Antimicrobial Secretions of Toads (Anura, Bufonidae): Bioactive Extracts and Isolated Compounds against Human Pathogens

 ,
,  , and
, and
Abstract
1. Introduction (True Toads, Antimicrobial Resistance of Human Pathogens against Drugs)

2. Methodology
3. Antimicrobial Activities
3.1. Antibacterial

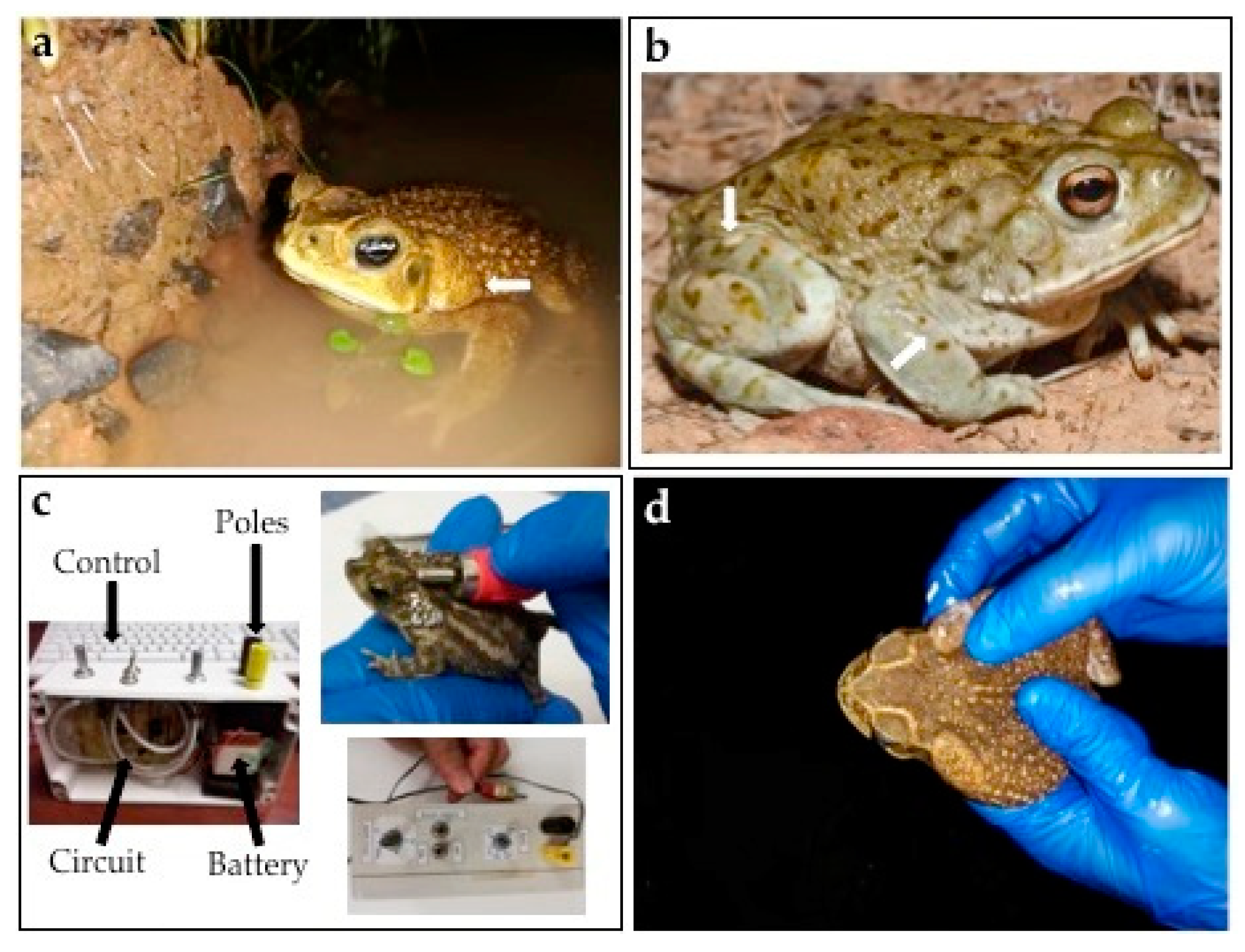{kind=link}
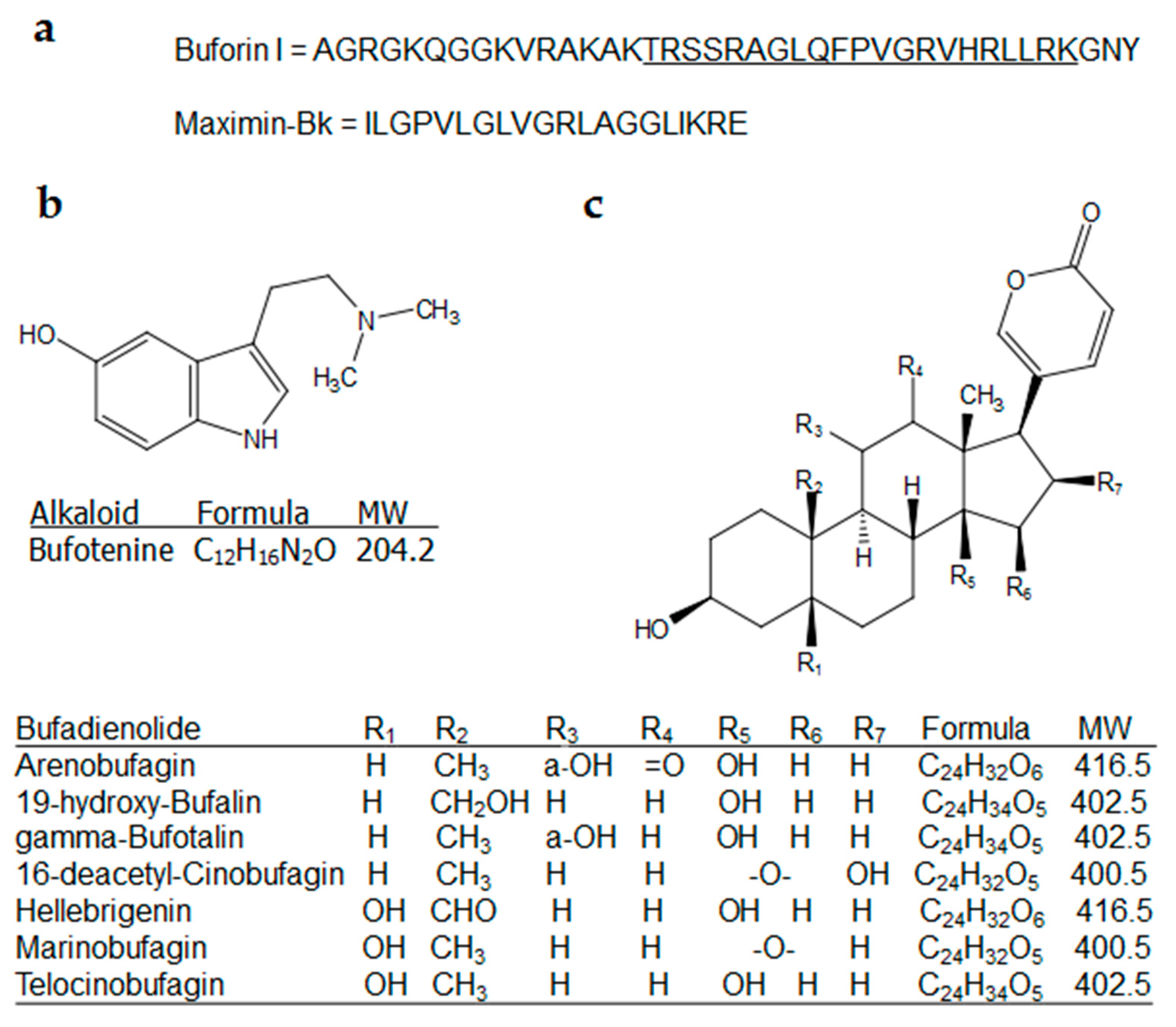{kind=link}
| Toad Species a | Sample (Solvent Used) | Effect | Bioactivity b,c,d | Toxicity (μg/mL) d,e | Reference |
|---|---|---|---|---|---|
| Bufo bufo | Skin gland + parotoid gland secretion (ultra-pure water) | Antibacterial | MIC (μg/mL): 250 (S. thyphimirium); 62.5 (S. aureus); 3.9 (E. Faecalis, E. faecium, S. epidermidis) | IC50: 0.35 (HEK-293) | [52] |
| Antifungal | MIC (μg/mL): 250 (C. albicans) | ||||
| Bufo gargarizans (Bufo andrewsi) | Ba-lysozyme | Antibacterial | MIC (μM): 8 (E. coli); 1 (S. aureus) | * | [53] |
| Bufo gargarizans | Buforin-I | Antibacterial | MIC (μg/mL): 8 (E. coli, Serratia sp., S. mutans); 4 (B. subtilis, P. putida, S. thyphimurium, S. aureus, S. pneumoniae) | * | [47] |
| Antifungal | MIC (μg/mL): 4 (C. albicans, C. neoformans) | ||||
| Bufo verrucosissimus | Skin gland + parotoid gland secretion (ultra-pure water) | Antibacterial | MIC (μg/mL): 62.5 (S. aureus); 3.9 (E. faecalis, E. faecium, S. epidermidis) | IC50: 0.99 (HEK-293) | [52] |
| Antifungal | MIC (μg/mL): 125 (C. albicans) | ||||
| Bufotes sitibundus (Bufo kavirensis) | Maximin-Bk | Antibacterial | MIC (μg/mL): 20.78 (L. mesenteroides); 19.4 (B. subtilis); 18.5 (B. cereus); 16.3 (S. aureus); 10.3 (P. aeruginosa); 8.9 (K. pneumoniae); 8.1 (E. coli) | * | [50] |
| Antifungal | MIC (μg/mL): 35.6 (A. fumigates); 32.1 (C. albicans); 28.6 (A. niger); 25.7 (P. lilacinum) | ||||
| Bufotes sitibundus (Bufotes variabilis) | Skin gland + parotoid gland secretion (ultra-pure water) | Antibacterial | MIC (μg/mL): 125 (S. aureus, S. thyphimirium); 7.8 (S. epidermidis); 3.9 (E. faecalis, E. faecium) | IC50: 1.46 (HEK-293) | [52] |
