Abundant Extractable Metabolites from Temperate Tree Barks: The Specific Antimicrobial Activity of Prunus Avium Extracts

 , , , ,
, , , ,  , and
, and
Abstract
1. Introduction
2. Results
2.1. Antimicrobial Screening of Ten Methanol Bark Extracts
2.2. Microbicidal Activities of the Three Bark Extracts Exhibiting the Lowest MICs
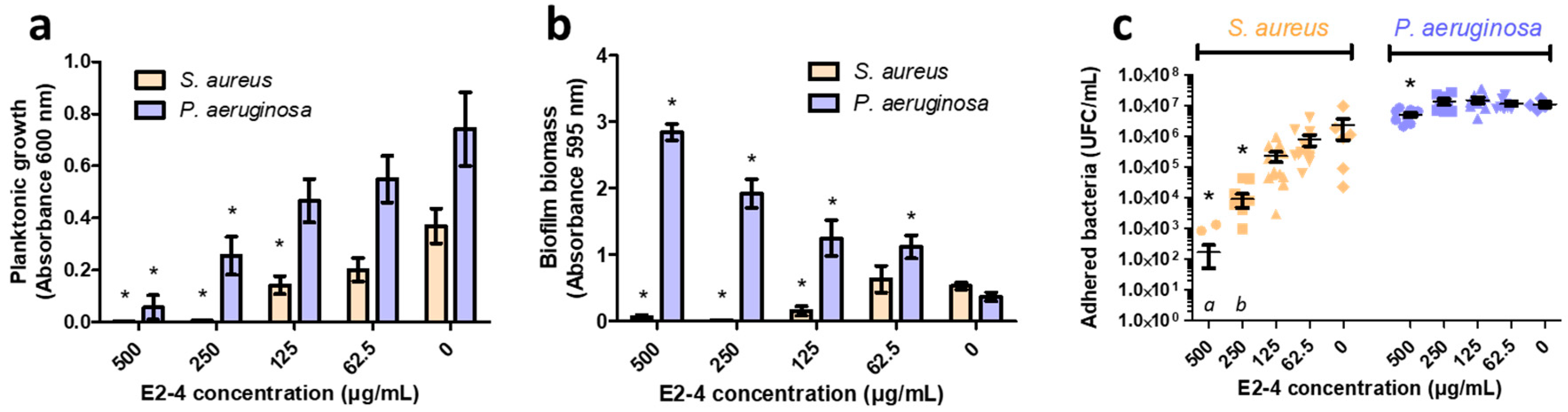
2.3. Evaluation of Antibiofilm Activity of the Prunus Avium Extract (E2-4)
2.4. Chemical Profiling of E2-4
2.5. Antimicrobial and Antibiofilm Activity of Three Fractions from E2-4
3. Discussion
4. Materials and Methods
4.1. Chemicals, Reagents, and Bark Materials
4.2. Microorganisms and Culture Media
4.3. Antimicrobial Screening of the Ten Methanol Bark Extracts
4.4. Static Biofilm Assays
4.5. Centrifugal Partition Chromatography (CPC)
4.6. NMR Analyses, Principal Component Analysis and Dereplication of the Major Potentially Active Compounds
4.7. Statiscal Methods
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- May, M. Drug development: Time for teamwork. Nature 2014, 509, S4–S5. [Google Scholar] [CrossRef] [PubMed]
- Theuretzbacher, U. Antibiotic innovation for future public health needs. Clin. Microbiol. Infect. 2017, 23, 713–717. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Priority List of ANTIBIOTIC-resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics; World Health Organization: Geneva, Switzerland, 2017; Available online: http://www.who.int/medicines/publications/WHO-PPL-Short_Summary_25Feb-ET_NM_WHO.pdf?ua=1 (accessed on 31 January 2020).
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014; Available online: https://www.who.int/drugresistance/documents/surveillancereport/en/ (accessed on 17 February 2020).
- Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States. 2019. Available online: https://www.cdc.gov/drugresistance/pdf/threats-report/2019-ar-threats-report-508.pdf (accessed on 17 February 2020).
