The Gut of Healthy Infants in the Community as a Reservoir of ESBL and Carbapenemase-Producing Bacteria

 , , and
, , and
Abstract
1. Introduction
2. Methods
2.1. Patient Enrollment and Sample Collection
2.2. Bacterial Isolate Testing and Antimicrobial Sensitivities
2.3. DNA Extraction and Molecular Testing for ESBL and Carbapenemases
2.4. Multi-Locus Sequence Typing (MLST)
2.5. Phylogenetic Analysis
2.6. Statistical Analysis
2.7. Ethical Approval
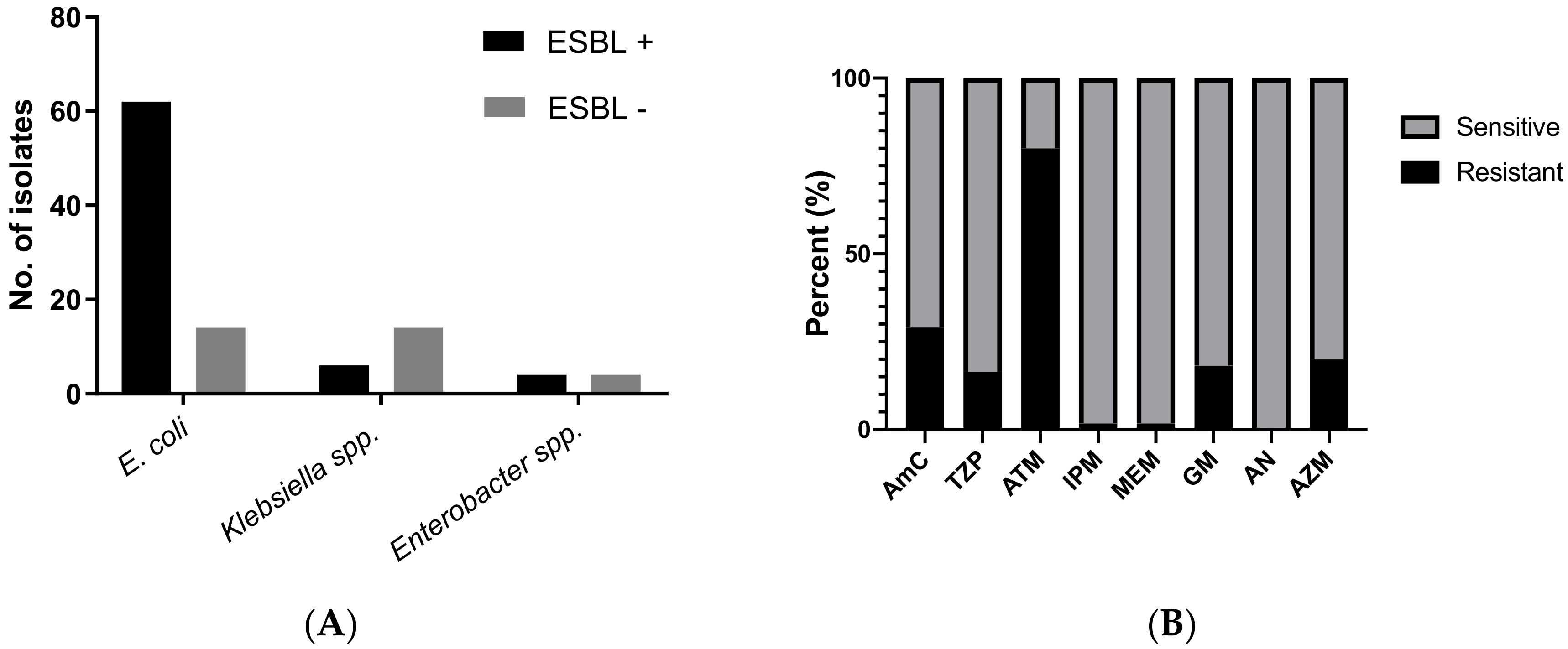
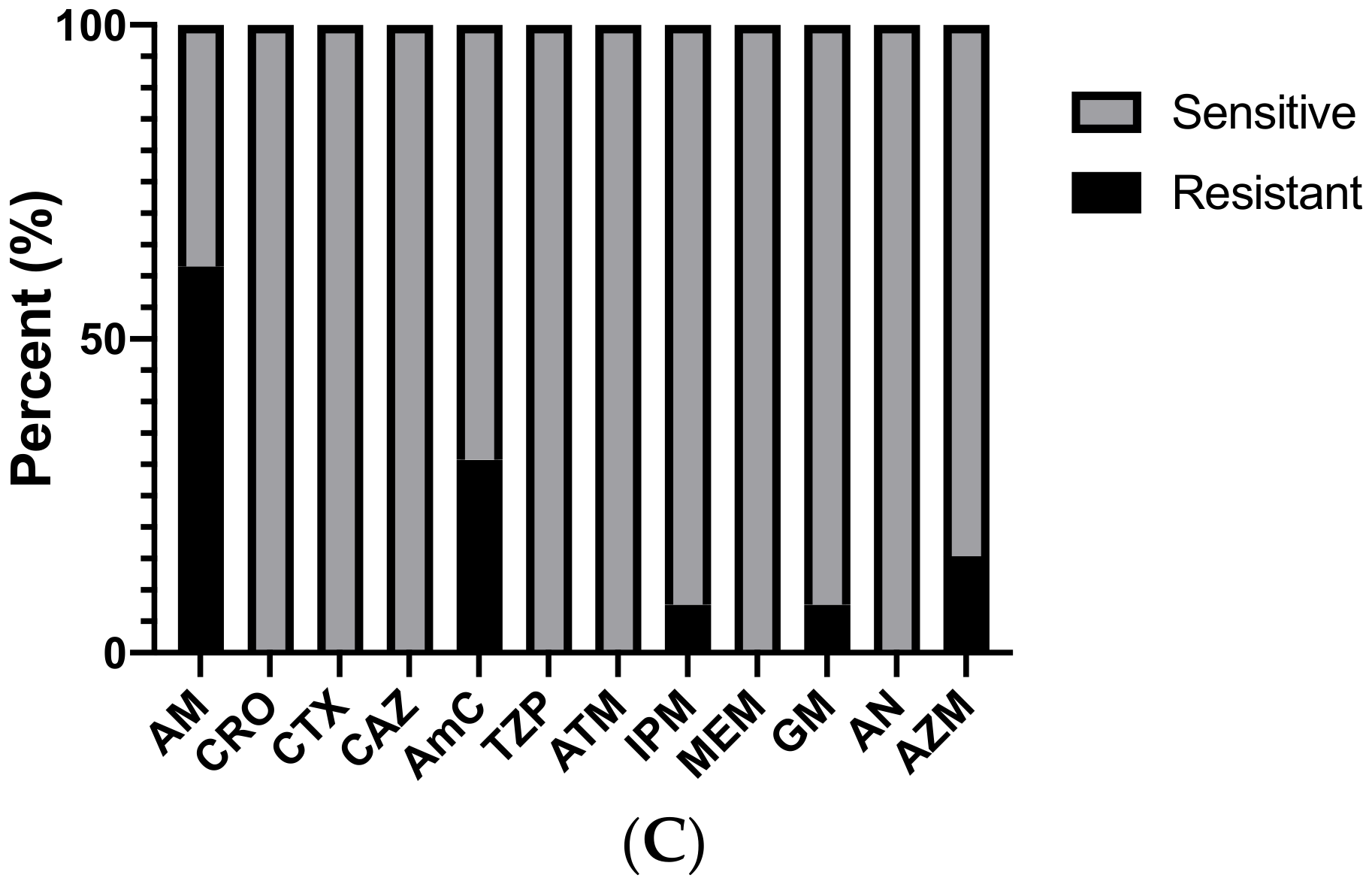
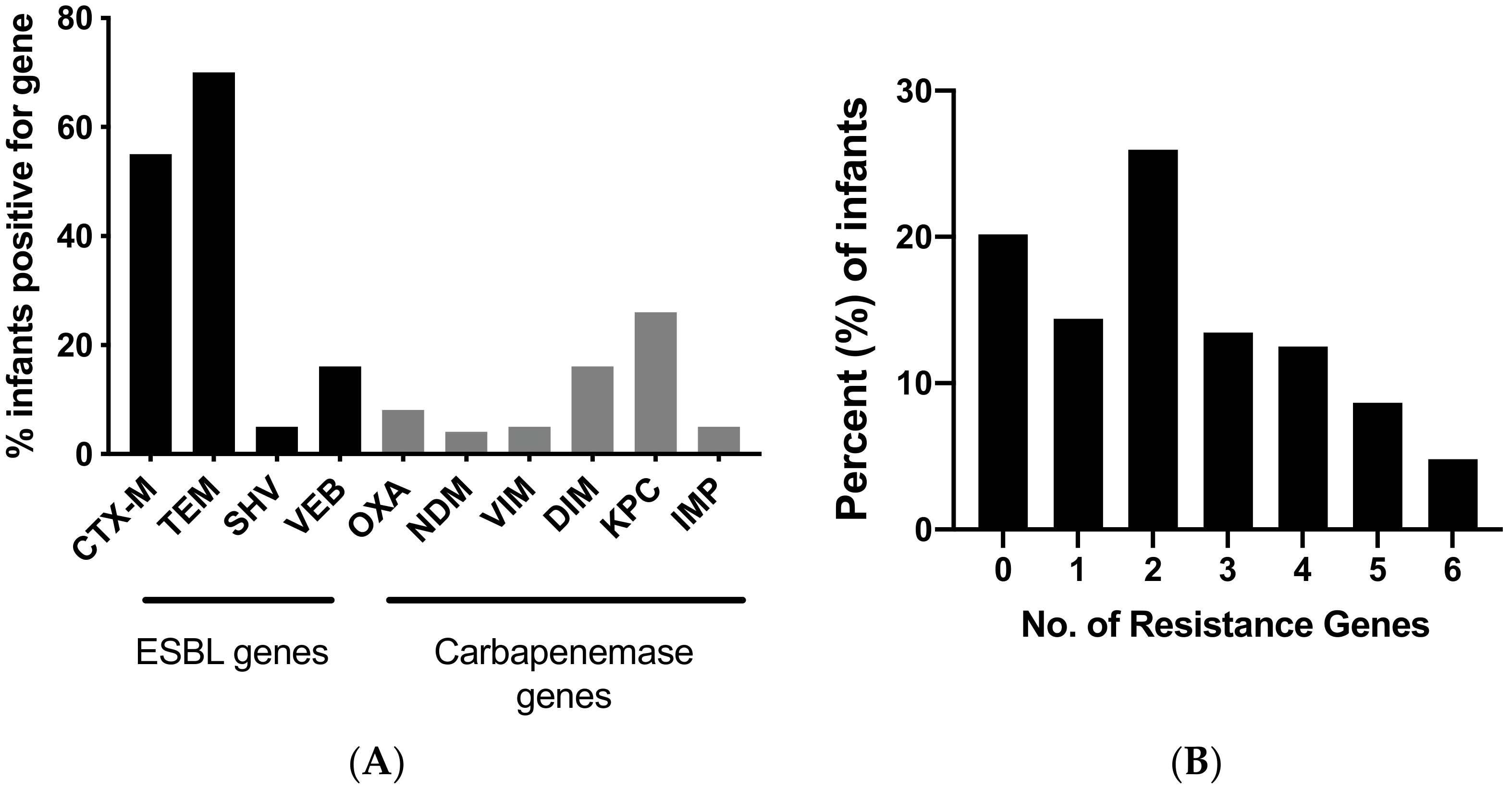
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Waters, D.; Jawad, I.; Ahmad, A.; Luksic, I.; Nair, H.; Zgaga, L.; Theodoratou, E.; Rudan, I.; Zaidi, A.K.; Campbell, H. Aetiology of community-acquired neonatal sepsis in low and middle income countries. J. Glob. Health 2011, 1, 154–170. [Google Scholar] [PubMed]
- Zaidi, A.K.; Thaver, D.; Ali, S.A.; Khan, T.A. Pathogens associated with sepsis in newborns and young infants in developing countries. Pediatr. Infect. Dis. J. 2009, 28, S10–S18. [Google Scholar] [CrossRef]
- Petty, N.K.; Ben Zakour, N.L.; Stanton-Cook, M.; Skippington, E.; Totsika, M.; Forde, B.M.; Phan, M.D.; Gomes Moriel, D.; Peters, K.M.; Davies, M.; et al. Global dissemination of a multidrug resistant Escherichia coli clone. Proc. Natl. Acad. Sci. USA 2014, 111, 5694–5699. [Google Scholar] [CrossRef] [PubMed]
- Litzow, J.M.; Gill, C.J.; Mantaring, J.B.; Fox, M.P.; MacLeod, W.B.; Mendoza, M.; Mendoza, S.; Scobie, R.; Huskins, C.W.; Goldman, D.A.; et al. High frequency of multidrug-resistant gram-negative rods in 2 neonatal intensive care units in the Philippines. Infect. Control Hosp. Epidemiol. 2009, 30, 543–549. [Google Scholar] [CrossRef][Green Version]
- Patel, S.J.; Saiman, L. Antibiotic resistance in neonatal intensive care unit pathogens: Mechanisms, clinical impact, and prevention including antibiotic stewardship. Clin. Perinatol. 2010, 37, 547–563. [Google Scholar] [CrossRef] [PubMed]
