Rev. Microbiol. 2015, 13, 310–317. [Google Scholar] [CrossRef]
- Singer, A.C.; Shaw, H.; Rhodes, V.; Hart, A. Review of antimicrobial resistance in the environment and its relevance to environmental regulators. Front. Microbiol. 2016, 7, 1728. [Google Scholar] [CrossRef]
- Knapp, C.W.; McCluskey, S.M.; Singh, B.K.; Campbell, C.D.; Hudson, G.; Graham, D.W. Antibiotic resistance gene abundances correlated with metal and geochemical conditions in archived Scottish soils. PLoS ONE 2011, 6, e27300. [Google Scholar] [CrossRef]
- Zhu, Y.G.; Johnson, T.A.; Su, J.Q.; Qiao, M.; Guo, G.X.; Stedtfeld, R.D.; Hashsham, S.A.; Tiedje, J.M. Diverse and abundant antibiotic resistance genes in Chinese swine farms. Proc. Nat. Acad. Sci. USA 2013, 110, 3435–3444. [Google Scholar] [CrossRef]
- Wales, A.; Davies, R.H. Co-Selection of Resistance to Antibiotics, Biocides and Heavy Metals, and Its Relevance to Foodborne Pathogens. Antibiotics 2015, 4, 567–604. [Google Scholar] [CrossRef]
- Icgen, B.; Yilmaz, F. Co-occurrence of antibiotic and heavy metal resistance in Kızılırmak river isolates. Bull. Environ. Contam. Toxicol. 2014, 93, 735–743. [Google Scholar] [CrossRef]
- Liu, B.; Li, Y.; Gao, S.; Chen, X. Copper exposure to soil under single and repeated application: Selection for the microbial community tolerance and effects on the dissipation of antibiotics. J. Hazard. Mater. 2017, 325, 129–135. [Google Scholar] [CrossRef]
- Poole, K. At the Nexus of Antibiotics and Metals: The Impact of Cu and Zn on Antibiotic Activity and Resistance. Trends Microbiol. 2017, 25, 820–832. [Google Scholar] [CrossRef]
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance; WHO Press: Geneva, Switzerland, 2014. [Google Scholar]
- Medardus, J.J.; Molla, B.Z.; Nicol, M.; Morrow, W.M.; Rajala-Schultz, P.J.; Kazwala, R.; Gebreyes, W.A. In-feed use of heavy metal micronutrients in US swine production systems and its role in persistence of multidrug-resistant salmonellae. Appl. Environ. Microbiol. 2014, 80, 2317–2325. [Google Scholar] [CrossRef] [PubMed]
- Slifierz, M.J.; Friendship, R.; Weese, J.S. Zinc oxide therapy increases preva-lence and persistence of methicillin-resistant Staphylococcus aureus in pigs: A randomized controlled trial. Zoonoses Public Health 2015, 62, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Amachawadi, R.G.; Scott, H.M.; Nitikanchana, S.; Vinasco, J.; Tokach, M.D.; Dritz, S.S.; Nelssen, J.L.; Goodband, R.D.; Nagaraja, T.G. Nasal carriage of mecA-positive methicillin-resistant Staphylococcus aureus in pigs exhibits dose-response to zinc supplementation. Foodborne Pathog. Dis. 2015, 12, 159–163. [Google Scholar] [CrossRef]
- Li, L.G.; Xia, Y.; Zhang, T. Co-occurrence of antibiotic and metal resistance genes revealed in complete genome collection. ISME J. 2017, 11, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Staehlin, B.M.; Gibbons, J.G.; Rokas, A.; O’Halloran, T.V.; Slot, J.C. Evolution of a heavy metal homeostasis/resistance island reflects increasing copper stress in enterobacteria. Genome Biol. Evol. 2016, 8, 811–826. [Google Scholar] [CrossRef]
- Berg, J.; Thorsen, M.K.; Holm, P.E.; Jensen, J.; Nybroe, O.; Brandt, K.K. Cu exposure under field conditions coselects for antibiotic resistance as determined by a novel cultivation-independent bacterial community tolerance assay. Environ. Sci. Technol. 2010, 44, 8724–8728. [Google Scholar] [CrossRef]