| Antifungal | MIC (μg/mL): 125 (C. albicans) | ||||
| Duttaphrynus melanostictus (Bufo melanostictus) | Granular gland secretion (0.9% NaCl) | Antifungal | Halo zones (7.5% w/v): C. albicans (20.3 mm); M. gypsum (24.5 mm); T. mentagrophytes (24.7 mm) | * | [59] |
| Parotoid gland secretion (distilled water) | Antibacterial | Halo zones: E.coli (7 mm); S. typhimurium (7 mm); S. epidermidis (8 mm); K. pneumoniae (9 mm); B. cereus (10 mm); S. aureus (11 mm) | * | [43] | |
| Skin (0.9% NaCl) | Antifungal | Halo zones: P. notatum (21 mm); A. niger (23 mm) | * | [60] | |
| Antibacterial | Halo zones: E. coli (19 mm); K. pneumoniae (24 mm); S. aureus (27 mm); P. vulgaris (33 mm) | * | [44] | ||
| Skin gland secretion (distilled water) | Antibacterial | Halo zones: K. pneumoniae (19 mm); E. coli (25 mm); P. vulgaris (28 mm); S. aureus (30 mm) | * | [44] | |
| Antifungal | Halo zones: P. notatum (23 mm); A. niger (25 mm) | * | [60] | ||
| Leptophryne cruentata | Skin gland secretion (acetate buffer) | Antifungal | Halo zone: T. mentagrophytes (14.5 mm) | * | [61] |
| Phrynoidis asper (Bufo asper) | Skin secretion (deionized water) | Antibacterial | MIC (μg/mL): 100 (E. coli); 50 (B. cereus, K. pneumoniae, P. aeruginosa); 25 (B. subtilis); 12 (S. aureus) | * | [46] |
| Rhaebo guttatus | Skin gland secretion (CHCl3/MeOH) | Antiparasitic | IC50 (μg/mL): 0.05 (P. falciparum, ring) | LD50: 34.83 (BGM) | [62] |
| Rhinella arenarum (Bufo arenarum) | Venom (distilled water) | Antibacterial | MIC (μg/mL): >1250 (A. baumannii, B. subtilis); 1250 (E. coli, K. pneumoniae, S. aureus); 625 (A. hydrophila); 312.5 (P. aeruginosa) | * | [63] |
| Antifungal | Inactive for C. albicans and A. niger | ||||
| LBP-1 | Antibacterial | Halo zones (25 μg): E. coli (16 mm); E. faecalis (12 mm); P. morganii (20 mm) | * | [51] | |
| LBP-2 | Antibacterial | Halo zones (25 μg): E. coli (17.5 mm); E. faecalis (12.5 mm); P. morganii (19 mm) | |||
| Rhinella centralis | 19-Hydroxy-bufalin | Antiparasitic | IC50 (μg/mL): 7.81 (T. cruzi, tryp) | IC50: 71.58 (Vero) | [64] |
