Peptidomic Analysis of Skin Secretions of the Caribbean Frogs Leptodactylus insularum and Leptodactylus nesiotus (Leptodactylidae) Identifies an Ocellatin with Broad Spectrum Antimicrobial Activity


 , , ,
, , ,  ,
,  and
and 
Abstract
1. Introduction
2. Results
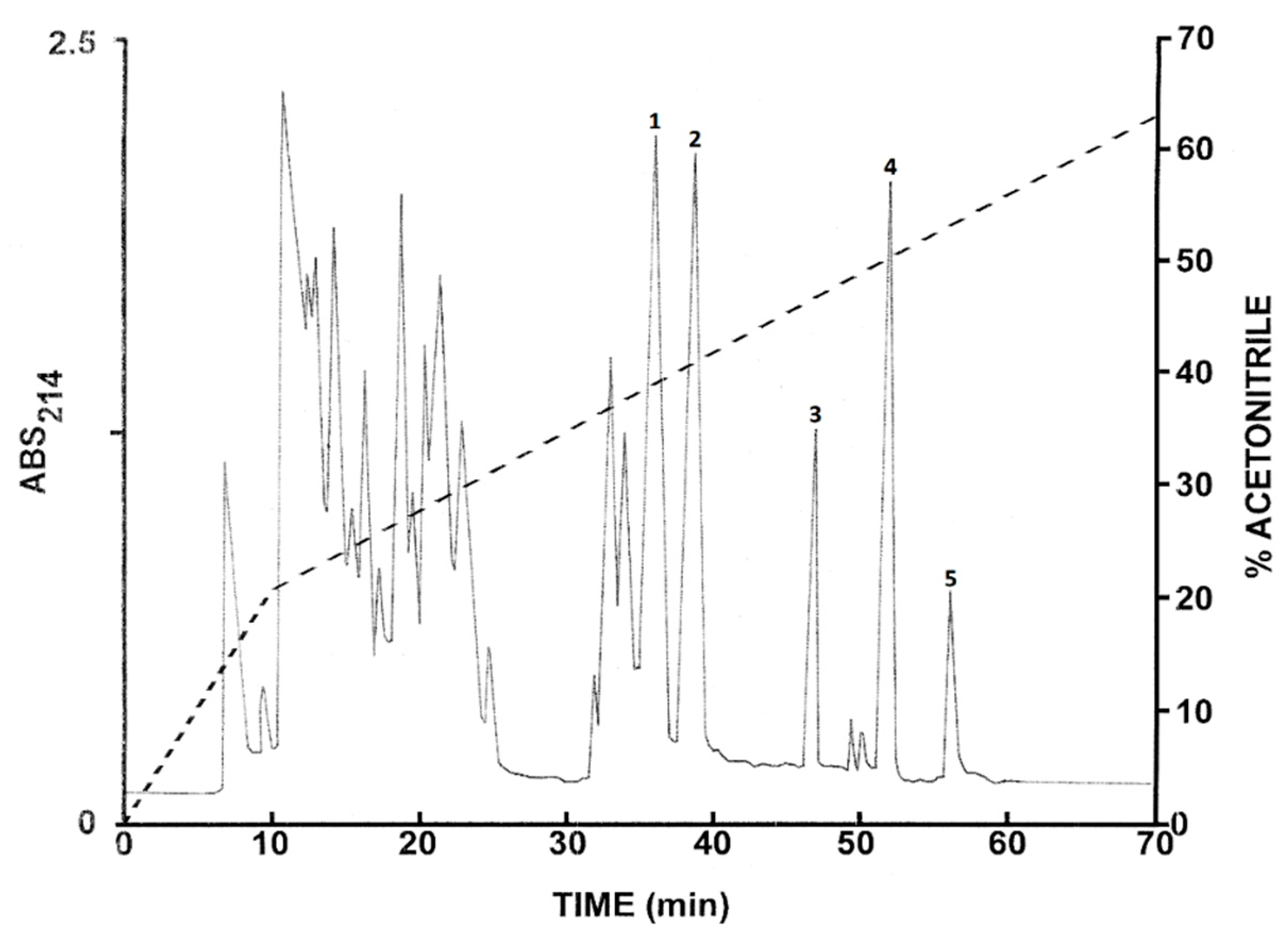
2.1. Purification of the Peptides from L. insularum
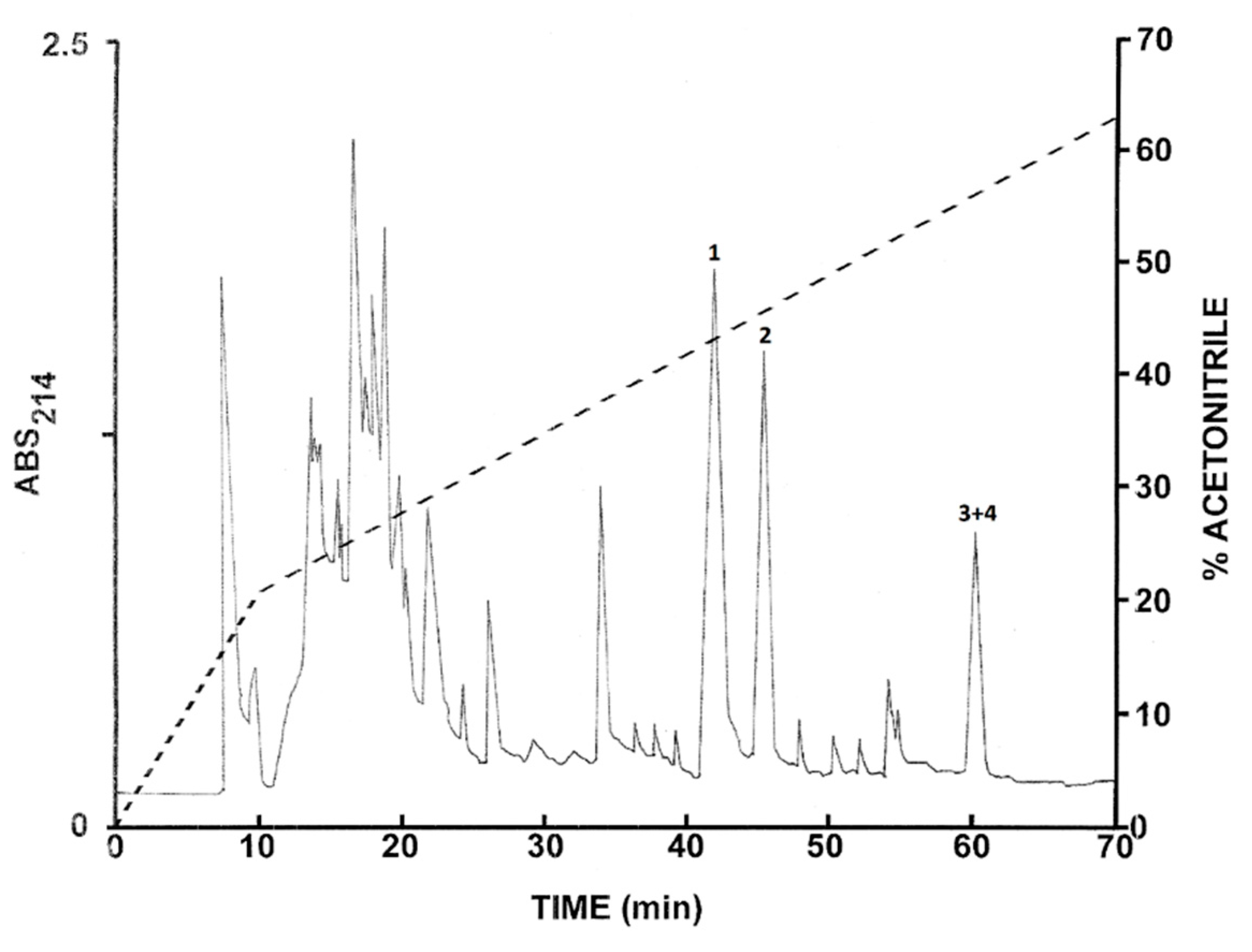
2.2. Purification of the Peptides from L. nesiotus