- Bondarczuk, K.; Piotrowska-Seget, Z. Molecular basis of active copper resistance mechanisms in Gram-negative bacteria. Cell Biol. Toxicol. 2013, 29, 397–405. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, L.; Ye, C.; Yu, X. Co-selection of antibiotic resistance via copper shock loading on bacteria from a drinking water bio-filter. Environ. Pollut. 2018, 233, 132–141. [Google Scholar] [CrossRef]
- Xu, Y.; Xu, J.; Mao, D.; Luo, Y. Effect of the selective pressure of sub-lethal level of heavy metals on the fate and distribution of ARGs in the catchment scale. Environ. Pollut. 2017, 220, 900–908. [Google Scholar] [CrossRef]
- Glibota, N.; Grande Burgos, M.J.; Gálvez, A.; Ortega, E. Copper tolerance and antibiotic resistance in soil bacteria from olive tree agricultural fields routinely treated with copper compounds. J. Sci. Food Agric. 2019, 99, 4677–4685. [Google Scholar] [CrossRef]
- Yu, Z.; Gunn, L.; Wall, P.; Fanning, S. Antimicrobial resistance and its association with tolerance to heavy metals in agriculture production. Food Microbiol. 2017, 64, 23–32. [Google Scholar] [CrossRef]
- Seiler, C.; Berendonk, T.U. Heavy metal driven co-selection of antibiotic resistance in soil and water bodies impacted by agriculture and aquaculture. Front. Microbiol. 2012, 3, 399. [Google Scholar] [CrossRef]
- Fard, R.M.; Heuzenroeder, M.W.; Barton, M.D. Antimicrobial and heavy metal resistance in commensal enterococci isolated from pigs. Vet. Microbiol. 2011, 48, 276–282. [Google Scholar] [CrossRef]
- Hu, H.W.; Wang, J.T.; Li, J.; Li, J.J.; Ma, Y.B.; Chen, D.; He, J.Z. Field-based evidence for copper contamination induced changes of antibiotic resistance in agricultural soils. Environ. Microbiol. 2016, 18, 3896–3909. [Google Scholar] [CrossRef]
- Knapp, C.W.; Callan, A.C.; Aitken, B.; Shearn, R.; Koenders, A.; Hinwood, A. Relationship between antibiotic resistance genes and metals in residential soil samples from Western Australia. Environ. Sci. Pollut. Res. Int. 2017, 24, 2484–2494. [Google Scholar] [CrossRef]
- Pal, C.; Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G. Co-occurrence of resistance genes to antibiotics; biocides and metals reveals novel insights into their co-selection potential. BMC Genomics 2015, 16, 964. [Google Scholar] [CrossRef]
- Fang, L.; Li, X.; Li, L.; Li, S.; Liao, X.; Sun, J.; Liu, Y. Co-spread of metal and antibiotic resistance within ST3-IncHI2 plasmids from E. coli isolates of food-producing animals. Sci. Rep. 2007, 6, 25312. [Google Scholar]
- Campos, J.; Cristino, L.; Peixe, L.; Antunes, P. MCR-1 in multidrug-resistant and copper-tolerant clinically relevant Salmonella 1;4;[5];12:i:- and S. Rissen clones in Portugal; 2011 to 2015. Eurosurveillance 2016, 21, 30270. [Google Scholar] [CrossRef]
- Pal, C.; Asiani, K.; Arya, S.; Rensing, C.; Stekel, D.J.; Larsson, D.G.J.; Hobman, J.L. Metal resistance and its association with antibiotic resistance. Adv. Microb. Physiol. 2017, 70, 261–313. [Google Scholar]
