Photodynamic Inactivation of Candida albicans in Blood Plasma and Whole Blood


 ,
,  and
and 
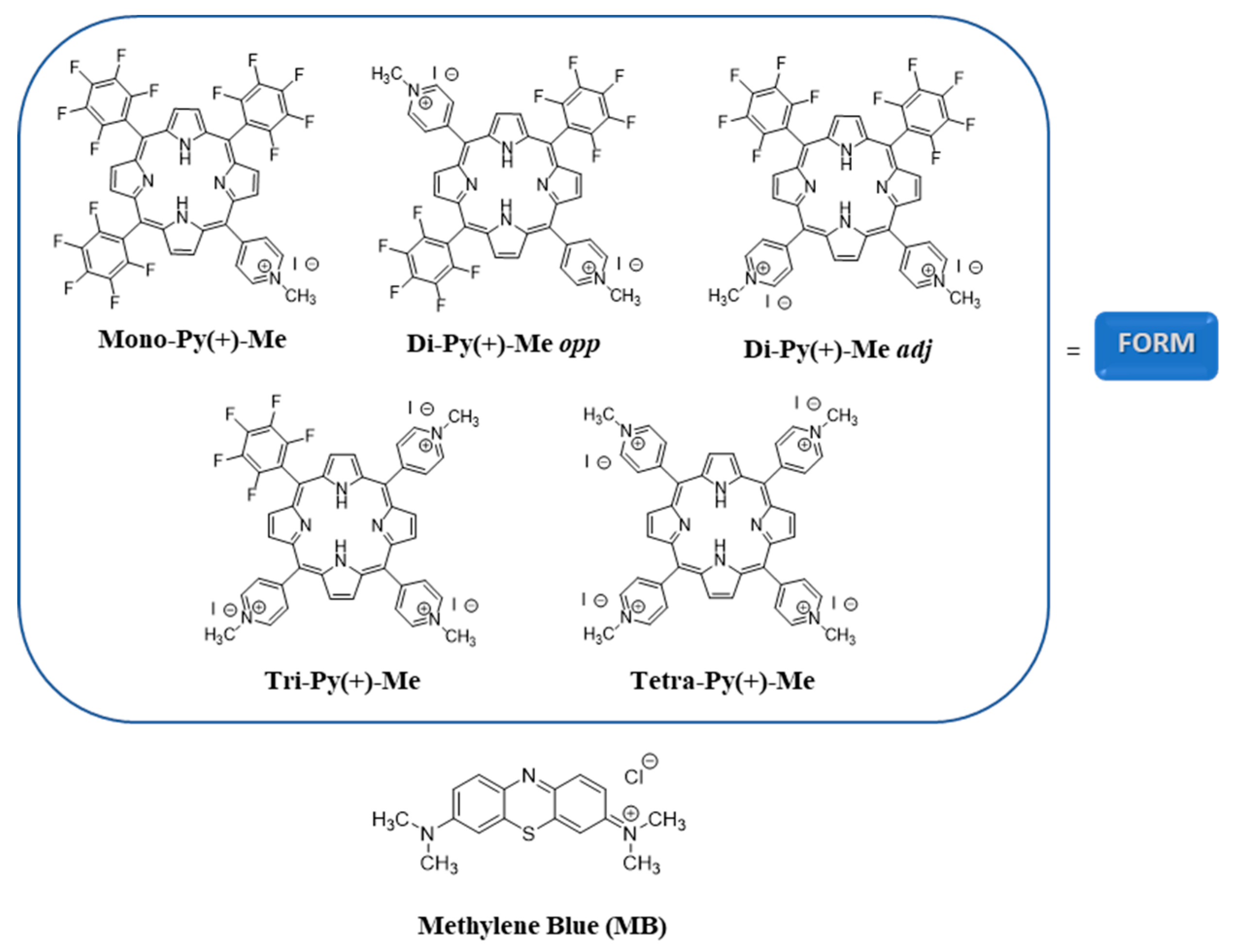{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Photodynamic Efficiency of FORM, Tri-Py(+)-Me, Tetra-Py(+)-Me, and MB in the Inactivation of C. albicans in PBS
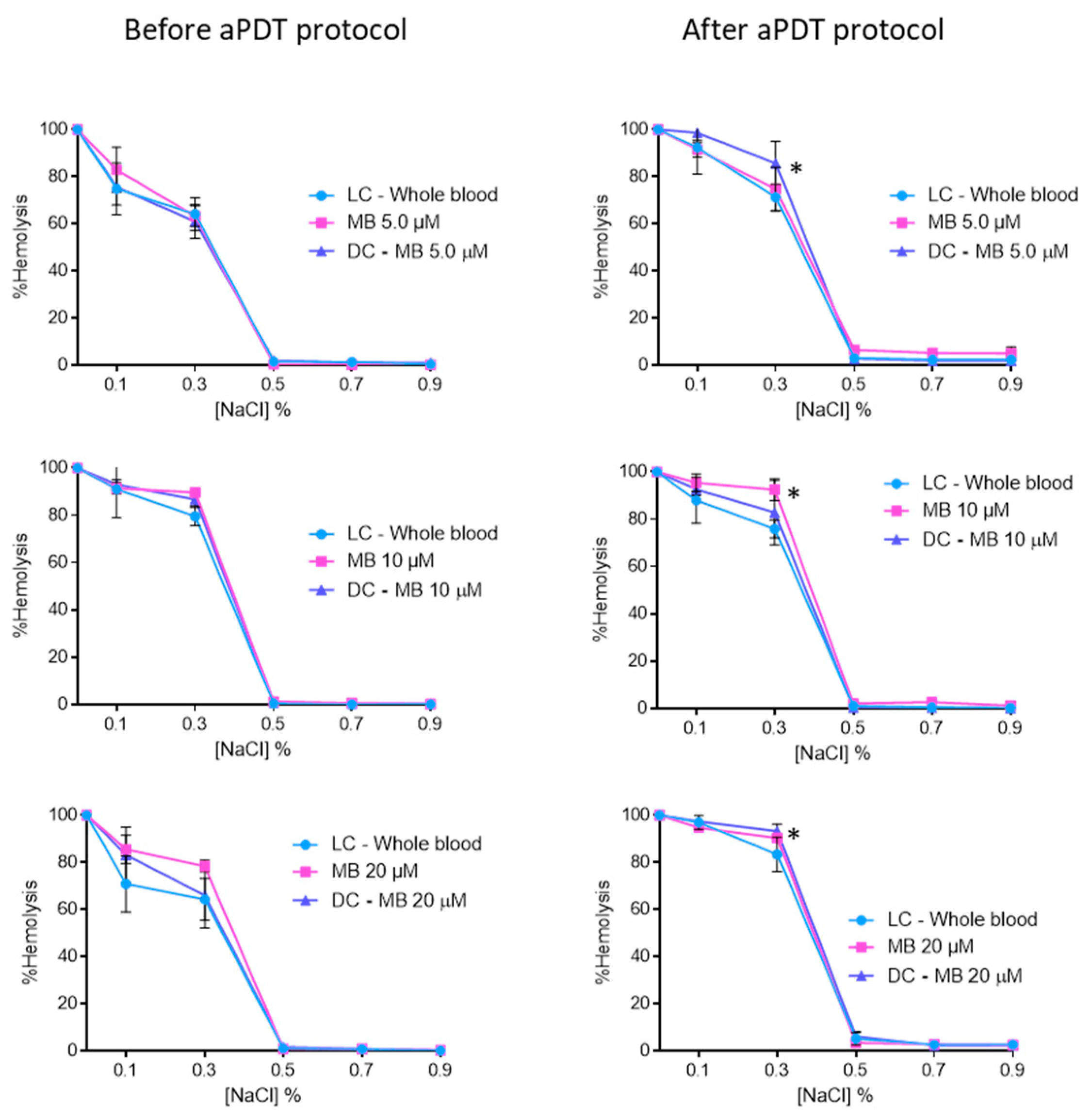
2.2. Evaluation of aPDT Effect on Erythrocyte Osmotic Fragility
2.3. aPDT of C. albicans in Blood Plasma using FORM, Tri-Py(+)-Me, and MB
2.4. aPDT of C. albicans in Whole Blood using FORM and Tri-Py-(+)-Me
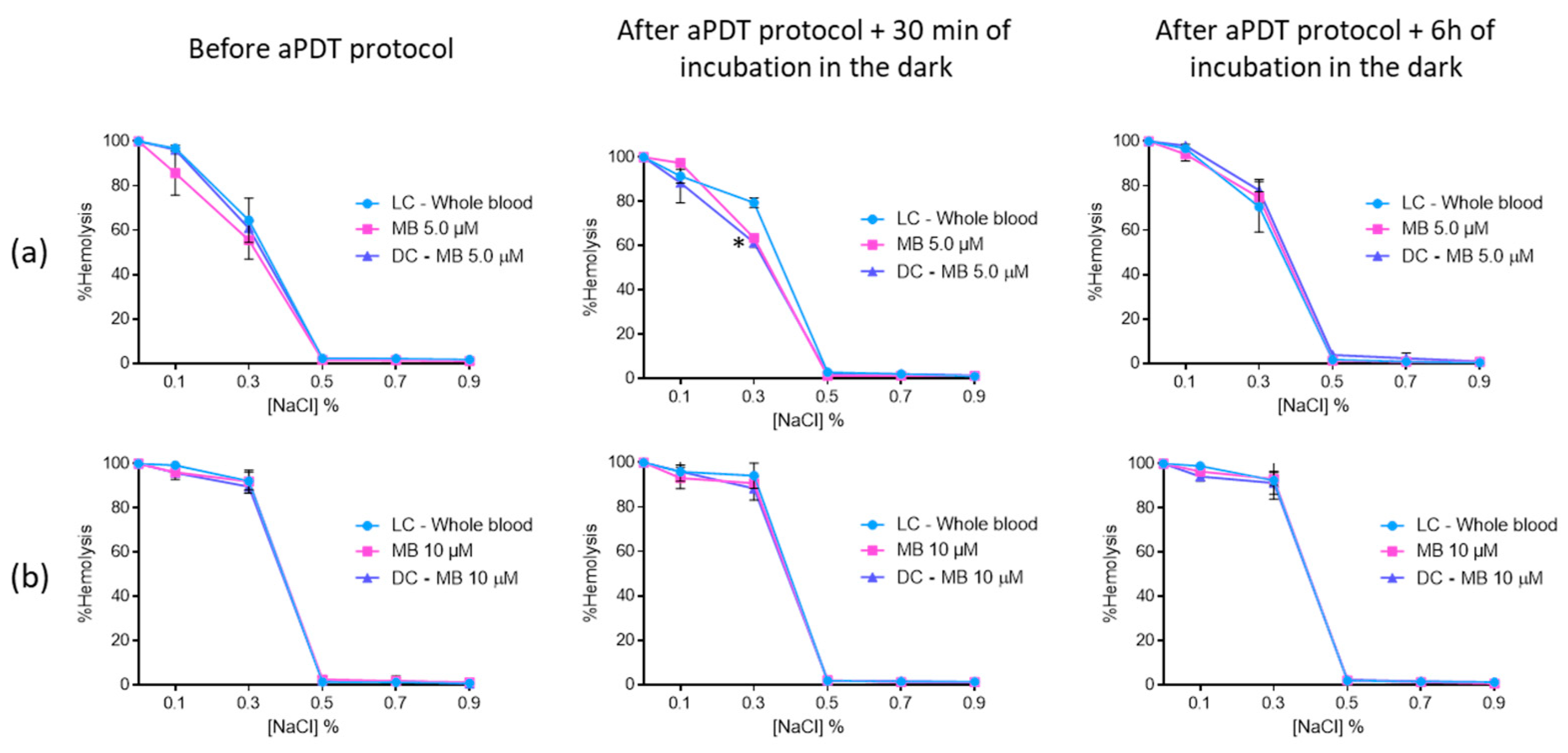
2.5. Evaluation of Erythrocyte Osmotic Fragility after the Addition of the Treated Plasma with FORM, Tri-Py(+)-Me and MB to the Concentrated Erythrocytes
3. Discussion
4. Materials and Methods
4.1. Blood Samples
4.2. Characterization of Microbial Strains and Culture Conditions
4.3. Light Sources
4.4. Photosensitizers
4.5. Antimicrobial Photodynamic Assays
4.6. Evaluation of aPDT and DMSO Effect on Erythrocyte Osmotic Fragility
4.7. Evaluation of Erythrocyte Osmotic Fragility after the Addition of the Treated Plasma with FORM, Tri(+)-Py-Me, and MB to the Concentrated Erythrocytes
4.8. Statistical
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mairbäurl, H.; Weber, R.E. Oxygen transport by hemoglobin. Compr. Physiol. 2012, 2, 1463–1489. [Google Scholar] [PubMed]
- Optical Blood Use Project. Manual Para Uso Óptimo do Sangue; Optical Blood Use Project: EU Countries, 2010. [Google Scholar]
