Abstract
The emergence of blaKPC-2 and blaNDM-1 producing Klebsiella pneumoniae represents a great problem in many Egyptian hospitals. One hundred and twenty-six K. pneumoniae isolates from patients admitted to Assiut University Hospital were identified by an API20E kit. Carbapenemase-producing K. pneumoniae (CPKP) was detected by the modified carbapenem inactivation method (mCIM), the EDTA-modified carbapenem inactivation method (eCIM), and an E-test. Based on the polymerase chain reaction, all isolates were negative for bla-VIM-1 and bla-IMP-1, fifteen of these isolates were positive for both blaKPC-2 and blaNDM-1, two isolates were positive for blaKPC-2 only, and twenty-eight isolates were positive for bla-NDM-1 only. Although one isolate was positive for the string test, all CPKP isolates were negative for capsular genes. Only 71.1% of CPKP transferred their plasmids to their corresponding transconjugants (E. coli J53). The resistance patterns of the clinical isolates and their transconjugates were similar, except for 12 isolates, which showed differences with their transconjugates in the resistance profile of four antibiotics. Molecular typing of the plasmids based on replicon typing showed that Inc FIIK and FII plasmids predominated in isolates and their transconjugants carrying blaKPC-2 and/or blaNDM-1. Conjugative Inc FII plasmids play an important role in the spread of CPKP, and their recognition is essential to limit their spread.
1. Introduction
K. pneumonia is a common bacterial pathogen that can cause several life-threatening infections, such as blood stream infections, pneumonia, as well as urinary tract, post-surgical, and intensive care-related infections [1,2]. Such infections result in significant morbidity and mortality. K. pneumonia also significantly increases medical hospital costs [3]. It can spread well and survive in the hospital environment and frequently causes nosocomial infections and outbreaks [4]. K. pneumonia’s major route of infection is aspiration of the oropharyngeal secretions of the patients colonized by it. The gastrointestinal tract of patients may also play a secondary role. Further, contaminated medical equipment and the hands of health care workers may contribute to sustaining colonization and creating an increased risk of infection [5]. Additionally, antibiotic therapy can suppress normal bacterial flora and lead to an over-growth of multidrug resistant K. pneumoniae [6].
The increasing incidence of carbapenemase-producing K. pneumoniae (CPKP) has become a great challenge for infection control in human health and has resulted in treatment failure [7]. K. pneumoniae was found to harbor plasmid-mediated carbapenemase of different classes, such as Class A (e.g., KPC), Class B as Verona integron-encoded MBL (VIM), imipenemase (IMP) and New Delhi-metallo beta-lactamase (NDM) or Class D (e.g., oxacillinases (OXA) enzymes [8,9]. Carbapenem resistant K. pneumoniae was reported and isolated from many Egyptian hospitalized patients [10,11,12]. Furthermore, the co-existence of carbapenems and other drug-resistance determinants, such as aminoglycoside and quinolone resistance genes in K. pneumoniae, was found to be a great challenge in treating infections caused by this pathogen [8].
Large conjugative plasmid-carrying genes that encode resistance were found to have a great role in the spread and acquisition of resistance among K. pneumoniae strains [13,14]. The molecular typing of resistance plasmids based on replicon typing is currently applied to study the acquisition and spread of antimicrobial resistance in K. pneumoniae [15]. Carbapenemase-producing K. pneumoniae (KPC) and New Delhi-metallo beta-lactamase (NDM) are the most noteworthy carbapenemase-encoding genes that are found to enable the spread of these resistance genes in transferable plasmids. Thus, tracing the source of endemic plasmids that could be responsible for the spread of a resistance trait among strains is the backbone for the development of efficient treatment policies, as well as applicable infection control measures [16,17,18,19]. Treating infections with antibiotics that are inactive against the causative agent leads to increasing the selective pressure for nosocomial organisms to express antibiotic resistance and dissemination and the persistence of an endemic plasmid among strains [20]. This means that the withdrawal of ineffective drug, earlier correct drug selection, or using effective combinations with reliable spectra of activity and effective treatments against these pathogens may stop selection pressure and interfere with the transmission of the plasmid [21,22]. Although many studies have recently reported the emergence of K. pneumoniae harboring NDM or KPC in Egypt, limited studies are available on plasmids responsible for transferring resistance determinants in multidrug resistant K. pneumoniae in this country. As a result, our aim was to study and identify the plasmid type responsible for the transmission of blaNDM-1 and blaKPC-2 in K. pneumoniae in Egypt in order to trace the transfer of this plasmid among K. pneumoniae strains [11].
2. Results
2.1. Cabapenemase Production by K. pneumoniae
One hundred and twenty-six K. pneumoniae strains isolated from different departments at Assiut University Hospital (April 2018 to May 2019) were tested for their production of carbapenemase enzymes using mCIM. Carbapenemase activity was found in 77/126 (61.1%) of the K. pneumoniae isolates. K. pneumoniae isolates positive for mCIM were tested for Metallo-β-lactamase production using the eCIM method. Out of 77 carbapenemase-producing K. pneumoniae, 51 isolates (66.2%) were positive for eCIM. In addition, it was observed that 61/77 of carbapenemase producing K. pneumoniae were imipenem resistant, with MIC of ≥4 μg/mL.