- Perron, K.; Caille, O.; Rossier, C.; Van Delden, C.; Dumas, J.; Köhler, T. CzcR-CzcS, a two-component system involved in heavy metal and carbapenem resistance in Pseudomonas aeruginosa. J. Biol. Chem. 2004, 279, 8761–8768. [Google Scholar] [CrossRef]
- Barbosa, T.M.; Phelan, R.W.; Leong, D.; Morrissey, J.P.; Adams, C.; Dobson, A.D.W.; O’Gara, F. A novel erythromycin resistance plasmid from Bacillus sp. strain HS24, isolated from the marine sponge Haliclona simulans. PLoS ONE 2014, 9, e115583. [Google Scholar] [CrossRef]
- Moradali, M.F.; Ghods, S.; Rehm, B.H.A. Pseudomonas aeruginosa Lifestyle: A Paradigm for Adaptation, Survival, and Persistence. Front. Cell. Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef]
- Sullivan, R.; Schaus, D.; John, M.; Delport, J.A. Extended spectrum beta-lactamases: A minireview of clinical relevant groups. J. Med. Microb. Diagn. 2016, 4, 203. [Google Scholar] [CrossRef]
- Hong, D.J.; Bae, I.K.; Jang, I.H.; Jeong, S.H.; Kang, H.K.; Lee, K. Epidemiology and characteristics of metallo-β-lactamase-producing Pseudomonas aeruginosa. Infect. Chemother. 2015, 47, 81–97. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. Environmental factors influencing the development and spread of antibiotic resistance. FEMS Microbiol. Rev. 2018, 42, 1. [Google Scholar] [CrossRef]
- Armalytė, J.; Skerniškytė, J.; Bakienė, E.; Krasauskas, R.; Šiugždinienė, R.; Kareivienė, V.; Kerzienė, S.; Klimienė, I.; Sužiedėlienė, E.; Ružauskas, M. Microbial Diversity and Antimicrobial Resistance Profile in Microbiota From Soils of Conventional and Organic Farming Systems. Front. Microbiol. 2019, 10, 892. [Google Scholar] [CrossRef]
- Youenou, B.; Favre-Bonté, S.; Bodilis, J.; Brothier, E.; Dubost, A.; Muller, D.; Nazaret, S. Comparative genomics of environmental and clinical Stenotrophomonas maltophilia strains with different antibiotic resistance profiles. Genome Biol. Evol. 2015, 7, 2484–2505. [Google Scholar] [CrossRef]
- Wang, Y.; He, T.; Shen, Z.; Wu, C. Antimicrobial Resistance in Stenotrophomonas spp. Microbiol. Spectr. 2018, 6, ARBA-0005-2017. [Google Scholar]
- Lin, X.-H.; Xu, Y.-H.; Sun, X.-H.; Huang, Y.; Li, J.-B. Genetic diversity analyses of antimicrobial resistance genes in clinical Chryseobacterium meningosepticum isolated from Hefei, China. Int. J. Antimicrob. Agents 2012, 40, 186–188. [Google Scholar] [CrossRef]
- Dolejska, M.; Villa, L.; Poirel, L.; Nordmann, P.; Carattoli, A. Complete sequencing of an IncHI1 plasmid encoding the carbapenemase NDM-1, the ArmA 16S RNA methylase and a resistance-nodulation-cell division/multidrug efflux pump. J. Antimicrob. Chemother. 2013, 68, 34–39. [Google Scholar] [CrossRef]
- Walsh, F.; Duffy, B. The culturable soil antibiotic resistome: A community of multi-drug resistant bacteria. PLoS ONE 2013, 8, e65567. [Google Scholar] [CrossRef] [PubMed]
- Swick, M.C.; Morgan-Linnell, S.K.; Carlson, K.M.; Zechiedrich, L. Expression of multidrug efflux pump genes acrAB-tolC; mdfA; and norE in Escherichia coli clinical isolates as a function of fluoroquinolone and multidrug resistance. Antimicrob. Agents Chemother. 2011, 55, 921–924. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.S.; Jensen, L.B.; Sørensen, H.I. Substrate specificity of the OqxAB multidrug resistance pump in Escherichia coli and selected enteric bacteria. J. Antimicrob. Chemother. 2007, 60, 145–147. [Google Scholar] [CrossRef]