| Rhinella icterica | Parotoid gland secretion | Antibacterial | medium inhibition at 25 mg/mL for E. coli; S. aureus | * | [54] |
| Rhinella jimi | Parotoid gland secretion (EtOH) | Antibacterial | MIC (μg/mL): ≥2048 (E. coli; P. aeruginosa; S. aureus) | LD50: 365.94 (shrimp) | [45] |
| Hellebrigenin | Antiparasitic | IC50 (μg/mL): 126.2 (L. chagasi, prom); 91.75 (T. cruzi, tryp) | IC50 > 200 (Macrophages) | [65] | |
| Telocinobufagin | Antiparasitic | IC50 (μg/mL): 61.2 (L. chagasi, prom) | |||
| Rhinella marina | Skin gland secretion (crude) | Antibacterial | MIC (μg/mL): >25 (E. coli); 21 (S. aureus); 10.79 (P. aeruginosa) | IC50 > 100 (MRC5) | [66] |
| Antiparasitic | IC50 (μg/mL): 14.82 (L. braziliensis, prom); 9.34 (L. guyanensis, prom); 2.43 (P. falciparum, ring) | ||||
| Skin gland secretion (MeOH) | Antiparasitic | MIC (μg/mL): ≥100 (L. braziliensis, prom); 12.04 (P. falciparum, ring); 3.99 (L. guyanensis, prom) | * | [66] | |
| Skin gland secretion (CHCl3/MeOH) | Antiparasitic | IC50: 0.534 (P. falciparium, ring) | LD50 > 200 (BGM) | [62] | |
| 16-Desacetil-cinobufagin | Antibacterial | MIC (μg/mL) < 3.12 (E. coli; P. aeruginosa; S. aureus) | IC50 > 100 (MRC5) | [66] | |
| Marinobufagin | Antibacterial | MIC (μg/mL) < 3.12 (S. aureus) | IC50 > 100 (MRC5) | [66] | |
| Telocinobufagin | Antiparasitic | IC50 (μg/mL): 1.28 (P. falciparum, ring) | IC50 > 200 (BGM) | [62] | |
| Rhinella rubescens (Bufo rubescens) | Marinobufagin | Antibacterial | MIC (μg/mL): 128 (S. aureus); 16 (E. coli) | * | [57] |
| Telocinobufagin | Antibacterial | MIC (μg/mL): 128 (S. aureus); 64 (E. coli) | |||
| Sclerophrys pantherina (Amietophrynus pantherinus) | Skin gland secretion (phosphate buffer) | Antifungal | MIC (mg/mL): 0.39 (F. verticillioides); 0.04 (C. albicans); 0.02 (A. flavus) | * | [67] |
3.2. Antifungal
3.3. Antiprotozoal
4. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- AmphibiaWeb. Available online: amphibiaweb.org (accessed on 29 September 2020).
- Amphibian Species of the World. Available online: https://amphibiansoftheworld.amnhorg/Amphibia/Anura/Bufonidae (accessed on 29 September 2020).