2.2. Hypermucoviscocity Phenotyping
Only one isolate (Kp27) showed the hypermucoviscocity phenotype (Positive string test).
2.3. Antimicrobial Resistance Pattern of Carbapenemase Producing K. pneumoniae (CPKP)
The antibacterial susceptibility profile was tested for all 77 CPKP isolates. It was found that Carbapenemase producing K. pneumoniae were completely resistant to Aztreonam, Amoxicillin, Amox-Clavulanic acid, Piperacillin, Ceftriaxone, Ceftazidime, Cefoperazone, Cefpodoxime, and Meropenem but showed high resistance to the other tested antibiotics (Figure 1).
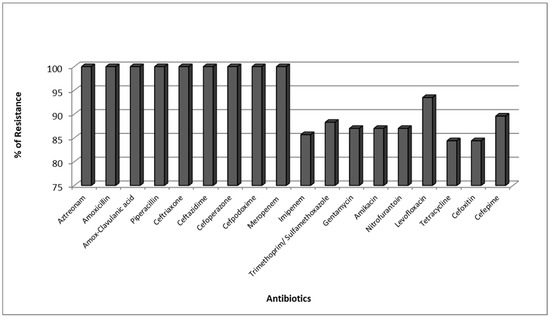
Figure 1.
Resistance Pattern of Carbapenemase producing K. pneumoniae (CPKP).
2.4. PCR for Carbapenemase Genes and KI and K2 Capsular Genes
All K. pneumoniae isolates were tested for carbapenemase genes and K1 and K2 capsular genes. It was found that all non CPKP were negative for these genes. On the other hand, Carbapenemase-encoding genes were founded in 45/77 (58.44%) of carbapenemase-producing K. pneumoniae isolates. BlaNDM-1 was present in 43/77 (55.84%) isolates and blaKPC-2 in 17/77 (22.07%) isolates.
All CPKP isolates were negative for blaVIM-1, blaIMP-1, wzy_K1, and wzy_K2 capsular genes (Figure 2). Among the isolates tested positive for carbapenemase genes, fifteen (15/45, 33.3%) were positive for both blaNDM-1 and blaKPC-2. Two isolates (2/45, 4.4%) were positive for blaKPC2 only and twenty-eight isolates (28/45, 62.2%) for blaNDM-1 only.
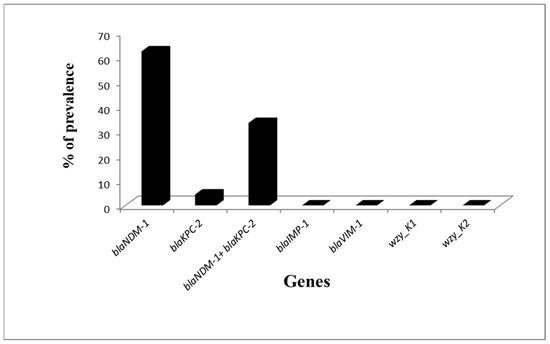
Figure 2.
Prevalence of Carbapenemase genes and capsular genes among Carbapenemase producing K. pneumoniae (CPKP).
2.5. Comparison of Phenotypic and Genotypic Methods for Detection of CPKP
Isolates were divided into four groups according to their positivity for the tested genes (Table 1). The isolates negative for all genes (Group 1) were negative for mCIM. Out of the 28 isolates (Group 2) that tested positive for blaNDM-1 and mCIM, nine isolates were metallo-β-lactamases (MBL) producers (positive for eCIM), while the two isolates (KP8, KP17) positive for blaKPC-2 (Group 3) were positive for the mCIM and eCIM test (Table 1). In addition, the five isolates of Group 4, which were positive for blaNDM1 and blaKPC2 and mCIM, were positive for eCIM and were considered to be MBL producers.

Table 1.
Phenotypic and genotypic methods for the detection of CPKP.
2.6. Plasmid Transfers
Conjugation experiments were carried out for all blaKPC-2 and/or blaNDM-1 carrying K. pneumoniae isolates (n = 45). Only 32 of these were found to have the ability to transfer their plasmids by conjugation to their corresponding transconjugants (E. coli J53). These transconjugants were tested for blaKPC-2 and/or blaNDM-1 genes by the PCR method. All thirty-two transconjugants were found to harbor blaKPC-2 and/or blaNDM-1 genes as their parent cells.
All transconjugants were observed to exhibit Multi-drug resistance (MDR) phenotypes like those of the donor K. pneumoniae isolates, except 12 of the transconjugants were found to have different antibiotic resistance profiles. The difference in the resistance profiles among these isolates was observed with the following antibiotics: Tetracycline, Gentamicin, Sulfamethoxazole/trimethoprim, and Levofloxacin. The resistance profiles of the 12 tested isolates and transconjugates against these four antibiotics are listed in Table 2.
Table 2.
Resistance patterns of the 12 isolates and transconjugates showing different resistance profiles.