- Chiras, D.D. The Blood. In Human Biology; Jones & Bartlett Learning: Woods Hole, MA, USA, 2019; pp. 1–711. [Google Scholar]
- Liumbruno, G.; Bennardello, F.; Lattanzio, A.; Piccoli, P.; Rossetti, G. Recommendations for the transfusion of red blood cells. Blood Transfus. 2009, 7, 49–64. [Google Scholar] [PubMed]
- Marciel, L.S.C.; Teles, L.; Moreira, B.; Pacheco, M.; Lourenço, L.M.; Neves, M.G.; Tomé, J.P.; Faustino, M.A.; Almeida, A.; Faustino, M.A. An effective and potentially safe blood disinfection protocol using tetrapyrrolic photosensitizers. Future Med. Chem. 2017, 9, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.F.; Wallington, T.; Kelsey, P.; Boulton, F.; Bruce, M.; Cohen, H.; Duguid, J.; Knowles, S.M.; Poole, G.; Williamson, L.M.; et al. Guidelines for the clinical use of red cell transfusions. Br. J. Haematol. 2001, 113, 24–31. [Google Scholar]
- Wainwright, M. Pathogen inactivation in blood products. Curr. Med. Chem. 2002, 9, 127–143. [Google Scholar] [CrossRef]
- Yaddanapudi, S.; Yaddanapudi, L. Indications for blood and blood product transfusion. Indian J. Anaesth. 2014, 58, 538–542. [Google Scholar] [CrossRef]
- Shander, A.; Goodnough, L.T. Update on Transfusion Medicine. Pharmacotherapy 2007, 27, 57S–68S. [Google Scholar] [CrossRef]
- Wainwright, M.; Mohr, H.; Walker, W.H. Phenothiazinium derivatives for pathogen inactivation in blood products. J. Photochem. Photobiol. B Biol. 2007, 86, 45–58. [Google Scholar] [CrossRef]
- Seghatchian, J.; De Sousa, G. Pathogen-reduction systems for blood components: The current position and future trends. Transfus. Apher. Sci. 2006, 35, 189–196. [Google Scholar] [CrossRef]