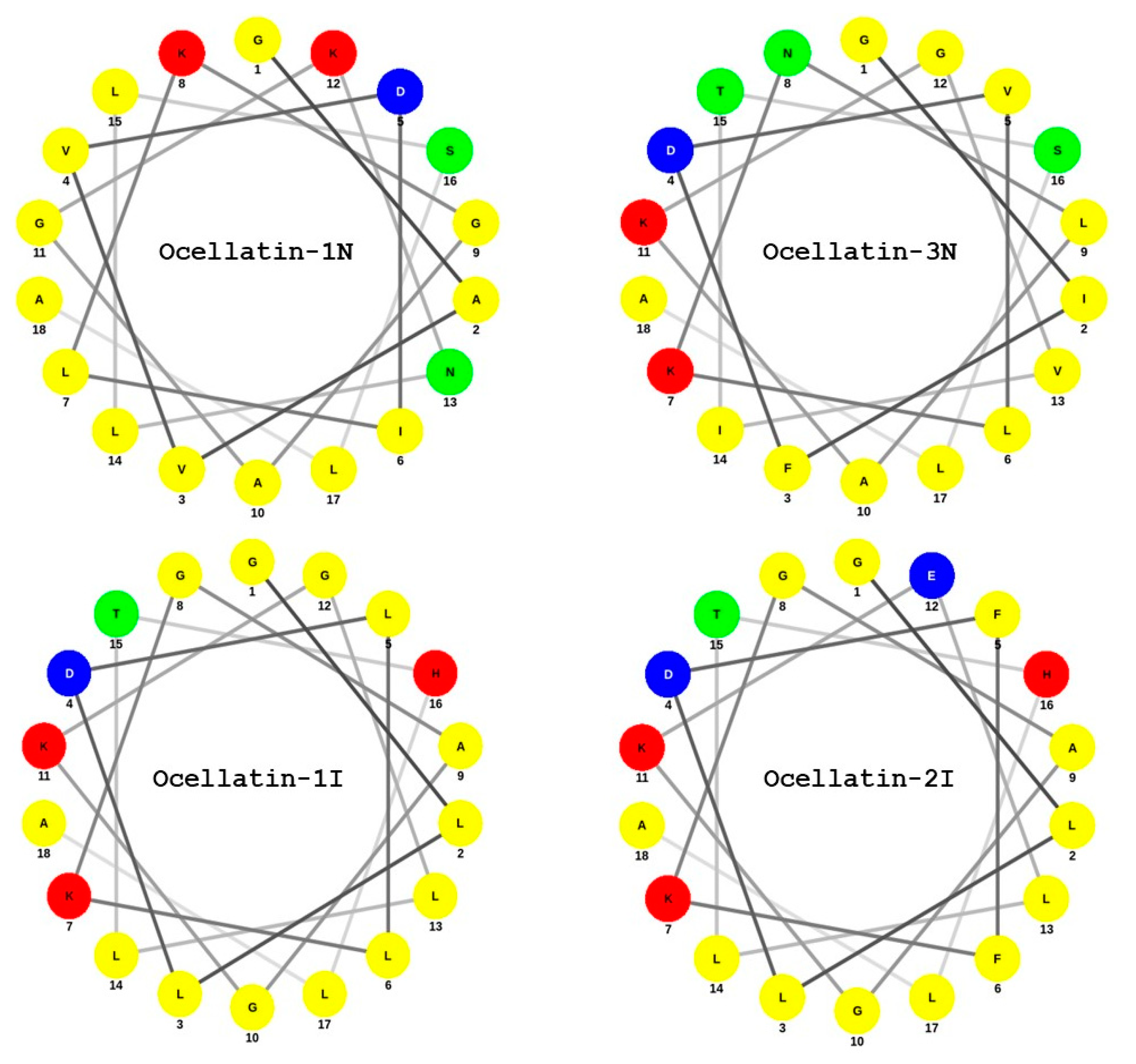
2.3. Structural Characterization
2.4. Antimicrobial and Hemolytic Activities
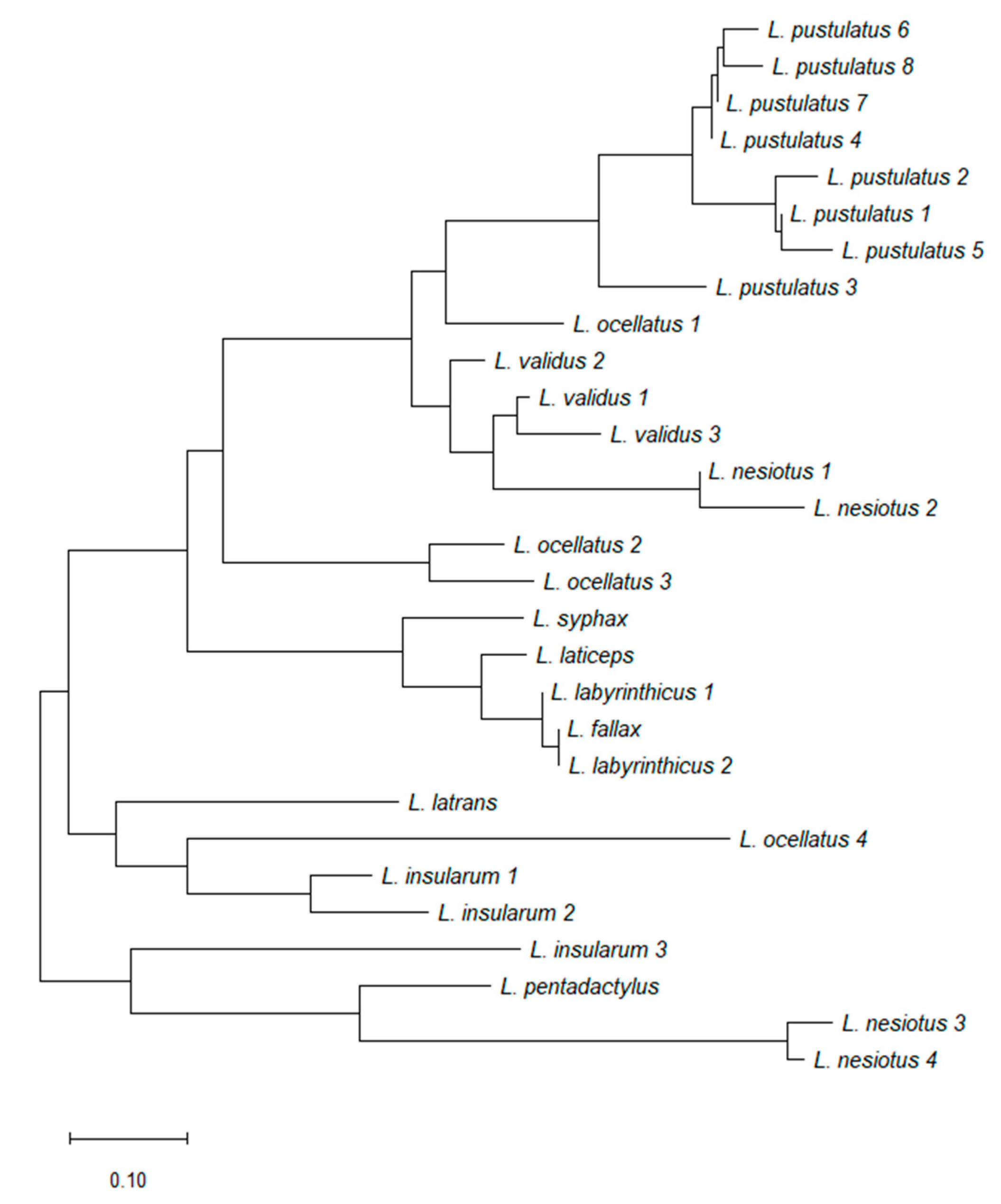
2.5. Cladistic Analysis
3. Discussion
4. Materials and Methods
4.1. Collection of Skin Secretions
4.2. Purification of the Peptides
4.3. Structural Characterization
4.4. Synthetic Peptides
4.5. Antimicrobial Assays
4.6. Hemolysis Assay
4.7. Cladistic Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Media Centre. News Release. WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. 2017. Available online: http://www.who.int/mediacentre/news/releases/2017/bacteria-antibiotics-needed/en/ (accessed on 19 August 2020).
- Bassetti, M.; Peghin, M.; Vena, A.; Giacobbe, D.R. Treatment of infections due to MDR Gram-negative bacteria. Front. Med. 2019, 6, 74. [Google Scholar] [CrossRef] [PubMed]
- WHO. News Room. Addressing the Crisis in Antibiotic Development. 2020. Available online: https://www.who.int/news/item/09-07-2020-addressing-the-crisis-in-antibiotic-development (accessed on 19 August 2020).
- WHO. WHO Director-General. Speeches. WHO Director-General Opening Remarks at the AMR Action Fund Launch. 2020. Available online: https://www.who.int/dg/speeches/detail/who-director-general-opening-remarks-at-the-amr-action-fund-launch (accessed on 19 August 2020).