- Folgori, L.; Tersigni, C.; Hsia, Y.; Kortsalioudaki, C.; Heath, P.; Sharland, M.; Bielicki, J. The relationship between Gram-negative colonization and bloodstream infections in neonates: A systematic review and meta-analysis. Clin. Microbiol. Infect. 2018, 24, 251–257. [Google Scholar] [CrossRef]
- UNICEF, W. World Bank Group and United Nations. Levels & Trends in Child Mortality: Report 2017, Estimates Developed by the UN Inter-agency Group for Child Mortality Estimation; United Nations Children’s Fund: New York, NY, USA, 2017. [Google Scholar]
- Reardon, S. Antibiotic resistance sweeping developing world. Nature 2014, 509, 141–142. [Google Scholar] [CrossRef]
- Shah, S.Q.; Colquhoun, D.J.; Nikuli, H.L.; Sorum, H. Prevalence of antibiotic resistance genes in the bacterial flora of integrated fish farming environments of Pakistan and Tanzania. Environ. Sci. Technol. 2012, 46, 8672–8679. [Google Scholar] [CrossRef]
- Siddiqi, S.; Hamid, S.; Rafique, G.; Chaudhry, S.A.; Ali, N.; Shahab, S.; Sauerborn, R. Prescription practices of public and private health care providers in Attock District of Pakistan. Int. J. Health Plann. Manag. 2002, 17, 23–40. [Google Scholar] [CrossRef]
- Vimont, S.; Boyd, A.; Bleibtreu, A.; Bens, M.; Goujon, J.M.; Garry, L.; Clermont, O.; Denamur, E.; Arlet, G.; Vandewalle, A. The CTX-M-15-producing Escherichia coli clone O25b: H4-ST131 has high intestine colonization and urinary tract infection abilities. PLoS ONE 2012, 7, e46547. [Google Scholar] [CrossRef]
- Gurnee, E.A.; Ndao, I.M.; Johnson, J.R.; Johnston, B.D.; Gonzalez, M.D.; Burnham, C.A.; Hall-Moore, C.M.; McGhee, J.E.; Mellmann, A.; Warner, B.B.; et al. Gut Colonization of Healthy Children and Their Mothers With Pathogenic Ciprofloxacin-Resistant Escherichia coli. J. Infect. Dis. 2015, 212, 1862–1868. [Google Scholar] [CrossRef] [PubMed]