- Salunkhe, V.; Van Der Meer, P.F.; De Korte, D.; Seghatchian, J.; Gutiérrez, L. Development of blood transfusion product pathogen reduction treatments: A review of methods, current applications and demands. Transfus. Apher. Sci. 2015, 52, 19–34. [Google Scholar] [CrossRef]
- Bihl, F.; Castelli, D.; Marincola, F.; Dodd, R.Y.; Brander, C. Transfusion-transmitted infections. J. Transl. Med. 2007, 5, 25. [Google Scholar] [CrossRef] [PubMed]
- Seed, C.R.; Maloney, R.; Kiely, P.; Bell, B.; Keller, A.J.; Pink, J. Blood Service Medical Services Lookback Team. Infectivity of blood components from donors with occult hepatitis B infection—Results from an Australian lookback programme. Vox Sang. 2015, 108, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Hillyer, C.D.; Josephson, C.D.; Blajchman, M.A.; Vostal, J.G.; Epstein, J.S.; Goodman, J.L. Bacterial Contamination of Blood Components: Risks, Strategies, and Regulation. Am. Soc. Hematol. 2003, 2003, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Domanović, D.; Cassini, A.; Bekeredjian-Ding, I.; Bokhorst, A.; Bouwknegt, M.; Facco, G.; Galea, G.; Grossi, P.; Jashari, R.; Jungbauer, C.; et al. Prioritizing of bacterial infections transmitted through substances of human origin in Europe. Transfusion 2017, 57, 1311–1317. [Google Scholar] [CrossRef] [PubMed]
- Fred, H.L.; Thangam, M.; Aisenberg, G.M. Pathogens transmitted in red blood cell transfusions: An up-to-date table. Bayl. Univ. Med. Cent. Proc. 2018, 31, 307–309. [Google Scholar] [CrossRef] [PubMed]