- Wright, G.D. Opportunities for natural products in 21st century antibiotic discovery. Nat. Prod. Rep. 2017, 34, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Genilloud, O. The re-emerging role of microbial natural products in antibiotic discovery. Antonie Van Leeuwenhoek 2014, 106, 173–188. [Google Scholar] [CrossRef]
- Kirst, H.A. Developing new antibacterials through natural product research. Expert Opin. Drug Discov. 2013, 8, 479–493. [Google Scholar] [CrossRef]
- Frederich, M.; Marcowycz, A.; Cieckiewicz, E.; Mégalizzi, V.; Angenot, L.; Kiss, R. In Vitro anticancer potential of tree extracts from the Walloon Region forest. Planta Med. 2009, 75, 1634–1637. [Google Scholar] [CrossRef]
- Zofou, D.; Ntie-Kang, F.; Sippl, W.; Efange, S.M. Bioactive natural products derived from the Central African flora against neglected tropical diseases and HIV. Nat. Prod. Rep. 2013, 30, 1098–1120. [Google Scholar] [CrossRef]
- Krol, S.K.; Kiełbus, M.; Rivero-Müller, A.; Stepulak, A. Comprehensive review on betulin as a potent anticancer agent. BioMed Res. Int. 2015, 584189. [Google Scholar] [CrossRef]
- Abedini, A.; Chollet, S.; Angelis, A.; Borie, N.; Nuzillard, J.M.; Skaltsounis, A.L.; Reynaud, R.; Gangloff, S.C.; Renault, J.H.; Hubert, J. Bioactivity-guided identification of antimicrobial metabolites in Alnus glutinosa bark and optimization of oregonin purification by Centrifugal Partition Chromatography. J. Chromatogr. B 2016, 1029, 121–127. [Google Scholar] [CrossRef]
- Elansary, H.; Szopa, A.; Kubica, P.; Ekiert, H.; A Mattar, M.; Al-Yafrasi, M.A.; El-Ansary, D.O.; Zin El-Abedin, T.K.; Yessoufou, K. Polyphenol profile and pharmaceutical potential of Quercus spp. bark extracts. Plants 2019, 8, 486. [Google Scholar] [CrossRef]
- Carbomelle, B.; Denis, F.; Mannonier, A.; Pinon, G.; Vague, R. Bacteriologie Medicale: Techniques Usuelles; SIMEP: Paris, France, 1987; pp. 228–282. [Google Scholar]
- Salem, M.Z.M.; Elansary, H.O.; Elkelish, A.A.; Zeidler, A.; Ali, H.M.; EL-Hefny, M.; Yessoufou, K. In vitro bioactivity and Antimicrobial Activity of Picea abies and Larix decidua Wood and Bark Extracts. BioResources 2016, 11, 9421–9437. [Google Scholar] [CrossRef]
- Drózdz, P.; Pyrzynska, K. Assessment of polyphenol content and antioxidant activity of oak bark extracts. Eur. J. Wood Wood Prod. 2018, 76, 793–795. [Google Scholar] [CrossRef]
- Hubert, J.; Angelis, A.; Aligiannis, N.; Rosalia, M.; Abedini, A.; Bakiri, A.; Reynaud, R.; Nuzillard, J.M.; Gangloff, S.; Skaltsounis, A.L.; et al. Potential dermo-cosmetic use of barks from common deciduous and coniferous trees growing in temperate areas. Planta Med. 2016, 82, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Lynskey, N.N.; Lawrenson, R.A.; Sriskandan, S. New understandings in Streptococcus pyogenes. Curr. Opin. Infect. Dis. 2011, 24, 196–202. [Google Scholar] [CrossRef]