- Noguchi, N.; Hase, M.; Kitta, M.; Sasatsu, M.; Deguchi, K.; Kono, M. Susceptibilities to antiseptic agents and distribution of antiseptic-resistance genes qacA/B and smr of methicillin-resistant Staphylococcus aureus isolated in Asia during 1998 and 1999. J. Med. Microbiol. 2005, 54, 557–565. [Google Scholar] [CrossRef]
- Chuanchuen, R.; Khemtong, S.; Padungtod, P. Occurrence of qacE/qacEDelta1 genes and their correlation with class 1 integrons in Salmonella enterica isolates from poultry and swine. Southeast Asian J. Trop. Med. Public Health 2007, 38, 855–862. [Google Scholar] [PubMed]
- Sáenz, Y.; Briñas, L.; Domínguez, E.; Ruiz, J.; Zarazaga, M.; Vila, J.; Torres, C. Mechanisms of resistance in multiple-antibiotic-resistant Escherichia coli strains of human; animal; and food origins. Antimicrob. Agents Chemother. 2004, 48, 3996–4001. [Google Scholar] [CrossRef]
- Vakulenko, S.B.; Donabedian, S.M.; Voskresenskiy, A.M.; Zervos, M.J.; Lerner, S.A.; Chow, J.W. Multiplex PCR for detection of aminoglycoside resistance genes in enterococci. Antimicrob. Agents Chemother. 2003, 47, 1423–1426. [Google Scholar] [CrossRef]
- Chiu, C.H.; Su, L.H.; Chu, C.H.; Wang, M.H.; Yeh, C.M.; Weill, F.X.; Chu, C. Detection of multidrug-resistant Salmonella enterica serovar Typhimurium phage types DT102; DT104; and U302 by multiplex PCR. J. Clin. Microbiol. 2006, 44, 2354–2358. [Google Scholar] [CrossRef]
- Bertrand, S.; Weill, F.X.; Cloeckaert, A.; Vrints, M.; Mairiaux, E.; Praud, K.; Dierick, K.; Wildemauve, C.; Godard, C.; Butaye, P.; et al. Clonal emergence of extended-spectrum β-lactamase (CTX-M-2)-producing Salmonella enterica serovar Virchow isolates with reduced susceptibilities to ciprofloxacin among poultry and humans in Belgium and France (2000 to 2003). J. Clin. Microbiol. 2006, 44, 2897–2903. [Google Scholar] [CrossRef]
- Ng, L.K.; Martin, I.; Alfa, M.; Mulvey, M. Multiplex PCR for the detection of tetracycline resistant genes. Mol. Cell. Probes. 2001, 15, 209–215. [Google Scholar] [CrossRef]
- Kamika, I.; Momba, M.N. Assessing the resistance and bioremediation ability of selected bacterial and protozoan species to heavy metals in metal-rich industrial wastewater. BMC Microbiol. 2013, 13, 28. [Google Scholar] [CrossRef] [PubMed]
- Badar, U.; Ahmed, N.; Shoeb, E.; Gadd, G.M. Identification of the pco operon in Enterobacter species isolated from contaminated soil. Int. J. Adv. Res. 2014, 2, 227–233. [Google Scholar]
- Mourão, J.; Novais, C.; Machado, J.; Peixe, L.; Antunes, P. Metal tolerance in emerging clinically relevant multidrug-resistant Salmonella enterica serotype 4,[5],12:i:− Clones circulating in Europe. Int. J. Antimicrob. Agents 2015, 45, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Chihomvu, P.; Stegmann, P.; Pillay, M. Characterization and Structure Prediction of Partial Length Protein Sequences of pcoA; pcoR and chrB Genes from Heavy Metal Resistant Bacteria from the Klip River; South Africa. Int. J. Mol. Sci. 2015, 16, 7352–7374. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.L.; Rouch, D.A.; Lee, B.T. Copper resistance determinants in bacteria. Plasmid 1992, 27, 41–51. [Google Scholar] [CrossRef]