2.7. PCR-Based Replicon Typing of blaKPC2 and/or blaNDM1 Encoding K. pneumoniae
PCR-based replicon typing (PBRT) showed typing for only 29/32 of the self-transmissible blaKPC-2 and/or blaNDM-1 encoding K. pneumoniae isolates. The replicon types identified were FIIK, FII, FIB, FIC, L, and Inc M. It was found that FIIK and FII were highly distributed among blaKPC-2 and/or blaNDM-1 encoding K. pneumoniae isolates, followed by the FIB, FIC, Inc L, and Inc M groups. Furthermore, PBRT failed to detect the replicon types of the plasmids of the remaining three isolates (Kp5, Kp6, Kp22) (Table 3).
Table 3.
Relation between the plasmid replicon type and the transfer of resistant genes.
2.8. Transfer of Plasmids with a Specific Replicon Type to the Transconjugants
PCR-based replicon typing of the transconjugants (E. coli J53) carrying blaKPC-2 and/or blaNDM-1 encoding genes (n = 32) revealed that nine (28.1%) of the strains were positive for Inc FIIK alone, while three (9.3%) of the transconjugants were positive for all FII, FIIK, and FIB incompatibility types. Two (6.25%) of the strains carried plasmids belonging to the Inc FII group only, whereas Inc FIB was found in one (3.1%) of them. On the other hand, PBRT confirmed the transfer of Inc FII and FIIK plasmids to two transconjugants and the transfer of Inc FIIK and FIB groups to other two transconjugants. The remaining strains (n = 13) could not be typed by the PBRT method (Table 3). Also, it was observed that Inc L, M, and FIC plasmids did not show the ability to transfer to the recipient cells.
3. Discussion
Carbapenemase-producing Klebsiella pneumoniae (CPKP) is one of the most widespread pathogens among hospital-acquired multi-drug resistant pathogen associated infections [7]. Although many reports exhibited the emergence of carbapenem resistant K. pneumoniae in Egypt [12,23] in the last decades, the data on plasmid epidemiology and which plasmid types are mainly associated with the spread of blaKPC2 and/or blaNDM1 plasmids in Egypt are rare. Thus, in this study, we attempted to determine the plasmids responsible for transferring the determinants of carbapenem-resistance among K. pneumoniae isolates in this country.
In the present study, carbapenemase phenotypic activity was detected in 77/126 (61.1%) of the K. pneumoniae isolates by an mCIM test. Although the rate we observed was lower than that in a study in the USA [24], it was higher than the previous reports in Egypt. The prevalence of CPKP was found to be 33.3% among K. pneumoniae isolates in a study performed by Moemen and Masallat [23]; 44.3% at the Suez Canal University Hospitals in a study performed by El-Sweify et al. [10]; and 13.9% in the Egyptian National Cancer Institute in a study performed by Ashour and El-Sharif [25]. Increasing rates of CPKP isolates may be due to the misuse of carbapenems, as carbapenems are considered the best choice for the treatment of serious infections caused by multi-drug resistant pathogens in our hospitals, where there is presently no implementation of an antimicrobial stewardship program [26].
Our results show that the mCIM negative isolates were also negative for both blaKPC2 and/or blaNDM1, and there was compatibility between the results of the mCIM and PCR results among carbapenemase producing isolates. The sensitivity of this test was 100% compared to PCR, the gold standard test, suggesting that this test can be used as a primary test for the detection of CPKP. On the contrary, the eCIM test did not identify isolates that were positive for both blaKPC2 and blaNDM1 in accordance with the CLSI [27] report.
The capsular serotypes K1 and K2 of K. pneumoniae are the predominant virulent strains that contribute to the high mortality rate associated with K. pneumoniae infections [28]. Although there was only one isolate (Kp27) positive for the string test, all CPKP isolates were negative for the K1 and K2 capsular gene. This may be due to the fact that antibiotic resistance is not dependent from the hypervirulence of KP [29], and isolates were recovered from hospital-acquired infections, not from community-acquired infections [30,31]. In addition, the incidence of blaNDM1 producers among the CPKP in our hospital (55.84%) was much higher than that previously reported by Yan et al. [8]. BlaNDM1 was first detected in Egypt from one K. pneumoniae isolate in 2013 by Abdelaziz et al. [32] and then from two Pseudomonas aeruginosa isolates [33], as well as in Acinetobacter baumannii isolates [34], which indicates that this gene may spread among Enterobacteriaceae by transferable plasmids [35,36].
In Egypt, the plasmid types that are mainly associated with the spread of blaKPC2 and/or blaNDM1 have not been identified, except for the eight carbapenem-resistant NDM1-producing K. pneumoniae isolates carried by non-transferrable plasmids (either IncR or untypeable), which were reported by Gamal et al. [11].
Our study showed that most blaKPC2 and/or blaNDM1 plasmids were successfully transferred by conjugation and that blaKPC2 and/or blaNDM1 were commonly carried on the Inc FII plasmid. Similar results were reported by [37], who reported that blaKPC-2 was located on Inc FII plasmids, but in another study performed by Jin et al. [38], blaKPC2 was found to be located on non-self-transmissible plasmids or on the chromosome.