- Cunningham, M.W. Pathogenesis of group A streptococcal infections. Clin. Microbiol. Rev. 2000, 13, 470–511. [Google Scholar] [CrossRef]
- Brook, I. Penicillin Failure in the Treatment of Streptococcal Pharyngo-Tonsillitis. Cur. Infect. Dis. Rep. 2013, 15, 232–235. [Google Scholar] [CrossRef]
- Abuelsaad, A.S.; Mohamed, I.; Allam, G.; Al-Solumani, A.A. Antimicrobial and immunomodulating activities of hesperidin and ellagic acid against diarrheic Aeromonas hydrophila in a murine model. Life Sci. 2013, 93, 714–722. [Google Scholar] [CrossRef]
- Miyasaki, Y.; Rabenstein, J.D.; Rhea, J.; Crouch, M.L.; Mocek, U.M.; Kittell, P.E.; Morgan, M.A.; Nichols, W.S.; Van Benschoten, M.M.; Hardy, W.D.; et al. Isolation and characterization of antimicrobial compounds in plant extracts against multidrug-resistant Acinetobacter baumannii. PLoS ONE 2013, 8, e61594. [Google Scholar] [CrossRef]
- Li, Z.J.; Guo, X.; Dawuti, G.; Aibai, S. Antifungal activity of ellagic acid in vitro and in vivo. Phytother. Res. 2015, 29, 1019–1025. [Google Scholar] [CrossRef]
- Mo, Y.Y.; Geibel, M.; Bonsall, R.F.; Gross, D.C. Analysis of sweet cherry (Prunus avium L.) leaves for plant signal molecules that activate the syrB gene required for synthesis of the phytotoxin, syringomycin, by Pseudomonas syringae pv syringae. Plant Physiol. 1995, 107, 603–612. [Google Scholar] [CrossRef]
- De la Fuente-Núñez, C.; Reffuveille, F.; Fernández, L.; Hancock, R.E. Bacterial biofilm development as a multicellular adaptation: Antibiotic resistance and new therapeutic strategies. Curr. Opin. Microbiol. 2013, 16, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Reffuveille, F.; De La Fuente-Núñnez, C.; Mansour, S.; Hancock, R.E. A broad-spectrum antibiofilm peptide enhances antibiotic action against bacterial biofilms. Antimicrob. Agents Chemother. 2014, 58, 5363–5371. [Google Scholar] [CrossRef] [PubMed]
- Shi, N.; Gao, Y.; Yin, D.; Song, Y.; Kang, J.; Li, X.; Zhang, Z.; Feng, X.; Duan, J. The effect of the sub-minimal inhibitory concentration and the concentrations within resistant mutation window of ciprofloxacin on MIC, swimming motility and biofilm formation of Pseudomonas aeruginosa. Microb. Pathog. 2019, 137, 103765. [Google Scholar] [CrossRef] [PubMed]
- Ancuceanu, R.; Dinu, M.; Dinu-Pirvu, C.; Anuţa, V.; Negulescu, V. Pharmacokinetics of B-Ring Unsubstituted Flavones. Pharmaceutics 2019, 11, 370. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically M07-A10, 10th ed.; Approved Standard Document M7; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015; Available online: http://www.facm.ucl.ac.be/intranet/CLSI/CLSI-2015-M07-A10-original.pdf (accessed on 31 January 2020).
- Belhadj-Mostefa, M.; Abedini, A.; Gangloff, S.C.; Kabouche, A.; Kabouche, Z.; Voutquenne-Nazabadioko, L. A new delta-tocotrienolic acid derivative and other constituents from the cones of Cedrus atlantica and their in vitro antimicrobial activity. Phytochem. Lett. 2017, 20, 252–258. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Extract Code | Binomial Name | Common Name |