- Adaikkalam, V.; Swarup, S. Molecular characterization of an operon; cueAR; encoding a putative P1-type ATPase and a MerR-type regulatory protein involved in copper homeostasis in Pseudomonas putida. Microbiology 2002, 148, 2857–2867. [Google Scholar] [CrossRef] [PubMed]
- Hasman, H.; Kempf, I.; Chidaine, B.; Cariolet, R.; Ersbøll, A.K.; Houe, H.; Aarestrup, F.M. Copper resistance in Enterococcus faecium, mediated by the tcrB gene, is selected by supplementation of pig feed with copper sulfate. Appl. Environ. Microbiol. 2006, 72, 5784–5789. [Google Scholar] [CrossRef]
- Silver, S. Bacterial silver resistance: Molecular biology and uses and misuses of silver compounds. FEMS Microbiol. Rev. 2003, 27, 341–353. [Google Scholar] [CrossRef]
- García Fernández, A.; Cloeckaert, A.; Bertini, A.; Praud, K.; Doublet, B.; Weill, F.X.; Carattoli, A. Comparative analysis of IncHI2 plasmids carrying blaCTX-M-2 or blaCTX-M-9 from Escherichia coli and Salmonella enterica strains isolated from poultry and humans. Antimicrob. Agents Chemother. 2007, 51, 4177–4180. [Google Scholar] [CrossRef]
- Liebert, C.A.; Wireman, J.; Smith, T.; Summers, A.O. Phylogeny of mercury resistance (mer) operons of gram-negative bacteria isolated from the fecal flora of primates. Appl. Environ. Microbiol. 1997, 63, 1066–1076. [Google Scholar] [CrossRef]
- Nies, A.; Nies, D.H.; Silver, S. Nucleotide sequence and expression of a plasmid-encoded chromate resistance determinant from Alcaligenes eutrophus. J. Biol. Chem. 1990, 265, 5648–5653. [Google Scholar] [PubMed]
- Naik, M.M.; Shamim, K.; Dubey, S.K. Biological characterization of lead resistant bacteria to explore role of bacterial metallothionein in lead resistance. Curr. Sci. 2012, 103, 426–429. [Google Scholar]
- Borremans, B.; Hobman, J.L.; Provoost, A.; Brown, N.L.; van Der Lelie, D. Cloning and functional analysis of thepbr lead resistance determinant of Ralstonia metallidurans CH34. J. Bacteriol. 2001, 183, 5651–5658. [Google Scholar] [CrossRef] [PubMed]
- Abou-Shanab, R.; Van Berkum, P.; Angle, J. Heavy metal resistance and genotypic analysis of metal resistance genes in gram-positive and gram-negative bacteria present in Ni-rich serpentine soil and in the rhizosphere of Alyssum murale. Chemosphere 2007, 68, 360–367. [Google Scholar] [CrossRef]
- Rensing, C.; Mitra, B.; Rosen, B.P. The zntA gene of Escherichia coli encodes a Zn (II)-translocating P-type ATPase. Proc. Natl. Acad. Sci. USA 1997, 94, 14326–14331. [Google Scholar] [CrossRef]


{kind=link}
| Species (Isolate) | Heavy Metal Tolerance Genetic Determinants | Antibiotic Resistance Genetic Determinants |
|---|---|---|
| Bacillus cereus (T22Pb3) | - | acrB |
| Bacillus endophyticus (T37Ni3) | - | tetA, tetC |
| Bacillus fordii (T11Ni1) | chrB | - |
| Bacillus fordii (T18Ni1) | pcoR, copA | qacA/B |
| Bacillus megaterium (T1Ni2) | pcoR, chrB, copA | intl1, tetA |
| Bacillus psychrosaccharolyticus (T4Ni3) | copA | intl1, tetA, tetC, blaPSE |
| Bacillus sp. (T17Ni3) | - | tetB, blaPSE, blaTEM |
| Bacillus sp. (T19Pb3) | - | blaCTX−M |
| Bacillus sp. (T20Pb2) | pcoD | acrB, aac(6′)-Ie-aph(2″)-Ia, tetA, tetE |
| Bacillus sp. (T21Zn1) | pcoA | blaCTX−M |
| Bacillus sp. (T28Pb3) | - | acrB, blaTEM |
| Bacillus sp. (T11Pb1) | - | acrB, blaCTX−M |
| Bacillus sp. (T12Pb1) | - | acrB |