- Hollmann, A.; Martinez, M.; Maturana, P.; Semorile, L.C.; Maffia, P.C. Antimicrobial Peptides: Interaction with model and biological membranes and synergism with chemical antibiotics. Front. Chem. 2018, 6, 204. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M. Structural diversity and species distribution of host-defense peptides in frog skin secretions. Cell. Mol. Life Sci. 2011, 68, 2303–2315. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Lai, R. The chemistry and biological activities of peptides from amphibian skin secretions. Chem. Rev. 2015, 115, 1760–1846. [Google Scholar] [CrossRef]
- Ladram, A.; Nicolas, P. Antimicrobial peptides from frog skin: Biodiversity and therapeutic promises. Front. Biosci. 2016, 21, 1341–1371. [Google Scholar] [CrossRef]
- Conlon, J.M.; Mechkarska, M.; Leprince, J. Peptidomic analysis in the discovery of therapeutically valuable peptides in amphibian skin secretions. Expert Rev. Proteom. 2019, 16, 897–908. [Google Scholar] [CrossRef]
- Patocka, J.; Nepovimova, E.; Klimova, B.; Wu, Q.; Kuca, K. Antimicrobial peptides: Amphibian host defense peptides. Curr. Med. Chem. 2019, 26, 5924–5946. [Google Scholar] [CrossRef]
- Wang, G. Bioinformatic analysis of 1000 amphibian antimicrobial peptides uncovers multiple length-dependent correlations for peptide design and prediction. Antibiotics 2020, 9, 491. [Google Scholar] [CrossRef]
- Tennessen, J.A.; Blouin, M.S. Balancing selection at a frog antimicrobial peptide locus: Fluctuating immune effector alleles? Mol. Biol. Evol. 2008, 12, 2669–2680. [Google Scholar] [CrossRef]
- Conlon, J.M. The contribution of skin antimicrobial peptides to the system of innate immunity in anurans. Cell Tissue Res. 2011, 343, 201–212. [Google Scholar] [CrossRef]
- Conlon, J.M.; Mechkarska, M.; Lukic, M.L.; Flatt, P.R. Potential therapeutic applications of multifunctional host-defense peptides from frog skin as anti-cancer, anti-viral, immunomodulatory, and anti-diabetic agents. Peptides 2014, 57, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Mechkarska, M.; Kolodziejek, J.; Musale, V.; Coquet, L.; Leprince, J.; Jouenne, T.; Nowotny, N.; Conlon, J.M. Peptidomic analysis of the host-defense peptides in skin secretions of Rana graeca provides insight into phylogenetic relationships among Eurasian Rana species. Comp. BioChem. Physiol. Part D Genom. Proteom. 2019, 29, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Mechkarska, M.; Coquet, L.; Leprince, J.; Auguste, R.J.; Jouenne, T.; Conlon, J.M. Peptidomic analysis of the host-defense peptides in skin secretions of the Trinidadian leaf frog Phyllomedusa trinitatis (Phyllomedusidae). Comp. BioChem. Physiol. Part D Genom. Proteom. 2018, 28, 72–79. [Google Scholar] [CrossRef]
- Conlon, J.M.; Mechkarska, M.; Coquet, L.; Leprince, J.; Jouenne, T.; Vaudry, H.; Measey, G.J. Evidence from peptidomic analysis of skin secretions that allopatric populations of Xenopus gilli (Anura:Pipidae) constitute distinct lineages. Peptides 2014, 63C, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Frost, D.R. Amphibian Species of the World; An Online Reference. Version 6.1; American Museum of Natural History: New York, NY, USA, 2020; Available online: https://amphibiansoftheworld.amnh.org/index.php (accessed on 19 August 2020). [CrossRef]
- Nascimento, A.C.C.; Zanotta, L.C.; Kyaw, C.M.; Schwartz, E.N.F.; Schwartz, C.A.; Sebben, A.; Sousa, M.V.; Fontes, W.; Castro, M.S. Ocellatins: New antimicrobial peptides from the skin secretion of the South American Frog Leptodactylus ocellatus (Anura: Leptodactylidae). Protein J. 2004, 23, 501–508. [Google Scholar] [CrossRef]
- Nascimento, A.; Chapeaurouge, A.; Perales, J.; Sebben, A.; Sousa, M.V.; Fontes, W.; Castro, M.S. Purification, characterization and homology analysis of ocellatin 4, a cytolytic peptide from the skin secretion of the frog Leptodactylus ocellatus. Toxicon 2007, 8, 1095–1104. [Google Scholar] [CrossRef]