|---|---|---|
| E2-1 | Fagus sylvatica | Common beech |
| E2-2 | Quercus robur | Pedunculate oak |
| E2-3 | Alnus glutinosa | Black alder |
| E2-4 | Prunus avium | Wild cherry |
| E2-5 | Acer pseudoplatanus | Sycamore maple |
| E2-6 | Fraxinus excelsior | European ash |
| E2-7 | Populus x canadensis (Robusta) | Canadian poplar |
| E2-8 | Larix decidua | European larch |
| E2-9 | Picea abies | Norway spruce |
| E2-10 | Populus tremula | Eurasian aspen |
| Micro-Organism | MIC (mg/mL) | Positive Controls | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E2-1 | E2-2 | E2-3 | E2-4 | E2-5 | E2-6 | E2-7 | E2-8 | E2-9 | E2-10 | G | V | A | |||
| Gram positive bacteria | Bacillus subtilis | 5 | ≤0.3 | ≤0.3 | ≤0.3 | 5 | 1.2 | 2.5 | 1.2 | 1.2 | 2.5 | S | S | NT | |
| Enterococcus faecalis ATCC 1034 | 10 | ≤0.3 | ≤0.3 | ≤0.3 | NA | 5 | 5 | 1.2 | 2.5 | 5 | R | R | NT | ||
| Staphylococcus aureus NCTC 8325 | 0.6 | ≤0.3 | ≤0.3 | ≤0.3 | NA | 2.5 | 2.5 | ≤0.3 | 1.2 | 5 | S | S | NT | ||
| Staphylococcus aureus CIP 53.154 | ≤0.3 | ≤0.3 | ≤0.3 | ≤0.3 | NA | 1.2 | 2.5 | 1.2 | 2.5 | 2.5 | S | S | NT | ||
| Staphylococcus epidermidis | 0.6 | ≤0.3 | ≤0.3 | ≤0.3 | 10 | 1.2 | 2.5 | ≤0.3 | ≤0.3 | 1.2 | S | S | NT | ||
| Listeria innocua | 5 | ≤0.3 | ≤0.3 | ≤0.3 | 5 | 2.5 | 5 | 1.2 | 1.2 | 2.5 | S | S | NT | ||
| Streptococcus pyogenes | 5 | ≤0.3 | ≤0.3 | ≤0.3 | ≤0.3 | ≤0.3 | 2.5 | ≤0.3 | ≤0.3 | ≤0.3 | S | S | NT | ||
| Micrococcus luteus | 1.2 | ≤0.3 | ≤0.3 | ≤0.3 | 5 | 2.5 | 2.5 | ≤0.3 | ≤0.3 | 2.5 | S | S | NT | ||
| Gram negative bacteria | Escherichia coli CIP 54.127 | NA | 2.5 | 0.6 | 10 | NA | NA | NA | NA | NA | NA | S | R | NT | |
| Enterobacter cloacae | NA | 2.5 | 0.6 | 10 | NA | NA | NA | NA | NA | NA | S | R | NT | ||
| Salmonella enterica | NA | 1.2 | 0.6 | 10 | NA | NA | NA | NA | NA | NA | S | R | NT | ||
| Serratia marcescens | NA | 1.2 | ≤0.3 | NA | NA | NA | NA | NA | NA | NA | S | R | NT | ||
| Proteus vulgaris | NA | ≤0.3 | ≤0.3 | 2.5 | NA | NA | NA | NA | NA | NA | S | R | NT | ||
| Klebsiella pneumoniae | NA | 2.5 | 0.6 | NA | NA | NA | NA | NA | NA | NA | R | R | NT | ||
| Providencia stuartii | NA | ≤0.3 | ≤0.3 | 2.5 | NA | NA | NA | NA | NA | NA | S | R | NT | ||
| Pseudomonas aeruginosa ATCC 9027 | NA | ≤0.3 | ≤0.3 | 10 | NA | NA | NA | NA | NA | NA | S | R | NT | ||
| Shigella sonnei | 0.6 | ≤0.3 | ≤0.3 | ≤0.3 | 10 | 1.2 | 2.5 | ≤0.3 | 0.6 | 1.2 | S | I | NT | ||
| Yeast | Candida glabrata | NA | ≤0.3 | ≤0.3 | ≤0.3 | NA | NA | NA | 10 | NA | NA | R | R | S | |
| Candida tropicalis | NA | ≤0.3 | 0.6 | ≤0.3 | NA | NA | NA | ≤0.3 | NA | NA | R | R | S | ||
| Candida kefyr | NA | ≤0.3 | ≤0.3 | ≤0.3 | NA | NA | NA | NA | NA | NA | R | R | S | ||
| Candida albicans | NA | ≤0.3 | 0.6 | ≤0.3 | NA | NA | NA | ≤0.3 | NA | NA | R | R | S | ||