Our results showed the predominance of both Inc FIIK and FII plasmids among the isolates and their transconjugants carrying blaKPC2 and/or blaNDM1, suggesting that these types of plasmids mediate horizontal transmission and contribute to the dissemination of blaKPC2 and/or blaNDM1 in the environment of the Egyptian hospitals, which represents a great challenge and an important factor in the dissemination of resistance and treatment failure in cases of severe infections. Agyekum et al. [39] reported that FIIK is the most common plasmid replicon found in K. pneumoniae. Another study done by Al-Marzooq et al. [40] reported that the most common plasmid replicons were IncR and IncL/M. Moreover, a high incidence of conjugative FIIK (69%) and L/M plasmids (66%) found in CPKP isolates were reported in Saudi Arabia [41].
Based on the previous findings, the Inc F plasmid is the most frequently described plasmid type to be associated primarily with resistance genes in humans [42,43].
In our study, Inc L, M, and FIC plasmids were not detected in the recipient cells, indicating that these types of plasmid were non-conjugative, and their role in the spread of carbapenem resistance among CPKP isolates is not clear; it is possible that these plasmids have no role in the transfer of resistance in our area. The prevalence of certain plasmid types is different even among samples collected from different sources within the same city and between different countries [42].
The antibiotic sensitivity tests of transconjugants showed that all transconjugants were resistant to Levofloxacin-like donor cells, except for KP19 and KP25, which can be explained by the co-existence and the transfer of quinolone resistant genes with carbapenem resistant determinants in the same strain [8].
Based on the previous results, diverse clones of multidrug-resistant (MDR) K. pneumoniae can spread between patients and the environment in the same hospital, especially with the increase of the number of patients and the inappropriate application of infection control guidelines among hospitals in Egypt. Therefore, the horizontal transmission of carbapenem-resistant plasmids among admitted patients may encourage the dissemination of carbapenem resistance to other species of Enterobacteriaceae, thereby leading to the maintenance of carbapenem-resistant clones in patients and/or the environment [7,40]. As a result, it is necessary to limit the spread of CPKP carrying highly conjugative Inc F plasmid types within healthcare settings and to implement effective infection control measures.
To the best of our knowledge, this is the first report on the identification of the conjugative Inc F plasmid that circulates and spreads in NDM-1 and KPC-2-producing CPKP isolates from a single hospital in Egypt.
4. Methods
4.1. Klebsiella pneumoniae Isolation and Identification
One hundred and twenty-six Klebsiella pneumoniae were isolated from patients admitted to different departments at Assiut University Hospital over a period of 14 months, from April 2018 to May 2019. Klebsiella pneumoniae were isolated from endotracheal aspirate samples, sputum samples, blood samples, urine samples, wound swabs, and throat swabs. Isolates were identified by an API20E kit (BioMerieux, Marcy L Etoile. France).
4.2. Antimicrobial Susceptibility Testing
The antimicrobial susceptibility of the isolated K. pneumoniae was tested by the Kirby-Bauer disc diffusion method according to recommendations of the clinical laboratory standards institute [27]. The following 18 commercial antimicrobial discs were used: Aztreonam (30 μg), Amoxicillin (10 μg), Amoxicillin-Clavulanic acid (2010 μg), Piperacillin (100 μg), Ceftriaxone (30 μg), Ceftazidime (30 μg), Cefoperazone (75 μg), Cefpodoxime (10 μg), Meropenem (10 μg), Imipenem (10 μg), Trimethoprim/Sulfamethoxazole (1.25/23.75 μg), Gentamycin (10 μg), Amikacin (30 μg), Nitrofurantoin (300 μg), Levofloxacin (5 μg), Tetracycline (30 μg), Cefoxitin (30 μg), and Cefepime (30 μg). The E. coli ATCC 25922 strain was used for quality control. The test was repeated twice for each isolate. In addition, the minimum inhibitory concentrations (MIC) of imipenem against the tested K. pneumoniae were tested using Imipenem E-test strips (bioMérieux, Solna, Sweden).
4.3. Detection of Carbapenemase-Producing Isolates
4.3.1. Modified Carbapenem Inactivation Methods (mCIM) for the Suspected Carbapenemase Producing Isolates
Modified carbapenem inactivation methods (mCIM) were performed as described by Pierce et al. [44] for suspected Carbapenemase Producing K. pneumoniae isolates. The inhibition zones for meropenem were determined after incubation at 35 °C for 18 or 24 h. A test is considered negative for carbapenemase production if the zone diameter is ≥19 mm and is considered positive if the zone diameter is 6 to 15 mm or features the appearance of pinpoint colonies within a 16 to 18 mm zone.
4.3.2. EDTA-Modified Carbapenem Inactivation Method (eCIM) for Detection of MβL Enzymes
If the mCIM test is positive, the EDTA-modified carbapenem inactivation method (eCIM) should be applied to differentiate between metallo-β-lactamases (MBL; Class B carbapenemases) and serine carbapenemases (Class A and D carbapenemases) in the K. pneumoniae isolates.