- Leite, J.M.A., Jr.; Silva, L.P.; Silva-Leite, R.R.; Ferrari, A.S.; Noronha, S.E.; Silva, H.R.; Bloch, C., Jr.; Leite, J.R.S.A. Leptodactylus ocellatus (Amphibia): Mechanism of defense in the skin and molecular phylogenetic relationships. J. Exp. Zool. 2010, 313A, 1–8. [Google Scholar] [CrossRef]
- Rollins-Smith, L.A.; King, J.D.; Nielsen, P.F.; Sonnevend, A.; Conlon, J.M. An antimicrobial peptide from the skin secretions of the mountain chicken frog Leptodactylus fallax (Anura: Leptodactylidae). Regul. Pept. 2005, 124, 173–178. [Google Scholar] [CrossRef]
- King, J.D.; Al-Ghaferi, N.; Abraham, B.; Sonnevend, A.; Leprince, J.; Nielsen, P.F.; Conlon, J.M. Pentadactylin: An antimicrobial peptide from the skin secretions of the South American bullfrog Leptodactylus pentadactylus. Comp. Biochem. Physiol. Part C 2005, 141, 393–397. [Google Scholar] [CrossRef]
- Sousa, J.C.; Berto, R.F.; Gois, E.A.; Fontenele-Cardi, N.C.; Honório-Júnior, J.E.R.; Konno, K.; Richardson, M.; Rocha, M.F.G.; Camargo, A.A.C.M.; Pimenta, D.C.; et al. Leptoglycin: A new glycine/leucine-rich antimicrobial peptide isolated from the skin secretion of the South American frog Leptodactylus pentadactylus (Leptodactylidae). Toxicon 2009, 54, 23–32. [Google Scholar] [CrossRef]
- Conlon, J.M.; Al-Ghaferi, N.; Abraham, B.; Sonnevend, A.; King, J.D.; Nielsen, P.F. Purification and properties of laticeptin, an antimicrobial peptide from skin secretions of the South American frog Leptodactylus laticeps. Protein Pept. Lett. 2006, 13, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Abdel-Wahab, Y.H.A.; Flatt, P.R.; Leprince, J.; Vaudry, H.; Jouenne, T.; Condamine, E. A glycine-leucine-rich peptide structurally related to the plasticins from skin secretions of the frog Leptodactylus laticeps (Leptodactylidae). Peptides 2009, 5, 888–892. [Google Scholar] [CrossRef]
- Dourado, F.S.; Leite, J.R.S.A.; Silva, L.P.; Melo, J.A.T.; Bloch, C., Jr.; Schwartz, E.F. Antimicrobial peptide from the skin secretion of the frog Leptodactylus syphax. Toxicon 2007, 50, 572–580. [Google Scholar] [CrossRef]
- King, J.D.; Leprince, J.; Vaudry, H.; Coquet, L.; Jouenne, T.; Conlon, J.M. Purification and characterization of antimicrobial peptides from the Caribbean frog, Leptodactylus validus (Anura: Leptodactylidae). Peptides 2008, 29, 1287–1292. [Google Scholar] [CrossRef] [PubMed]
- Liberio, M.S.; Joanitti, G.A.; Azevedo, R.B.; Cilli, E.M.; Zanotta, L.C.; Nascimento, A.C.C.; Sousa, M.V.; Pires, O.R., Jr.; Fontes, W.; Castro, M.S. Anti-proliferative and cytotoxic activity of pentadactylin isolated from Leptodactylus labyrinthicus on melanoma cells. Amino Acids 2011, 40, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Gusmão, K.A.G.; dos Santos, D.M.; Santos, V.M.; Cortes, M.E.; Reis, P.V.M.; Santos, V.L.; Pilo-Veloso, D.; Verly, R.M.; de Lima, M.E.; Resende, J.M. Ocellatin peptides from the skin secretion of the South American frog Leptodactylus labyrinthicus (Leptodactylidae): Characterization, antimicrobial activities and membrane interactions. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 4. [Google Scholar] [CrossRef] [PubMed]
- Marani, M.M.; Dourado, F.S.; Quelemes, P.V.; de Araujo, A.R.; Perfeito, M.L.G.; Barbosa, E.A.; Veras, M.L.C.; Coelho, A.L.R.; Andrade, E.B.; Eaton, P.; et al. Characterization and biological activities of ocellatin peptides from the skin secretion of the frog Leptodactylus pustulatus. J. Nat. Prod. 2015, 78, 1495–1504. [Google Scholar] [CrossRef]
- Siano, A.; Humpola, M.V.; de Oliveira, E.; Albericio, F.; Simonetta, A.C.; Lajmanovich, R.; Tonarelli, G.G. Leptodactylus latrans amphibian skin secretions as a novel source for the isolation of antibacterial peptides. Molecules 2018, 23, 2943. [Google Scholar] [CrossRef]