- Ódonohoe, M.E.A.; Luna, M.C.; Regueira, E.; Brunetti, A.E.; Basso, N.G.; Lynch, J.D.; Pereyra, M.O.; Hermida, G.N. Diversity and evolution of the parotoid macrogland in true toads (Anura: Bufonidae). Zool. J. Linn. Soc. 2019, 187, 453–478. [Google Scholar] [CrossRef]
- Rodriguez, C.; Rollins-Smith, L.; Ibáñez, R.; Durant-Archibold, A.A.; Gutiérrez, M. Toxins and pharmacologically active compounds from species of the family Bufonidae (Amphibia, Anura). J. Ethnopharmacol. 2017, 198, 235–254. [Google Scholar] [CrossRef] [PubMed]
- Clarke, B.T. The natural history of amphibian skin secretions, their normal functioning and potential medical applications. Biol. Rev. 1997, 72, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Toledo, R.C.; Jared, C. Cutaneous granular glands and amphibian venoms. Comp. Biochem. Physiol. Part A Physiol. 1995, 111, 1–29. [Google Scholar] [CrossRef]
- Mailho-Fontana, P.L.; Antoniazzi, M.M.; Toledo, L.F.; Verdade, V.K.; Sciani, J.M.; Barbaro, K.C.; Pimenta, D.C.; Rodrigues, M.T.; Jared, C. Passive and active defense in toads: The parotoid macroglands in Rhinella marina and Rhaebo guttatus. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2014, 321, 65–77. [Google Scholar] [CrossRef]
- Barnhart, K.; Forman, M.E.; Umile, T.P.; Kueneman, J.; McKenzie, V.; Salinas, I.; Minbiole, K.P.C.; Woodhams, D.C. Identification of bufadienolides from the boreal toad, Anaxyrus boreas, active against a fungal pathogen. Microb. Ecol. 2017, 74, 990–1000. [Google Scholar] [CrossRef]
- Sun, T.; Zhan, B.; Gao, Y. A novel cathelicidin from Bufo bufo gargarizans Cantor showed specific activity to its habitat bacteria. Gene 2015, 571, 172–177. [Google Scholar] [CrossRef]
- Qi, J.; Zulfiker, A.H.M.; Li, C.; Good, D.; Wei, M.Q. The development of toad toxins as potential therapeutic agents. Toxins 2018, 10, 336. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/results?cond=&term=cinobufacini&cntry=&state=&city=&dist= (accessed on 30 September 2020).