- Casteel, M.J.; Jayaraj, K.; Gold, A.; Ball, L.M.; Sobsey, M.D. Photoinactivation of Hepatitis A Virus by Synthetic Porphyrins. Photochem. Photobiol. 2004, 80, 294–300. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Guidelines on Viral Inactivation and Removal Procedures Intended to Assure the Viral Safety of Human Blood Plasma Products; World Health Organization: Geneva, Switzerland, 2004; Volume 924, pp. 150–224. [Google Scholar]
- Harrison, E.; Stalhberger, T.; Whelan, R.; Sugrue, M.; Wingard, J.R.; Alexander, B.D.; Follett, S.A.; Bowyer, P.; Denning, D.W. Denning for the Aspergillus Technology Consortium (AsTeC). Aspergillus DNA contamination in blood collection tubes. Diagn. Microbiol. Infect. Dis. 2010, 67, 392–394. [Google Scholar] [CrossRef]
- Singh, G.; Sehgal, R. Transfusion-transmitted parasitic infections. Asian J. Transfus. Sci. 2010, 4, 73–77. [Google Scholar] [CrossRef]
- Lobo, C.A.; Cursino-Santos, J.R.; Alhassan, A.; Rodrigues, M. Babesia: An Emerging Infectious Threat in Transfusion Medicine. PLoS Pathog. 2013, 9, e1003387. [Google Scholar] [CrossRef]
- Liumbruno, G.M.; Catalano, L.; Piccinini, V.; Pupella, S.; Grazzini, G. Reduction of the risk of bacterial contamination of blood components through diversion of the first part of the donation of blood and blood components. Blood Transfus. 2009, 7, 86–93. [Google Scholar]