- Sousa, N.A.; Oliveira, G.A.; de Oliveira, A.P.; Lopes, A.L.F.; Iles, B.; Nogueira, K.M.; Araújo, T.S.; Souza, L.K.; Araújo, A.R.; Ramos-Jesus, J.; et al. Novel ocellatin peptides mitigate LPS-induced ROS formation and NF-kB activation in microglia and hippocampal neurons. Sci. Rep. 2020, 10, 2696. [Google Scholar] [CrossRef]
- Conlon, J.M. A proposed nomenclature for antimicrobial peptides from frogs of the genus Leptodactylus. Peptides 2008, 29, 1631–1632. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.C.; Downie, J.R.; Smith, J.M.; Livingstone, S.R.; Mohammed, R.S.; Lehtinen, R.M.; Eyre, M.; Sewlal, J.; Noriega, N.; Caspar, G.S.; et al. A Field Guide to the Amphibians and Reptiles of Trinidad and Tobago; Trinidad and Tobago Field Naturalists’ Club: Port of Spain, Trinidad and Tobago, 2018; p. 336. ISBN -978-976-8255-47-1. [Google Scholar]
- Jairam, R.; Fouquet, A. First records of Leptodactylus nesiotus (Anura:Leptodactylidae) for Guyana, Suriname and French Guiana. Herpetol. Notes 2018, 11, 997–999. [Google Scholar]
- Kyte, J.; Doolittle, D.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef]
- Muñoz, V.; Serrano, L. Elucidating the folding problem of helical peptides using empirical parameters. Nat. Struct. Biol. 1994, 1, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Giangaspero, A.; Sandri, L.; Tossi, A. Amphipathic α-helical peptides. A systematic study of the effects of structural and physical properties on biological activity. Eur. J. Biochem. 2001, 268, 5589–5600. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef]
- Conlon, J.M.; Al-Ghaferi, N.; Abraham, B.; Leprince, J. Strategies for development of naturally occurring antimicrobial peptides into therapeutically valuable anti-infective agents. Methods 2007, 42, 349–357. [Google Scholar] [CrossRef]
- Haney, E.F.; Mansour, S.C.; Hancock, R.E.W. Antimicrobial Peptides: An Introduction. In Antimicrobial Peptides. Methods in Molecular Biology; Hansen, P., Ed.; Humana Press: New York, NY, USA, 2017; Volume 1548. [Google Scholar] [CrossRef]
- Dathe, M.; Wieprecht, T.; Nikolenko, H.; Handel, L.; Maloy, W.L.; MacDonald, D.L.; Beyermann, M.; Bienert, M. Hydrophobicity, hydrophobic moment and angle subtended by charged residues modulate antibacterial and haemolytic activity of amphipathic helical peptides. FEBS Lett. 1997, 403, 208–212. [Google Scholar] [CrossRef]
- Schiffer, M.; Edmundson, A.B. Use of helical wheels to represent the structures of proteins and to identify segments with helical potential. Biophys. J. 1967, 7, 121–135. [Google Scholar] [CrossRef]
- Bessa, L.J.; Eaton, P.; Dematei, A.; Plácido, A.; Vale, N.; Gomes, P.; Delerue-Matos, C.; Leite, J.R.S.A.; Gameiro, P. Synergistic and antibiofilm properties of ocellatin peptides against multidrug-resistant Pseudomonas aeruginosa. Future Microbiol. 2018, 13, 151–163. [Google Scholar] [CrossRef]
- Gharibkandi, N.A.; Conlon, J.M.; Hosseinimehr, S.J. Strategies for improving stability and pharmacokinetic characteristics of radiolabeled peptides for imaging and therapy. Peptides 2020, 170385. [Google Scholar] [CrossRef]
- Patrulea, V.; Borchard, G.; Jordan, O. An update on antimicrobial peptides (AMPs) and their delivery strategies for wound infections. Pharmaceutics 2020, 12, 840. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, M.C.; Carbone, C.; Sousa, M.C.; Espina, M.; Garcia, M.L.; Sanchez-Lopez, E.; Souto, E.B. Nanomedicines for the delivery of antimicrobial peptides (AMPs). Nanomaterials 2020, 10, 560. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Sonnevend, A. Antimicrobial peptides in frog skin secretions. Methods Mol. Biol. 2010, 618, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Leprince, J. Identification and analysis of bioactive peptides in amphibian skin secretions. Methods Mol. Biol. 2010, 615, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Moffett, R.C.; Leprince, J.; Flatt, P.R. Identification of components in frog skin secretions with therapeutic potential as antidiabetic agents. Methods Mol. Biol. 2018, 1719, 319–333. [Google Scholar] [CrossRef]