- Riós-Orjuela, J.C.; Falcón-Espitia, N.; Arias-Escobar, A.; Espejo-Uribe, M.J.; Chamorro-Vargas, C. Knowledge and interactions of the local community with the herpetofauna in the forest reserve of Quininí (Tibacuy-Cundinamarca, Colombia). J. Ethnobiol. Ethnomed. 2020, 16, 1–11. [Google Scholar] [CrossRef]
- Barros, F.B.; Varela, S.A.M.; Pereira, H.M.; Vicente, L. Medicinal use of fauna by a traditional community in the Brazilian Amazonia. J. Ethnobiol. Ethnomed. 2012, 8, 37. [Google Scholar] [CrossRef]
- Ferreira, F.S.; Albuquerque, U.P.; Melo Coutinho, H.D.; Almeida, W.D.O.; Nóbrega Alves, R.R. The trade in medicinal animals in northeastern Brazil. Evid.-Based Complement. Altern. Med. 2012, 2012, 126938. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.B.; Land, B. Transcutaneous Amphibian Stimulator (TAS): A device for the collection of amphibian skin secretions. Herpetol. Rev. 2002, 33, 38–41. [Google Scholar]
- Koo, H.B.; Seo, J. Antimicrobial peptides under clinical investigation. Pept. Sci. 2019, 111, 24122. [Google Scholar] [CrossRef]
- Xu, X.; Lai, R. The chemistry and biological activities of peptides from amphibian skin secretions. Chem. Rev. 2015, 115, 1760–1846. [Google Scholar] [CrossRef] [PubMed]
- Huo, Y.; Xv, R.; Ma, H.; Zhou, J.; Xi, X.; Wu, Q.; Duan, J.; Zhou, M.; Chen, T. Identification of <10 KDa peptides in the water extraction of Venenum Bufonis from Bufo gargarizans using Nano LC-MS/MS and De novo sequencing. J. Pharm. Biomed. Anal. 2018, 157, 156–164. [Google Scholar] [PubMed]
- Yumi, P.; Shibao, T.; Cologna, C.T.; Morandi-Filho, R.; Wiezel, G.A.; Fujimura, P.T.; Ueira-Viera, C.; Arantes, E.C. Deep sequencing analysis of toad Rhinella schneideri skin glands and partial biochemical characterization of its cutaneous secretion. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 1–15. [Google Scholar]
- Dailami, M.; Artika, I.M.; Kusrini, M.D. Analysis and prediction of some histone-derived antimicrobial peptides from toads Duttaphrynus melanostictus and Phyrinoidis asper. J. Pure Appl. Chem. Res. 2016, 5, 67–76. [Google Scholar] [CrossRef][Green Version]
- World Health Organization (WHO). Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 6 September 2020).
- Centers for Disease Control (CDC)—Fungal Diseases. Available online: https://www.cdc.gov/fungal/index.html (accessed on 15 September 2020).
- Centers for Disease Control (CDC)—Parasites. Available online: https://www.cdc.gov/parasites/index.html (accessed on 15 September 2020).
- Morse, S.S. Factors in the emergence of infectious diseases. Emerg. Infect. Dis. 1995, 1, 7–15. [Google Scholar] [CrossRef]
- Smolinski, M.; Hamburg, M.A.; Lederberg, J. Microbial Threats to Health; National Academies Press: Cambridge, MA, USA, 2003. [Google Scholar]
- Wang, Z.A. Literature review of bacterial drug resistance. Mater. Sci. Forum 2020, 980, 197–209. [Google Scholar] [CrossRef]
- Yoneyama, H.; Nakae, T. Mechanism of efficient elimination of protein D2 in outer membrane of imipenem-resistant Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1993, 37, 2385–2390. [Google Scholar] [CrossRef]
- Ducret, V.; Gonzalez, M.R.; Leoni, S.; Valentini, M.; Perron, K. The CzcCBA efflux system requires the CadA P-type ATPase for timely expression upon zinc excess in Pseudomonas aeruginosa. Front. Microbiol. 2020, 11, 911. [Google Scholar] [CrossRef]
- Lockart, M.M.; Butler, J.T.; Mize, C.J.; Fair, M.N.; Cruce, A.A.; Conner, K.P.; Atkins, W.M.; Bowman, M.K. Multiple drug binding modes in Mycobacterium tuberculosis CYP51B1. J. Inorg. Biochem. 2020, 205, 110994. [Google Scholar] [CrossRef]
- Shaw, W.V.; Brodsky, R.F. Characterization of chloramphenicol acetyltransferase from chloramphenicol-resistant Staphylococcus aureus. J. Bacteriol. 1968, 95, 28–36. [Google Scholar] [CrossRef]
- Knowles, J.R. Penicillin resistance: The chemistry of β-lactamase inhibition. Acc. Chem. Res. 1985, 18, 97–104. [Google Scholar] [CrossRef]