Bacterial isolates showing a positive mCIM test were cultured on Trypticase soy agar with sheep blood (TSAB). The EDTA-modified carbapenem inactivation method (eCIM) was performed according to Sfeir et al. [45]. The isolates were considered positive for MBL production if the zone diameter of meropenem disc increased by ≥5 mm in comparison to the zone diameter observed for the mCIM and were considered negative for MBL production if the increase in the zone diameter was ≤4 mm.
4.4. Detection of Hypermucoviscocity Phenotyping
The hypermucoviscocity phenotypes of the CPKP strains were checked by using the string test. The formation of a viscous filament of ≥5 mm was observed after stretching a Klebsiella spp colony with a loop cultured on an agar plate. CPKP strains positive on string tests may be considered hypervirulent CPKP (hvCPKP) [46].
4.5. PCR for Carbapenemase Genes and KI and K2 Genes
K. pneumoniae isolates were tested by PCR for the detection of carbapenemase genes. The primers used in this study were tested for bla-NDM1, bla-KPC2, bla-VIM1, bla-IMP1, and capsular type (wzy_K1 and wzy_K2) genes, as described previously by Poirel et al. [35]; Fang et al. [47]; Yigit et al. [18]; and Shibata et al. [48].
4.6. Plasmid Transfers
Conjugation experiments were performed with modifications according to Hardiman et al. [17]. For broth culture mating, the donor and recipient (E. coli J-53, azide resistant strain) cultures were mixed at a 1:4 ratio in fresh TSB and incubated at 37 °C without agitation for 20 h. The selection of transconjugants was done on a MacConkey agar containing azide (100 μg/mL) and meropenem (0.5 μg/mL). Transconjugants were tested against the same antibiotic discs used against the donor isolates. Then, the transconjugants were tested for bla-NDM1 and/or bla-KPC2 by PCR, as mentioned above.
4.7. PCR-Based Replicon Typing (Inc/Rep PCR) for Major Plasmid Incompatibility Groups among Klebseilla Isolates and Transconjugant DNA Lysates
Fourteen pairs of primers were obtained to perform PCR-based replicon typing (PBRT), in which the multiplex PCR for FII, FIA, and FIB was performed as previously described by Carattoli et al. [49], while 11 simplex PCRs were performed for IncL, IncM, IncT, FIC, FIIK, IncN, IncX3, IncH12, IncW, IncY, and IncA/C as described by Carattoli et al. [50]; Carattoli et al. [49]; Villa et al. [51]; Johnson et al. [52]; García-Fernández et al. [16]; and Carloni et al. [53].
5. Conclusions
Molecular typing of plasmids based on replicon typing showed that Inc FIIK and FII plasmids were predominant among isolates in our area and their transconjugants carrying blaKPC-2 and/or blaNDM-1. Also, conjugative Inc FII plasmids were found to have an important role in the spread of CPKP, and their recognition is essential to limit their spread.
Author Contributions
Data curation, R.M.A.E.-B.; formal analysis, E.R.M., H.M.H., and N.G.F.M.W.; methodology, R.M.A.E.-B., E.R.M., and N.G.F.M.W.; supervision, R.M.A.E.-B., M.Y.A., H.M.H. and N.G.F.M.W.; writing—original draft, E.R.M and M.Y.A.; writing—review and editing, R.M.A.E.-B. and H.M.H.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Acknowledgments
The authors received no financial support for the research or publication of this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Decre, D.; Verdet, C.; Emirian, A.; Le Gourrierec, T.; Petit, J.C.; Offenstadt, G.; Maury, E.; Brisse, S.; Arlet, G. Emerging severe and fatal infections due to Klebsiella pneumoniae in two university hospitals in France. J. Clin. Microbiol. 2011, 49, 3012–3014. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Li, X.; Zou, Y.; Wang, J.; Wang, J.; Namba, F.; Hiroyuki, Y.; Yu, J.; Yamauchi, Y.; Guo, C. Risk factors and pathogen profile of ventilator-associated pneumonia in a neonatal intensive care unit in China. Pediatrics Int. Off. J. Jpn. Pediatric Soc. 2011, 53, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Foglia, E.; Meier, M.D.; Elward, A. Ventilator-associated pneumonia in neonatal and pediatric intensive care unit patients. Clin. Microbiol. Rev. 2007, 20, 409–425, table of contents. [Google Scholar] [CrossRef] [PubMed]