- Saleem, A.F.; Mach, O.; Yousafzai, M.T.; Khan, A.; Weldon, W.C.; Steven Oberste, M.; Zaidi, S.S.; Alam, M.M.; Quadri, F.; Sutter, R.W.; et al. Immunogenicity of Different Routine Poliovirus Vaccination Schedules: A Randomized, Controlled Trial in Karachi, Pakistan. J. Infect. Dis. 2018, 217, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Bridson, E. The Oxoid Manual; Oxoid Limited: Hampshire, UK, 2006. [Google Scholar]
- Patel, J.B. Performance Standards For Antimicrobial Susceptibility Testing, 28th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Trung, N.T.; Hien, T.T.; Huyen, T.T.; Quyen, D.T.; Binh, M.T.; Hoan, P.Q.; Meyer, C.G.; Velavan, T.P.; Song, L.H. Simple multiplex PCR assays to detect common pathogens and associated genes encoding for acquired extended spectrum betalactamases (ESBL) or carbapenemases from surgical site specimens in Vietnam. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Jaureguy, F.; Landraud, L.; Passet, V.; Diancourt, L.; Frapy, E.; Guigon, G.; Carbonnelle, E.; Lortholary, O.; Clermont, O.; Denamur, E.; et al. Phylogenetic and genomic diversity of human bacteremic Escherichia coli strains. BMC Genom. 2008, 9, 560. [Google Scholar] [CrossRef] [PubMed]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [PubMed]
- Bonacorsi, S.P.; Clermont, O.; Tinsley, C.; Le Gall, I.; Beaudoin, J.C.; Elion, J.; Nassif, X.; Bingen, E. Identification of regions of the Escherichia coli chromosome specific for neonatal meningitis-associated strains. Infect. Immun. 2000, 68, 2096–2101. [Google Scholar] [CrossRef]
- Fuchs, A.; Bielicki, J.; Mathur, S.; Sharland, M.; Van Den Anker, J.N. Reviewing the WHO guidelines for antibiotic use for sepsis in neonates and children. Paediatr. Int. Child Health 2018, 38, S3–S15. [Google Scholar] [CrossRef]
- Hijazi, S.M.; Fawzi, M.A.; Ali, F.M.; Abd El Galil, K.H. Multidrug-resistant ESBL-producing Enterobacteriaceae and associated risk factors in community infants in Lebanon. J. Infect. Dev. Ctries. 2016, 10, 947–955. [Google Scholar] [CrossRef]