| Bacillus sp. (T13Ni2) | - | intl1, tetG, blaCTX−M |
| Bacillus sp. (T13Pb1) | - | acrB, blaCTX−M |
| Bacillus sp. (T14Pb1) | - | intl1 |
| Bacillus sp. (T26Ni3) | smtA | acrB, qacA/B, intl1, dfrA12, tetG, blaCTX−M, blaCTX−M2 |
| Bacillus sp. (T26Pb2) | - | acrB |
| Bacillus sp. (T37Ni2) | - | dfrA12 |
| Bacillus sp. (T8Pb1) | - | acrB |
| Bacillus sp. (T9Ni2) | - | blaCTX−M, blaCTX−M2 |
| Bacillus sp. (T9Pb1) | - | - |
| Burkholderia zhejiangensis (T37Cd2) | zntA | tetA |
| Burkholderia zhejiangensis (T38Zn3) | zntA | qacA/B, blaCTX−M |
| Chryseobacterium formosense (T11Zn1) | - | tetA, tetC, blaTEM |
| Chryseobacterium gleum (T14Cd2) | - | tetC |
| Chryseobacterium gleum (T4Ni2) | - | tetA, tetC, blaCTX−M2 |
| Chryseobacterium gleum (T6Pb1) | pcoR, zntA | acrB, tetC, tetG, blaCTX−M |
| Chryseobacterium gleum (T6Zn3) | pcoR, copB | tetG, blaCTX−M, blaCTX−M2 |
| Chryseobacterium hispalense (T3Zn3) | - | tetA, tetC, blaCTX−M |
| Chryseobacterium oranimense (E5Zn1) | zntA | acrB, tetD, blaCTX−M |
| Chryseobacterium oranimense (T18Cd1) | - | acrB, qacA/B, tetA, blaCTX−M2 |
| Chryseobacterium oranimense (T30Zn1) | - | aac(6′)-Ie-aph(2″)-Ia, tetA, tetC |
| Chryseobacterium oranimense (T31Zn1) | pcoR, zntA | tetC |
| Chryseobacterium oranimense (T37Cd3) | pcoR | - |
| Chryseobacterium oranimense (T3Cd1) | - | acrB, tetA, tetC, blaCTX−M |
| Chryseobacterium oranimense (T3Ni2) | pcoR | tetA, tetD, blaCTX−M |
| Chryseobacterium oranimense (T3Zn2) | - | tetA, tetE, blaCTX−M |
| Chryseobacterium oranimense (T4Cd1) | pcoR | acrB, blaCTX−M |
| Chryseobacterium oranimense (T5Cd1) | pcoR | tetA, tetC, blaCTX−M |
| Chryseobacterium oranimense (T5Ni3) | pcoR | tetA, tetC, blaCTX−M |
| Chryseobacterium oranimense (T6Cd1) | - | acrB, intl1, tetA, tetC, blaCTX−M |
| Chryseobacterium piperi (T8Cd1) | - | acrB, qacA/B, tetA, tetC, blaCTX−M |
| Enterobacter (Klebsiella) aerogenes (E1Pb3) | copB, copC, pcoD, tcrB, merA, zntA | blaTEM |
| Enterococcus faecalis (T26Zn2) | pcoR | tetA, tetB, tetD, blaCTX−M |
| Flavobacterium johnsoniae (E1Zn1) | - | acrB, qacE∆1, tetA, tetC |
| Janthinobacterium lividum (T25Cd3) | - | - |
| Pseudomonas entomophila (E1Cu3) | copA, pcoA, czcD, smtA, zntA | acrB, intl1, tetA, blaCTX−M |
| Pseudomonas entomophila (T11Cd1) | - | tetA, blaCTX−M2, blaTEM |
| Pseudomonas entomophila (T17Pb1) | copA, zntA | acrB, tetA, tetG, blaTEM |
| Pseudomonas entomophila (T23Cu2) | copA, zntA | acrB, tetA |
| Pseudomonas entomophila (T34Cu2) | copA, zntA | acrB, intl1, aac(62)-Ie-aph(2″)-Ia |
| Pseudomonas entomophila (T5Cu2) | copA, smtA | acrB, blaCTX−M |
| Pseudomonas entomophila (T6Cu1) | czcD, smtA, zntA | acrB, tetA |
| Pseudomonas entomophila (T7Ni1) | copA, pcoD, czcD, smtA, zntA | - |
| Pseudomonas fluorescens (E5Zn2) | copA, pcoD, zntA | aac(6′)-Ie-aph(2″)-Ia, tetG, blaCTX−M |
| Pseudomonas fluorescens (E5Zn3) | copA, pcoD, zntA | acrB, aac(6′)-Ie-aph(2″)-Ia, blaCTX−M, blaPSE |
| Pseudomonas fluorescens (T12Cu1) | pcoR, pcoD, zntA | blaCTX−M |
| Pseudomonas fluorescens (T15Cd1) | chrB, czcD, zntA | tetA |
| Pseudomonas fluorescens (T25Cu3) | zntA | tetB, blaCTX−M2 |
| Pseudomonas fluorescens (T25Ni3) | czcD | qacA/B, tetG |
| Pseudomonas fluorescens (T31Cd3) | czcD, zntA | qacA/B, tetD, blaPSE, blaTEM |