- Everest, P. The Enterobacteria, 2nd Edition. Gut 2007, 56, 1331. [Google Scholar] [CrossRef][Green Version]
- Sydnor, E.R.M.; Perl, T.M. Hospital epidemiology and infection control in acute-care settings. Clin. Microbiol. Rev. 2011, 24, 141–173. [Google Scholar] [CrossRef]
- Hawkey, P.M.; Warren, R.E.; Livermore, D.M.; McNulty, C.A.M.; Enoch, D.A.; Otter, J.A.; Wilson, A.P.R. Treatment of infections caused by multidrug-resistant Gram-negative bacteria: Report of the British Society for Antimicrobial Chemotherapy/Healthcare Infection Society/British Infection Association Joint Working Party. J. Antimicrob. Chemother. 2018, 73, iii2–iii78. [Google Scholar] [CrossRef]
- Wang, X.; Chen, G.; Wu, X.; Wang, L.; Cai, J.; Chan, E.W.; Chen, S.; Zhang, R. Increased prevalence of carbapenem resistant Enterobacteriaceae in hospital setting due to cross-species transmission of the blaNDM-1 element and clonal spread of progenitor resistant strains. Front. Microbiol. 2015, 6, 595. [Google Scholar] [CrossRef]
- Yan, J.; Pu, S.; Jia, X.; Xu, X.; Yang, S.; Shi, J.; Sun, S.; Zhang, L. Multidrug resistance mechanisms of carbapenem resistant Klebsiella pneumoniae strains isolated in Chongqing, China. Ann. Lab. Med. 2017, 37, 398–407. [Google Scholar] [CrossRef][Green Version]
- Miriagou, V.; Cornaglia, G.; Edelstein, M.; Galani, I.; Giske, C.; Gniadkowski, M.; Malamou-Lada, E.; Martinez-Martinez, L.; Navarro, F.; Nordmann, P. Acquired carbapenemases in Gram-negative bacterial pathogens: Detection and surveillance issues. Clin. Microbiol. Infect. 2010, 16, 112–122. [Google Scholar] [CrossRef]
- El-Sweify, M.; Gomaa, N.; El-Maraghy, N.; Mohamed, H. Phenotypic Detection of Carbapenem Resistance among Klebsiella pneumoniae in Suez Canal University Hospitals, Ismailiya, Egypt. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 10–18. [Google Scholar]
- Gamal, D.; Fernández-Martínez, M.; Salem, D.; El-Defrawy, I.; Montes, L.Á.; Ocampo-Sosa, A.A.; Martínez-Martínez, L. Carbapenem-resistant Klebsiella pneumoniae isolates from Egypt containing blaNDM-1 on IncR plasmids and its association with rmtF. Int. J. Infect. Dis. 2016, 43, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Morsi, S.S. Comparative Evaluation of Phenotypic and Genotypic Methods for Detection of Carbapenemases in Clinically Significant Klebsiella pneumoniae Isolates. Egypt. J. Med Microbiol. 2016, 38, 1–8. [Google Scholar] [CrossRef]
- Carattoli, A. Plasmids and the spread of resistance. Int. J. Med Microbiol. 2013, 303, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Hawkey, P.M.; Jones, A.M. The changing epidemiology of resistance. J. Antimicrob. Chemother. 2009, 64, i3–i10. [Google Scholar] [CrossRef]
- Carattoli, A. Plasmids in Gram negatives: Molecular typing of resistance plasmids. Int. J. Med Microbiol. 2011, 301, 654–658. [Google Scholar] [CrossRef]
- García-Fernández, A.; Villa, L.; Carta, C.; Venditti, C.; Giordano, A.; Venditti, M.; Mancini, C.; Carattoli, A. Klebsiella pneumoniae ST258 producing KPC-3 identified in Italy carries novel plasmids and OmpK36/OmpK35 porin variants. Antimicrob. Agents Chemother. 2012, 56, 2143–2145. [Google Scholar] [CrossRef]
- Hardiman, C.; Weingarten, R.; Conlan, S.; Khil, P.; Dekker, J.; Mathers, A.; Sheppard, A.; Segre, J.; Frank, K. Horizontal transfer of carbapenemase-encoding plasmids and comparison with hospital epidemiology data. Antimicrob. Agents Chemother. 2016, 60, 4910–4919. [Google Scholar] [CrossRef]
- Yigit, H.; Queenan, A.M.; Anderson, G.J.; Domenech-Sanchez, A.; Biddle, J.W.; Steward, C.D.; Alberti, S.; Bush, K.; Tenover, F.C. Novel carbapenem-hydrolyzing β-lactamase, KPC-1, from a carbapenem-resistant strain of Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2001, 45, 1151–1161. [Google Scholar] [CrossRef]
- Nordmann, P.; Poirel, L.; Walsh, T.R.; Livermore, D.M. The emerging NDM carbapenemases. Trends Microbiol. 2011, 19, 588–595. [Google Scholar] [CrossRef]
- Bottery, M.J.; Wood, A.J.; Brockhurst, M.A. Selective Conditions for a Multidrug Resistance Plasmid Depend on the Sociality of Antibiotic Resistance. Antimicrob. Agents Chemother. 2016, 60, 2524–2527. [Google Scholar] [CrossRef]