- Berman, J.; Krysan, D.J. Drug resistance and tolerance in fungi. Nat. Rev. Microbiol. 2020, 18, 319–331. [Google Scholar] [CrossRef]
- Posteraro, B.; Sanguinetti, M.; Sanglard, D.; La Sorda, M.; Boccia, S.; Romano, L.; Morace, G.; Fadda, G. Identification and characterization of a Cryptococcus neoformans ATP binding cassette (ABC) transporter-encoding gene, CnAFR1, involved in the resistance to fluconazole. Mol. Microbiol. 2003, 47, 357–371. [Google Scholar] [CrossRef]
- Katiyar, S.K.; Edlind, T.D. Identification and expression of multidrug resistance-related ABC transporter genes in Candida krusei. Med. Mycol. 2001, 39, 109–116. [Google Scholar] [CrossRef]
- Seo, K.; Akiyoshi, H.; Ohnishi, H. Alteration of cell wall composition leads amphotericin B resistance in Aspergillus flavus. Microbiol. Immunol. 1999, 43, 1017–1025. [Google Scholar] [CrossRef]
- Martel, C.M.; Parker, J.E.; Bader, O.; Weig, M.; Gross, U.; Warrilow, A.G.S.; Rolley, N.; Kelly, D.E.; Kelly, S.L. Identification and characterization of four azole-resistant erg3 mutants of Candida albicans. Antimicrob. Agents Chemother. 2010, 54, 4527–4533. [Google Scholar] [CrossRef]
- Baker, N.; Glover, L.; Munday, J.C.; Andrés, D.A.; Barret, M.P.; De Koning, H.P.; Horn, D. Aquaglyceroporin 2 controls susceptibility to melarsoprol and pentamidine in African trypanosomes. Proc. Natl. Acad. Sci. USA 2012, 109, 10996–11001. [Google Scholar] [CrossRef]
- Ross, L.S.; Dhingra, S.K.; Mok, S.; Yeo, T.; Wicht, K.J.; Kumpornsin, K.; Takala-Harrison, S.; Witkowski, B.; Fairhurst, R.M.; Ariey, F.; et al. Emerging Southeast Asian PfCRT mutations confer Plasmodium falciparum resistance to the first-line antimalarial piperaquine. Nat. Commun. 2018, 9, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Victoria, J.M.; Cortés-Selva, F.; Parodi-Talice, A.; Bavchvarov, B.I.; Pérez-Victoria, F.J.; Muñoz-Martínez, F.; Maitrejean, M.; Costi, M.P.; Barron, D.; Di Pietro, A.; et al. Combination of suboptimal doses of inhibitors targeting different domains of LtrMDR1 efficiently overcomes resistance of Leishmania spp. to miltefosine by inhibiting drug efflux. Antimicrob. Agents Chemother. 2006, 50, 3102–3110. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Adaui, V.; Schnorbusch, K.; Zimic, M.; Gutirrez, A.; Decuypere, S.; Vanaerschot, M.; De Donker, S.; Maes, I.; Llanos-Cuentas, A.; Chappuis, F.; et al. Comparison of gene expression patterns among Leishmania braziliensis clinical isolates showing a different in vitro susceptibility to pentavalent antimony. Parasitology 2011, 138, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Domingo-Calap, P.; Delgado-Martínez, J. Bacteriophages: Protagonists of a post-antibiotic era. Antibiotics 2018, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Holford, B.M.; Daly, M.; King, G.F.; Norton, R.S. Venoms to the rescue. Science 2018, 361, 842–844. [Google Scholar] [CrossRef]
- Syafiq, M.; Zahari, A.; Darnis, D.S.; Haziyamin, T.; Abdul, T. Protein profiles and antimicrobial activity of common Sudan toad, Duttaphrynus melanostictus paratoid secretions. Nat. Preced. 2015. [Google Scholar] [CrossRef]
- Thirupathi, K.; Chandrakala, G.; Krishna, L.; Rao, T.B.; Venkaiah, Y. Antibacterial activity of skin secretion and its extraction from the toad Bufo melanostictus. Eur. J. Pharm. Med. Res. 2018, 3, 283–286. [Google Scholar]
- Sales, D.L.; Bezerra Morais-Braga, M.F.; Lucas Dos Santos, A.T.; Targino Machado, A.J.; De Araujo Filho, J.A.; De Queiroz Dias, D.; Bezerra Da Cunha, F.A.; De Aquino Saraiva, R.; Alencar De Menezes, I.R.; Melo Coutinho, H.D.; et al. Antibacterial, modulatory activity of antibiotics and toxicity from Rhinella jimi (Stevaux, 2002) (Anura: Bufonidae) glandular secretions. Biomed. Pharmacother. 2017, 92, 554–561. [Google Scholar] [CrossRef]