- Monira, S.; Shabnam, S.A.; Ali, S.I.; Sadique, A.; Johura, F.T.; Rahman, K.Z.; Alam, N.H.; Watanabe, H.; Alam, M. Multi-drug resistant pathogenic bacteria in the gut of young children in Bangladesh. Gut Pathog. 2017, 9, 19. [Google Scholar] [CrossRef]
- Chan, G.J.; Lee, A.C.C.; Baqui, A.H.; Tan, J.; Black, R.E. Risk of Early-Onset Neonatal Infection with Maternal Infection or Colonization: A Global Systematic Review and Meta-Analysis. PLoS Med. 2013, 10, e1001502. [Google Scholar] [CrossRef]
- Klemm, E.J.; Shakoor, S.; Page, A.J.; Qamar, F.N.; Judge, K.; Saeed, D.K.; Wong, V.K.; Dallman, T.J.; Nair, S.; Baker, S.; et al. Emergence of an Extensively Drug-Resistant Salmonella enterica Serovar Typhi Clone Harboring a Promiscuous Plasmid Encoding Resistance to Fluoroquinolones and Third-Generation Cephalosporins. mBio 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Khan, E.; Irfan, S.; Sultan, B.A.; Nasir, A.; Hasan, R. Dissemination and spread of New Delhi Metallo-beta-lactamase-1 Superbugs in hospital settings. J. Pak. Med. Assoc. 2016, 66, 999–1004. [Google Scholar] [PubMed]
- Qamar, S.; Shaheen, N.; Shakoor, S.; Farooqi, J.; Jabeen, K.; Hasan, R. Frequency of colistin and fosfomycin resistance in carbapenem-resistant Enterobacteriaceae from a tertiary care hospital in Karachi. Infect. Drug Resist. 2017, 10, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Organization, W.H. Guideline: Managing Possible Serious Bacterial Infection in Young Infants When Referral is Not Feasible; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Bush, K.; Jacoby, G.A. Updated functional classification of beta-lactamases. Antimicrob. Agents Chemother. 2010, 54, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Bevan, E.R.; Jones, A.M.; Hawkey, P.M. Global epidemiology of CTX-M beta-lactamases: Temporal and geographical shifts in genotype. J. Antimicrob. Chemother. 2017, 72, 2145–2155. [Google Scholar] [CrossRef] [PubMed]