- Raymond, B. Five rules for resistance management in the antibiotic apocalypse, a road map for integrated microbial management. Evol. Appl. 2019. [Google Scholar] [CrossRef] [PubMed]
- Kamruzzaman, M.; Shoma, S.; Thomas, C.M.; Partridge, S.R.; Iredell, J.R. Plasmid interference for curing antibiotic resistance plasmids in vivo. PLoS ONE 2017, 12, e0172913. [Google Scholar] [CrossRef] [PubMed]
- Moemen, D.; Masallat, D.T. Prevalence and characterization of carbapenem-resistant Klebsiella pneumoniae isolated from intensive care units of Mansoura University hospitals. Egypt. J. Basic Appl. Sci. 2017, 4, 37–41. [Google Scholar] [CrossRef][Green Version]
- Marquez, P.; Terashita, D.; Dassey, D.; Mascola, L. Population-based incidence of carbapenem-resistant Klebsiella pneumoniae along the continuum of care, Los Angeles County. Infect. Control Hosp. Epidemiol. 2013, 34, 144–150. [Google Scholar] [CrossRef][Green Version]
- Ashour, H.M.; El-Sharif, A. Species distribution and antimicrobial susceptibility of gram-negative aerobic bacteria in hospitalized cancer patients. J. Transl. Med. 2009, 7, 14. [Google Scholar] [CrossRef]
- Mainardi, J.-L.; Villet, R.; Bugg, T.D.; Mayer, C.; Arthur, M. Evolution of peptidoglycan biosynthesis under the selective pressure of antibiotics in Gram-positive bacteria. FEMS Microbiol. Rev. 2008, 32, 386–408. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; Clsi Suppl. M100; Clinical and Laboratory Standards Institute: Waynepa, PA, USA, 2018. [Google Scholar]
- Yu, W.-L.; Ko, W.-C.; Cheng, K.-C.; Lee, C.-C.; Lai, C.-C.; Chuang, Y.-C. Comparison of prevalence of virulence factors for Klebsiella pneumoniae liver abscesses between isolates with capsular K1/K2 and non-K1/K2 serotypes. Diagn. Microbiol. Infect. Dis. 2008, 62, 1–6. [Google Scholar] [CrossRef]
- Bialek-Davenet, S.; Criscuolo, A.; Ailloud, F.; Passet, V.; Jones, L.; Delannoy-Vieillard, A.-S.; Garin, B.; Le Hello, S.; Arlet, G.; Nicolas-Chanoine, M.-H. Genomic definition of hypervirulent and multidrug-resistant Klebsiella pneumoniae clonal groups. Emerg. Infect. Dis. 2014, 20, 1812. [Google Scholar] [CrossRef]
- Turton, J.F.; Perry, C.; Elgohari, S.; Hampton, C.V. PCR characterization and typing of Klebsiella pneumoniae using capsular type-specific, variable number tandem repeat and virulence gene targets. J. Med Microbiol. 2010, 59, 541–547. [Google Scholar] [CrossRef]
- Lin, Y.-T.; Wang, Y.-P.; Wang, F.-D.; Fung, C.-P. Community-onset Klebsiella pneumoniae pneumonia in Taiwan: Clinical features of the disease and associated microbiological characteristics of isolates from pneumonia and nasopharynx. Front. Microbiol. 2015, 6, 122. [Google Scholar] [CrossRef]
- Abdelaziz, M.O.; Bonura, C.; Aleo, A.; Fasciana, T.; Mammina, C. NDM-1-and OXA-163-producing Klebsiella pneumoniae isolates in Cairo, Egypt, 2012. J. Glob. Antimicrob. Resist. 2013, 1, 213–215. [Google Scholar] [CrossRef] [PubMed]
- Zafer, M.M.; Amin, M.; El Mahallawy, H.; Ashour, M.S.E.-D.; Al Agamy, M. First report of NDM-1-producing Pseudomonas aeruginosa in Egypt. Int. J. Infect. Dis. 2014, 29, 80–81. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, M.A.E.-G.; Amin, M.A.; Tawakol, W.M.; Loucif, L.; Bakour, S.; Rolain, J.-M. High prevalence of blaNDM-1 carbapenemase-encoding gene and 16S rRNA armA methyltransferase gene among Acinetobacter baumannii clinical isolates in Egypt. Antimicrob. Agents Chemother. 2015, 59, 3602–3605. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Dortet, L.; Bernabeu, S.; Nordmann, P. Genetic features of blaNDM-1-positive Enterobacteriaceae. Antimicrobial agents and chemotherapy 2011, 55, 5403–5407. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Benouda, A.; Hays, C.; Nordmann, P. Emergence of NDM-1-producing Klebsiella pneumoniae in Morocco. J. Antimicrob. Chemother. 2011, 66, 2781–2783. [Google Scholar] [CrossRef]
- Shen, P.; Zhang, Y.; Li, G.; Jiang, X. Characterization of the genetic environment of the bla KPC-2 gene among Klebsiella pneumoniae isolates from a Chinese hospital. Braz. J. Infect. Dis. 2016, 20, 384–388. [Google Scholar] [CrossRef]
- Jin, Y.; Shao, C.; Li, J.; Fan, H.; Bai, Y.; Wang, Y. Outbreak of multidrug resistant NDM-1-producing Klebsiella pneumoniae from a neonatal unit in Shandong Province, China. PLoS ONE 2015, 10, e0119571. [Google Scholar] [CrossRef]