- Dahham, S.S.; Sen Hew, C.; Jaafar, I.; Gam, L.H. The protein profiling of asian giant toad skin secretions and their antimicrobial activity. Int. J. Pharm. Pharm. Sci. 2016, 8, 88–95. [Google Scholar]
- Park, C.B.; Kim, M.S.; Kim, S.C. A novel antimicrobial peptide from Bufo bufo gargarizans. Biochem. Biophys. Res. Commun. 1996, 218, 408–413. [Google Scholar] [CrossRef]
- Park, C.B.; Kim, H.S.; Kim, S.C. Mechanism of action of the antimicrobial peptide buforin II: Buforin II kills microorganisms by penetrating the vell membrane and inhibiting cellular functions. Biochem. Biophys. Res. Commun. 1998, 244, 253–257. [Google Scholar] [CrossRef]
- Park, C.B.; Yi, K.S.; Matsuzaki, K.; Kim, M.S.; Kim, S.C. Structure-activity analysis of buforin II, a histone H2A-derived antimicrobial peptide: The proline hinge is responsible for the cell-penetrating ability of buforin II. Proc. Natl. Acad. Sci. USA 2000, 97, 8245–8250. [Google Scholar] [CrossRef]
- Zare-Zardini, H.; Ebrahimi, L.; Mahdi Ejtehadi, M.; Hashemi, A.; Ghorani Azam, A.; Atefi, A.; Soleimanizadeh, M. Purification and characterization of one novel cationic antimicrobial peptide from skin secretion of Bufo kavirensis. Turk. J. Biochem. 2013, 38, 416–424. [Google Scholar] [CrossRef]
- Sánchez Riera, A.; Daud, A.; Gallo, A.; Genta, S.; Aybar, M.; Sánchez, S. Antibacterial activity of lactose-binding lectins from Bufo arenarum skin. Biocell 2003, 27, 37–46. [Google Scholar] [CrossRef]
- Nalbantsoy, A.; Karis, M.; Tansel Yalcin, H.; Göçmen, B. Biological activities of skin and parotoid gland secretions of bufonid toads (Bufo bufo, Bufo verrucosissimus and Bufotes variabilis) from Turkey. Biomed. Pharmacother. 2016, 80, 298–303. [Google Scholar] [CrossRef]
- Zhao, Y.; Jin, Y.; Lee, W.H.; Zhang, Y. Purification of a lysozyme from skin secretions of Bufo andrewsi. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 142, 46–52. [Google Scholar] [CrossRef]
- Pinto, E.G.; Felipe, A.C.; Nadaletto, D.; Rall, V.L.M.; Martinez, R.M. Research of the antimicrobial activity of the poison from Rhinella icterica (Amphibia, Anura). Rev. Inst. Adolfo Lutz 2009, 68, 471–475. [Google Scholar]
- Cei, J.M.; Erspamer, V.; Roseghini, M. Taxonomic and evolutionary significance of biogenic amines and polypeptides occurring in amphibian skin. II. Toads of the genera Bufo and Melanophryniscus. Syst. Zool. 1968, 17, 232–245. [Google Scholar] [CrossRef]
- Preusser, H.J.; Habermehl, G.; Sablofski, M.; Schmall-Haury, D. Antimicrobial activity of alkaloids from amphibian venoms and effects on the ultrastructure of yeast cells. Toxicon 1975, 13, 285–289. [Google Scholar] [CrossRef]
- Cunha Filho, G.A.; Alberto, C.; Lemos, S.; Castro, M.S.; Ma, M.; Leite, R.S.; Kyaw, C.; Pires, O.R.; Bloch, C.; Ferroni, E. Antimicrobial activity of the bufadienolides marinobufagin and telocinobufagin isolated as major components from skin secretion of the toad Bufo rubescens. Toxicon 2005, 45, 777–782. [Google Scholar] [CrossRef]
- Wu, S.C.; Fu, B.D.; Shen, H.Q.; Yi, P.F.; Zhang, L.Y.; Lv, S.; Guo, X.; Xia, F.; Wu, Y.L.; Wei, X.B. Telocinobufagin enhances the Th1 immune response and protects against Salmonella typhimurium infection. Int. Immunopharmacol. 2015, 25, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Barlian, A.; Anggadiredja, K.; Kusumorini, A. Damage in fungal morphology underlies the antifungal effect of lyophilisate of granular gland secretion from Duttaphrynus melanostictus frog. J. Biol. Sci. 2011, 11, 282–287. [Google Scholar] [CrossRef][Green Version]
- Thirupathi, K.; Shankar, C.; Chandrakala, G.; Krishna, L.; Venkaiah, Y. The antifungal activity of skin secretion and its extract of Indian toad Bufo melanosctictus. Int. J. Environ. Ecol. Fam. Urban. Stud. 2019, 9, 7–12. [Google Scholar]
- Made Artika, I.; Pinontoan, S.; Kusrini, M.D. Antifungal activity of skin secretion of bleeding toad Leptophryne cruentata and javan tree frog Rhacophorus margaritifer. Am. J. Biochem. Biotechnol. 2015, 11, 5–10. [Google Scholar] [CrossRef]