- Mathai, D.; Kumar, V.A.; Paul, B.; Sugumar, M.; John, K.R.; Manoharan, A.; Kesavan, L.M. Fecal carriage rates of extended-spectrum beta-lactamase-producing Escherichia coli among antibiotic naive healthy human volunteers. Microb. Drug Resist. 2015, 21, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Evans, B.A.; Amyes, S.G. OXA beta-lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef] [PubMed]
- Cao, V.; Lambert, T.; Nhu, D.Q.; Loan, H.K.; Hoang, N.K.; Arlet, G.; Courvalin, P. Distribution of extended-spectrum beta-lactamases in clinical isolates of Enterobacteriaceae in Vietnam. Antimicrob. Agents Chemother. 2002, 46, 3739–3743. [Google Scholar] [CrossRef]
- Girlich, D.; Naas, T.; Leelaporn, A.; Poirel, L.; Fennewald, M.; Nordmann, P. Nosocomial spread of the integron-located veb-1-like cassette encoding an extended-pectrum beta-lactamase in Pseudomonas aeruginosa in Thailand. Clin. Infect. Dis. 2002, 34, 603–611. [Google Scholar] [CrossRef]
- Naas, T.; Poirel, L.; Nordmann, P. Minor extended-spectrum beta-lactamases. Clin. Microbiol. Infect. 2008, 14 (Suppl. 1), 42–52. [Google Scholar] [CrossRef]
- Poirel, L.; Naas, T.; Guibert, M.; Chaibi, E.B.; Labia, R.; Nordmann, P. Molecular and biochemical characterization of VEB-1, a novel class A extended-spectrum beta-lactamase encoded by an Escherichia coli integron gene. Antimicrob. Agents Chemother. 1999, 43, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Rodriguez-Martinez, J.M.; Al Naiemi, N.; Debets-Ossenkopp, Y.J.; Nordmann, P. Characterization of DIM-1, an integron-encoded metallo-beta-lactamase from a Pseudomonas stutzeri clinical isolate in the Netherlands. Antimicrob. Agents Chemother. 2010, 54, 2420–2424. [Google Scholar] [CrossRef] [PubMed]
- Cuzon, G.; Naas, T.; Nordmann, P. Functional characterization of Tn4401, a Tn3-based transposon involved in blaKPC gene mobilization. Antimicrob. Agents Chemother. 2011, 55, 5370–5373. [Google Scholar] [CrossRef] [PubMed]
- Pica-Ciamarra, U.; Tasciotti, L.; Otte, J.; Zezza, A. Livestock in the Household Economy: Cross-Country Evidence from Microeconomic Data. Dev. Policy Rev. 2015, 33, 61–81. [Google Scholar] [CrossRef]