| Cryptococcus neoformans | 2.5 | ≤0.3 | ≤0.3 | ≤0.3 | 2.5 | NA | NA | NA | NA | NA | R | R | S | ||
| Micro-Organism | µg/mL | ||||||
|---|---|---|---|---|---|---|---|
| E2-2 | E2-3 | E2-4 | |||||
| MIC | MBC | MIC | MBC | MIC | MBC | ||
| Gram positive bacteria | Bacillus subtilis | 62.5 | NA | 125 | NA | 62.5 | 125 |
| Enterococcus faecalis ATCC 1034 | 62.5 | NA | 125 | NA | 62.5 | 250 | |
| Staphylococcus aureus NCTC 8325 | 62.5 | 250 | 62.5 | 250 | 62.5 | 125 | |
| Staphylococcus aureus CIP 53.154 | 31.2 | NA | 62.5 | NA | 62.5 | 125 | |
| Staphylococcus epidermidis | 31.2 | NA | 62.5 | NA | 62.5 | NA | |
| Listeria innocua | 31.2 | 125 | 125 | 250 | 62.5 | 125 | |
| Streptococcus pyogenes | 31.2 | NA | 7.8 | NA | 31.2 | NA | |
| Micrococcus luteus | 31.2 | 62.5 | 62.5 | NA | 62.5 | 250 | |
| Gram negative bacteria | Serratia marcescens | - | - | 62.5 | NA | - | - |
| Proteus vulgaris | 62.5 | NA | 31.2 | NA | - | - | |
| Providencia stuartii | 15.6 | NA | 125 | NA | - | - | |
| Pseudomonas aeruginosa ATCC 9027 | 31.2 | NA | 125 | 125 | - | - | |
| Shigella sonnei | 15.6 | NA | 250 | NA | 125 | NA | |
| MIC | MFC | MIC | MFC | MIC | MFC | ||
| Yeast | Candida glabrata | 31.2 | 125 | 125 | NA | 125 | NA |
| Candida tropicalis | 31.2 | NA | - | - | 31.2 | 250 | |
| Candida kefyr | 62.5 | 250 | 125 | NA | 62.5 | 250 | |
| Candida albicans | 62.5 | 125 | - | - | 125 | 125 | |
| Cryptococcus neoformans | 125 | NA | 125 | NA | 62.5 | NA | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abedini, A.; Colin, M.; Hubert, J.; Charpentier, E.; Angelis, A.; Bounasri, H.; Bertaux, B.; Kotland, A.; Reffuveille, F.; Nuzillard, J.-M.; et al. Abundant Extractable Metabolites from Temperate Tree Barks: The Specific Antimicrobial Activity of Prunus Avium Extracts. Antibiotics 2020, 9, 111. https://doi.org/10.3390/antibiotics9030111
Abedini A, Colin M, Hubert J, Charpentier E, Angelis A, Bounasri H, Bertaux B, Kotland A, Reffuveille F, Nuzillard J-M, et al. Abundant Extractable Metabolites from Temperate Tree Barks: The Specific Antimicrobial Activity of Prunus Avium Extracts. Antibiotics. 2020; 9(3):111. https://doi.org/10.3390/antibiotics9030111
Chicago/Turabian StyleAbedini, Amin, Marius Colin, Jane Hubert, Emilie Charpentier, Apostolis Angelis, Heithem Bounasri, Benjamin Bertaux, Alexis Kotland, Fany Reffuveille, Jean-Marc Nuzillard, and et al. 2020. "Abundant Extractable Metabolites from Temperate Tree Barks: The Specific Antimicrobial Activity of Prunus Avium Extracts" Antibiotics 9, no. 3: 111. https://doi.org/10.3390/antibiotics9030111
APA StyleAbedini, A., Colin, M., Hubert, J., Charpentier, E., Angelis, A., Bounasri, H., Bertaux, B., Kotland, A., Reffuveille, F., Nuzillard, J.-M., Renault, J.-H., & Gangloff, S. C. (2020). Abundant Extractable Metabolites from Temperate Tree Barks: The Specific Antimicrobial Activity of Prunus Avium Extracts. Antibiotics, 9(3), 111. https://doi.org/10.3390/antibiotics9030111