- Clinical Laboratory and Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; Approved Standard M07; CLSI: Wayne, PA, USA, 2019. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Zuckerkandl, E.; Pauling, L. Evolutionary Divergence and Convergence in Proteins. In Evolving Genes and Proteins; Bryson, V., Vogel, H.J., Eds.; Academic Press: New York, NY, USA, 1965; pp. 97–166. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | No. of Amino Acids | Charge at pH 7 | pI | GRAVY | Alpha-Helical Domain * |
|---|---|---|---|---|---|
| Ocellatin-1I (1–16) | 16 | +1 | 9.9 | 0.488 | 2–9 |
| Ocellatin-2I (1–16) | 16 | 0 | 7.8 | 0.169 | none |
| Ocellatin-1I | 21 | +2 | 10.6 | 0.648 | 2–9, 13–19 |
| Ocellatin-2I | 21 | +1 | 9.9 | 0.405 | 11–19 |
| Ocellatin-3I | 26 | +1 | 9.8 | 0.205 | 2–10, 13–21 |
| Ocellatin-1N | 26 | +3 | 10.6 | 0.496 | 3–10, 12–24 |
| Ocellatin-2N | 26 | +2 | 10.2 | 0.281 | 12–24 |
| Ocellatin-3N | 19 | +2 | 10.2 | 0.911 | 4–11, 13–18 |
| Ocellatin-4N | 19 | +2 | 10.2 | 0.874 | 2–11, 13–18 |
| Bacteria | Ocellatin-1I | Ocellatin-2I | Ocellatin-1N | Ocellatin-3N |
|---|---|---|---|---|
| Gram-positive | ||||
| S. aureus (ATCC 12600) | 250 (575) | >250 (>620) | 250 (725) | 31.25 (67) |
| S. aureus (ATCC BAA-2312) | 250 (575) | >250 (>620) | 250 (725) | 31.25 (67) |
| E. faecium (ATCC 19434) | n.d. | 250 (620) | 250 (725) | 62.5 (134) |
| E. faecalis (ATCC 51299) | >250 (>575) | >250 (>620) | >250 (>725) | 250 (535) |
| Gram-negative | ||||
| E. coli (ATCC 35218) | 62.5 (144) | 62.5 (155) | 62.5 (181) | 31.25 (67) |
| K. pneumoniae (ATCC 49472) | 125 (288) | 125 (310) | 125 (362) | 62.5 (134) |
| K. pneumoniae (ATCC BAA-2814) * | >125 (>288) | 125 (310) | 125 (362) | 62.5 (134) |
| P. aeruginosa (ATCC 27853) | n.d. | >125 (>310) | >125 (>362) | 62.5 (134) |
| S. typhimurium (ATCC 14028) | 250 (575) | 125 (310) | 250 (725) | 62.5 (134) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barran, G.; Kolodziejek, J.; Coquet, L.; Leprince, J.; Jouenne, T.; Nowotny, N.; Conlon, J.M.; Mechkarska, M. Peptidomic Analysis of Skin Secretions of the Caribbean Frogs Leptodactylus insularum and Leptodactylus nesiotus (Leptodactylidae) Identifies an Ocellatin with Broad Spectrum Antimicrobial Activity. Antibiotics 2020, 9, 718. https://doi.org/10.3390/antibiotics9100718
Barran G, Kolodziejek J, Coquet L, Leprince J, Jouenne T, Nowotny N, Conlon JM, Mechkarska M. Peptidomic Analysis of Skin Secretions of the Caribbean Frogs Leptodactylus insularum and Leptodactylus nesiotus (Leptodactylidae) Identifies an Ocellatin with Broad Spectrum Antimicrobial Activity. Antibiotics. 2020; 9(10):718. https://doi.org/10.3390/antibiotics9100718
Chicago/Turabian StyleBarran, Gervonne, Jolanta Kolodziejek, Laurent Coquet, Jérôme Leprince, Thierry Jouenne, Norbert Nowotny, J. Michael Conlon, and Milena Mechkarska. 2020. "Peptidomic Analysis of Skin Secretions of the Caribbean Frogs Leptodactylus insularum and Leptodactylus nesiotus (Leptodactylidae) Identifies an Ocellatin with Broad Spectrum Antimicrobial Activity" Antibiotics 9, no. 10: 718. https://doi.org/10.3390/antibiotics9100718
APA StyleBarran, G., Kolodziejek, J., Coquet, L., Leprince, J., Jouenne, T., Nowotny, N., Conlon, J. M., & Mechkarska, M. (2020). Peptidomic Analysis of Skin Secretions of the Caribbean Frogs Leptodactylus insularum and Leptodactylus nesiotus (Leptodactylidae) Identifies an Ocellatin with Broad Spectrum Antimicrobial Activity. Antibiotics, 9(10), 718. https://doi.org/10.3390/antibiotics9100718