- Agyekum, A.; Fajardo-Lubián, A.; Ai, X.; Ginn, A.N.; Zong, Z.; Guo, X.; Turnidge, J.; Partridge, S.R.; Iredell, J.R. Predictability of phenotype in relation to common β-lactam resistance mechanisms in Escherichia coli and Klebsiella pneumoniae. J. Clin. Microbiol. 2016, 54, 1243–1250. [Google Scholar] [CrossRef]
- Al-Marzooq, F.; Mohd Yusof, M.Y.; Tay, S.T. Molecular Analysis of Antibiotic Resistance Determinants and Plasmids in Malaysian Isolates of Multidrug Resistant Klebsiella pneumoniae. PLoS ONE 2015, 10, e0133654. [Google Scholar] [CrossRef]
- uz Zaman, T.; Alrodayyan, M.; Albladi, M.; Aldrees, M.; Siddique, M.I.; Aljohani, S.; Balkhy, H.H. Clonal diversity and genetic profiling of antibiotic resistance among multidrug/carbapenem-resistant Klebsiella pneumoniae isolates from a tertiary care hospital in Saudi Arabia. BMC Infect. Dis. 2018, 18, 205. [Google Scholar] [CrossRef]
- Rozwandowicz, M.; Brouwer, M.; Fischer, J.; Wagenaar, J.; Gonzalez-Zorn, B.; Guerra, B.; Mevius, D.; Hordijk, J. Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 1121–1137. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.; Machado, E.; Ramos, H.; Peixe, L.; Novais, Â. Expansion of ESBL-producing Klebsiella pneumoniae in hospitalized patients: A successful story of international clones (ST15, ST147, ST336) and epidemic plasmids (IncR, IncFIIK). Int. J. Med Microbiol. 2014, 304, 1100–1108. [Google Scholar] [CrossRef] [PubMed]
- Pierce, V.M.; Simner, P.J.; Lonsway, D.R.; Roe-Carpenter, D.E.; Johnson, J.K.; Brasso, W.B.; Bobenchik, A.M.; Lockett, Z.C.; Charnot-Katsikas, A.; Ferraro, M.J.; et al. Modified Carbapenem Inactivation Method for Phenotypic Detection of Carbapenemase Production among Enterobacteriaceae. J. Clin. Microbiol. 2017, 55, 2321–2333. [Google Scholar] [CrossRef] [PubMed]
- Sfeir, M.M.; Hayden, J.A.; Fauntleroy, K.A.; Mazur, C.; Johnson, J.K.; Simner, P.J.; Das, S.; Satlin, M.J.; Jenkins, S.G.; Westblade, L.F. EDTA-Modified Carbapenem Inactivation Method: A Phenotypic Method for Detecting Metallo-β-Lactamase-Producing Enterobacteriaceae. J. Clin. Microbiol. 2019, 57, e01757-18. [Google Scholar] [CrossRef] [PubMed]
- Shon, A.S.; Bajwa, R.P.; Russo, T.A. Hypervirulent (hypermucoviscous) Klebsiella pneumoniae: A new and dangerous breed. Virulence 2013, 4, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.-T.; Lai, S.-Y.; Yi, W.-C.; Hsueh, P.-R.; Liu, K.-L.; Chang, S.-C. Klebsiella pneumoniae genotype K1: An emerging pathogen that causes septic ocular or central nervous system complications from pyogenic liver abscess. Clin. Infect. Dis. 2007, 45, 284–293. [Google Scholar] [CrossRef]
- Shibata, N.; Doi, Y.; Yamane, K.; Yagi, T.; Kurokawa, H.; Shibayama, K.; Kato, H.; Kai, K.; Arakawa, Y. PCR typing of genetic determinants for metallo-β-lactamases and integrases carried by gram-negative bacteria isolated in Japan, with focus on the class 3 integron. J. Clin. Microbiol. 2003, 41, 5407–5413. [Google Scholar] [CrossRef]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 2005, 63, 219–228. [Google Scholar] [CrossRef]
- Carattoli, A.; Seiffert, S.N.; Schwendener, S.; Perreten, V.; Endimiani, A. Differentiation of IncL and IncM Plasmids Associated with the Spread of Clinically Relevant Antimicrobial Resistance. PLoS ONE 2015, 10, e0123063. [Google Scholar] [CrossRef]
- Villa, L.; Garcia-Fernandez, A.; Fortini, D.; Carattoli, A. Replicon sequence typing of IncF plasmids carrying virulence and resistance determinants. J. Antimicrob. Chemother. 2010, 65, 2518–2529. [Google Scholar] [CrossRef]
- Johnson, T.J.; Bielak, E.M.; Fortini, D.; Hansen, L.H.; Hasman, H.; Debroy, C.; Nolan, L.K.; Carattoli, A. Expansion of the IncX plasmid family for improved identification and typing of novel plasmids in drug-resistant Enterobacteriaceae. Plasmid 2012, 68, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Carloni, E.; Andreoni, F.; Omiccioli, E.; Villa, L.; Magnani, M.; Carattoli, A. Comparative analysis of the standard PCR-Based Replicon Typing (PBRT) with the commercial PBRT-KIT. Plasmid 2017, 90, 10–14. [Google Scholar] [CrossRef] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).