- Rettedal, S.; Lohr, I.H.; Bernhoff, E.; Natas, O.B.; Sundsfjord, A.; Oymar, K. Extended-spectrum beta-lactamase-producing Enterobacteriaceae among pregnant women in Norway: Prevalence and maternal-neonatal transmission. J. Perinatol. 2015, 35, 907–912. [Google Scholar] [CrossRef]
- Picard, B.; Garcia, J.S.; Gouriou, S.; Duriez, P.; Brahimi, N.; Bingen, E.; Elion, J.; Denamur, E. The link between phylogeny and virulence in Escherichia coli extraintestinal infection. Infect. Immun. 1999, 67, 546–553. [Google Scholar] [CrossRef]
- Nowrouzian, F.L.; Wold, A.E.; Adlerberth, I. Escherichia coli strains belonging to phylogenetic group B2 have superior capacity to persist in the intestinal microflora of infants. J. Infect. Dis. 2005, 191, 1078–1083. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Demographic Parameter | N (%) |
|---|---|
| Gender of child | |
| Male | 58 (55) |
| Female | 46 (45) |
| n | 104 |
| Age groups (months) | |
| Median age | 5.1 |
| Interquartile range | 0.36 (5.059–5.420) |
| Birth weight (kg) | |
| Median birth weight | 2.83 |
| Interquartile range | 0.55 (2.56–3.11) |
| History of breastfeeding | |
| Exclusively breast fed | 74 (69.2) |
| Type of milk (if not exclusively breastfed) | |
| Formula milk | 21 (63.6) |
| Cow milk | 5 (15.2) |
| Other (goat milk or weaning) | 7 (21.2) |
| Sequence Type | Frequency (No. of Isolates) | Identification Method 1 | Phylogenetic Group | Reported Source 2 |
|---|---|---|---|---|
| 2 | 5 | PUBMLST | B2 | human/domesticated animal |
| 4 | 1 | PUBMLST | D | human/food |
| 5 | 1 | PUBMLST | D | human |
| 8 | 11 | PUBMLST | D | domesticated animal |
| 21 | 1 | PUBMLST | D | humans/livestock |
| 87 | 1 | PUBMLST | D | primate |
| 132 | 2 | PUBMLST | B2 | animal |
| 244 | 1 | PUBMLST | D | human |
| 316 | 2 | PUBMLST | D | human/fish/poultry/livestock/environment |
| 357 | 1 | PUBMLST | D | human/poultry |
| 398 | 1 | PUBMLST | D | human/livestock/environment |