- Banfi, F.F.; Guedes, K.S.; Andrighetti, C.R.; Aguiar, A.C.; Debiasi, B.W.; Noronha, J.C.; Rodrigues, D.J.; Vieira Junior, G.M.; Sanchez Marino, B.A. Antiplasmodial and cytotoxic activities of toad venoms from Southern Amazon, Brazil. Korean J. Parasitol. 2016, 54, 415–421. [Google Scholar] [CrossRef]
- Kalayci, S.; Iyigundogdu, Z.U.; Yazici, M.M.; Asutay, B.A.; Demir, O.; Sahin, F. Evaluation of antimicrobial and antiviral activities of different venoms. Infect. Disord. Targets 2016, 16, 44–53. [Google Scholar] [CrossRef]
- Rodriguez, C.; Ibáñez, R.; NG, M.; Spadafora, C.; Durant-Archibold, A.A.; Gutiérrez, M. 19-Hydroxy-bufalin, a major bufadienolide isolated from the parotoid gland secretions of the Panamanian endemic toad Rhinella centralis (Bufonidae), inhibits the growth of Trypanosoma cruzi. Toxicon 2020, 177, 89–92. [Google Scholar] [CrossRef]
- Tempone, A.G.; Pimenta, D.C.; Lebrun, I.; Sartoreli, P.; Taniwaki, N.M.; De Andrade, H.F.; Antoniazzi, M.M.; Jared, C. Antileishmanial and antitrypanosomal activity of bufadienolides isolated from the toad Rhinella jimi parotoid macrogland secretion. Toxicon 2008, 52, 13–21. [Google Scholar] [CrossRef]
- De Medeiros, D.S.S.; Rego, T.B.; Santos, A.A.; Pontes, A.S.; Moreira-Dill, L.S.; Matos, N.B.; Zuliani, J.P.; Stábeli, R.G.; Teles, C.B.G.; Soares, A.M.; et al. Biochemical and biological profile of parotoid zecretion of the Amazonian Rhinella marina (Anura: Bufonidae). Biomed. Res. Int. 2019, 2019, 292315. [Google Scholar]
- Katerere, D.R.; Dawood, A.; Esterhuyse, A.J.; Vismer, H.F.; Govender, T. Antifungal activity of epithelial secretions from selected frog species of South Africa. Afr. J. Biotechnol. 2013, 12, 6411–6418. [Google Scholar]
- Erspamer, V.; Vitali, T.; Roseghini, M.; Cei, J.M. 5-Methoxy- and 5-Hydroxyindoles in the skin of Bufo alvarius. Biochem. Pharmacol. 1967, 16, 1149–1164. [Google Scholar] [CrossRef]
- Sales, L.D.; Oliveira, O.P.; Santos, C.M.E.; Queiroz, D.D.; Kerntopf, M.R.; Melo, C.H.D.; Martins, C.J.G.; Dias, F.F.R.; Silva, F.F.; Nóbrega, A.R.R.; et al. Chemical identification and evaluation of the antimicrobial activity of fixed oil extracted from Rhinella jimi. Pharm. Biol. 2014, 53, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.E.A.; Araújo, S.G.; Morais, M.I.; Sá, N.P.; Lima, C.M.; Rosa, C.A.; Siqueira, E.P.; Johann, S.; Lima, L.A.R.S. Antifungal and antioxidant activity of fatty acid methyl esters from vegetable oils. An. Acad. Bras. Cienc. 2017, 89, 1671–1681. [Google Scholar] [CrossRef] [PubMed]
- Habermehl, G.G. Antimicrobial activity of amphibian venoms. Stud. Nat. Prod. Chem. 1995, 15, 327–339. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodriguez, C.; Ibáñez, R.; Rollins-Smith, L.A.; Gutiérrez, M.; Durant-Archibold, A.A. Antimicrobial Secretions of Toads (Anura, Bufonidae): Bioactive Extracts and Isolated Compounds against Human Pathogens. Antibiotics 2020, 9, 843. https://doi.org/10.3390/antibiotics9120843
Rodriguez C, Ibáñez R, Rollins-Smith LA, Gutiérrez M, Durant-Archibold AA. Antimicrobial Secretions of Toads (Anura, Bufonidae): Bioactive Extracts and Isolated Compounds against Human Pathogens. Antibiotics. 2020; 9(12):843. https://doi.org/10.3390/antibiotics9120843
Chicago/Turabian StyleRodriguez, Candelario, Roberto Ibáñez, Louise A. Rollins-Smith, Marcelino Gutiérrez, and Armando A. Durant-Archibold. 2020. "Antimicrobial Secretions of Toads (Anura, Bufonidae): Bioactive Extracts and Isolated Compounds against Human Pathogens" Antibiotics 9, no. 12: 843. https://doi.org/10.3390/antibiotics9120843
APA StyleRodriguez, C., Ibáñez, R., Rollins-Smith, L. A., Gutiérrez, M., & Durant-Archibold, A. A. (2020). Antimicrobial Secretions of Toads (Anura, Bufonidae): Bioactive Extracts and Isolated Compounds against Human Pathogens. Antibiotics, 9(12), 843. https://doi.org/10.3390/antibiotics9120843