| Pseudomonas fluorescens (T32Cd3) | pcoR, copA, zntA | tetA |
| Pseudomonas fluorescens (T35Cd3) | copA, czcD, zntA | tetA, blaTEM |
| Pseudomonas fluorescens (T35Cu2) | copA, pcoD, smtA | acrB, tetA |
| Pseudomonas fluorescens (T3Ni3) | copA, pcoD | tetA, blaTEM |
| Pseudomonas fluorescens (T5Cd2) | pcoR, chrB, czcD, zntA | tetA, blaCTX−M, blaTEM |
| Pseudomonas fluorescens (T7Cd3) | pcoR, zntA | tetA |
| Pseudomonas lutea (E3Cu2) | czcD, zntA | intl1 |
| Pseudomonas lutea (T8Ni2) | - | - |
| Pseudomonas putida (T1Cu3) | copA, copC, czcD, smtA | acrB, intl1, tetE, blaCTX−M, blaCTX−M2, blaTEM |
| Pseudomonas sp. (T15Ni1) | copA, zntA | dfrA12, tetE |
| Pseudomonas sp. (T16Ni3) | - | tetA, blaPSE |
| Pseudomonas sp. (T15Cu3) | copA, zntA | aac(6′)-Ie-aph(2″)-Ia, tetE, blaTEM |
| Pseudomonas sp. (T18Cu3) | copA, silE, zntA | acrB, intl1, tetA, blaCTX−M2 |
| Pseudomonas sp. (T19Cu2) | copA, silE, | acrB, intl1 |
| Pseudomonas sp. (T22Cu1) | pcoR, pcoD, zntA | tetD, tetE, tetG, blaCTX−M |
| Pseudomonas sp. (T24Cu3) | pcoR, zntA | acrB, intl1 |
| Pseudomonas sp. (T24Ni1) | pcoR, copA | acrB |
| Pseudomonas sp. (T28Cu2) | chrB, copA, zntA | acrB, blaTEM |
| Pseudomonas sp. (T36Cu3) | chrB, copA, czcD, smtA, zntA | intl1 |
| Pseudomonas sp. (T40Cd3) | czcD, zntA | qacA/B, acrB, tetB, blaCTX−M2 |
| Pseudomonas sp. (T7Cu2) | pcoD, zntA | acrB |
| Serratia proteamaculans (T3Cd2) | zntA | acrB, tetB, tetE, tetG |
| Serratia proteamaculans (T4Pb1) | pcoR, copA | aac(6′)-Ie-aph(2″)-Ia, tetB, tetG, blaCTX−M, blaPSE |
| Sphingobacterium paucimobilis (T30Zn3) | pcoR, copA, copC | tetC, tetG, blaPSE |
| Stenotrophomonas maltophilia (T19Zn1) | zntA | blaTEM |
| Stenotrophomonas maltophilia (T40Zn1) | zntA | acrB, blaTEM |
| Stenotrophomonas rhizophila (E1Zn3) | pcoA | - |
| Stenotrophomonas rhizophila (T31Zn2) | zntA | - |
| Variovorax paradoxus (E4Zn1) | pcoR | acrB, tetA |
| Variovorax paradoxus (T18Zn2) | czcD, smtA, zntA | tetB |
| Variovorax paradoxus (T23Zn2) | zntA | - |
| Variovorax paradoxus (T32Zn2) | chrB | tetE |
| Variovorax paradoxus (T32Zn3) | chrB | - |
| Variovorax paradoxus (T7Zn1) | - | tetA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glibota, N.; Grande, M.J.; Galvez, A.; Ortega, E. Genetic Determinants for Metal Tolerance and Antimicrobial Resistance Detected in Bacteria Isolated from Soils of Olive Tree Farms. Antibiotics 2020, 9, 476. https://doi.org/10.3390/antibiotics9080476
Glibota N, Grande MJ, Galvez A, Ortega E. Genetic Determinants for Metal Tolerance and Antimicrobial Resistance Detected in Bacteria Isolated from Soils of Olive Tree Farms. Antibiotics. 2020; 9(8):476. https://doi.org/10.3390/antibiotics9080476
Chicago/Turabian StyleGlibota, Nicolás, Mª José Grande, Antonio Galvez, and Elena Ortega. 2020. "Genetic Determinants for Metal Tolerance and Antimicrobial Resistance Detected in Bacteria Isolated from Soils of Olive Tree Farms" Antibiotics 9, no. 8: 476. https://doi.org/10.3390/antibiotics9080476
APA StyleGlibota, N., Grande, M. J., Galvez, A., & Ortega, E. (2020). Genetic Determinants for Metal Tolerance and Antimicrobial Resistance Detected in Bacteria Isolated from Soils of Olive Tree Farms. Antibiotics, 9(8), 476. https://doi.org/10.3390/antibiotics9080476