| 399 | 1 | PUBMLST | D | human/fish/poultry/livestock/environment |
| 511 | 2 | PUBMLST | D | human |
| 535 | 1 | PUBMLST | D | human |
| 537 | 1 | PUBMLST | D | human/livestock |
| 551 | 1 | PUBMLST | D | human |
| 569 | 2 | PUBMLST | B2 | human/environment/poultry |
| 573 | 1 | PUBMLST | D | human |
| 638 | 2 | PUBMLST | B2 | human |
| 647 | 1 | PUBMLST | D | human/livestock |
| 661 | 2 | PUBMLST | D | human/environment/livestock |
| 681 | 1 | PUBMLST | A | human |
| 740 | 1 | PUBMLST | D | human |
| 756 | 1 | PUBMLST | B2 | animal |
| 767 | 1 | PUBMLST | B2 | human/livestock |
| 809 | 1 | PUBMLST | D | human |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saleem, A.F.; Allana, A.; Hale, L.; Diaz, A.; Salinas, R.; Salinas, C.; Qureshi, S.M.; Hotwani, A.; Rahman, N.; Khan, A.; et al. The Gut of Healthy Infants in the Community as a Reservoir of ESBL and Carbapenemase-Producing Bacteria. Antibiotics 2020, 9, 286. https://doi.org/10.3390/antibiotics9060286
Saleem AF, Allana A, Hale L, Diaz A, Salinas R, Salinas C, Qureshi SM, Hotwani A, Rahman N, Khan A, et al. The Gut of Healthy Infants in the Community as a Reservoir of ESBL and Carbapenemase-Producing Bacteria. Antibiotics. 2020; 9(6):286. https://doi.org/10.3390/antibiotics9060286
Chicago/Turabian StyleSaleem, Ali F., Ahreen Allana, Lauren Hale, Alondra Diaz, Raul Salinas, Cristina Salinas, Shahida M. Qureshi, Aneeta Hotwani, Najeeb Rahman, Asia Khan, and et al. 2020. "The Gut of Healthy Infants in the Community as a Reservoir of ESBL and Carbapenemase-Producing Bacteria" Antibiotics 9, no. 6: 286. https://doi.org/10.3390/antibiotics9060286
APA StyleSaleem, A. F., Allana, A., Hale, L., Diaz, A., Salinas, R., Salinas, C., Qureshi, S. M., Hotwani, A., Rahman, N., Khan, A., Zaidi, A. K., Seed, P. C., & Arshad, M. (2020). The Gut of Healthy Infants in the Community as a Reservoir of ESBL and Carbapenemase-Producing Bacteria. Antibiotics, 9(6), 286. https://doi.org/10.3390/antibiotics9060286