- Brecher, M.E.; Hay, S.N. Bacterial Contamination of Blood Components. Clin. Microbiol. Rev. 2005, 18, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Heger, A.; Kannicht, C.; Römisch, J.; Svae, T.E. Normal levels of ADAMTS13 and factor H are present in the pharmaceutically licensed plasma for transfusion (Octaplas) and in the universally applicable plasma (Uniplas) in development. Vox Sang. 2007, 92, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Solheim, B.; Rollag, H.; Svennevig, J.; Arafa, O.; Fosse, E.; Bergerud, U. Viral safety of solvent/detergent-treated plasma. Blood Compon. 2000, 40, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Hellstern, P.; Solheim, G. The Use of Solvent/Detergent Treatment in Pathogen. Transfus. Med. Hemother. 2011, 38, 65–70. [Google Scholar] [CrossRef]
- Di Minno, G.; Perno, C.F.; Tiede, A.; Navarro, D.; Canaro, M.; Güertler, L.; Ironside, J.W. Current concepts in the prevention of pathogen transmission via blood/plasma-derived products for bleeding disorders. Blood Rev. 2016, 30, 35–48. [Google Scholar] [CrossRef]
- Messina, G.; Ceriale, E.; Lenzi, D.; Burgassi, S.; Azzolini, E.; Manzi, P. Environmental Contaminants in Hospital Settings and Progress in Disinfecting Techniques. Biomed Res. Int. 2013, 2013, 1–9. [Google Scholar] [CrossRef]
- Burnouf, T.; Radosevich, M. Reducing the risk of infection from plasma products: Specific preventative strategies. Blood Rev. 2000, 14, 94–110. [Google Scholar] [CrossRef]
- Cieplik, F.; Tabenski, L.; Buchalla, W.; Maisch, T. Antimicrobial photodynamic therapy for inactivation of biofilms formed by oral key pathogens. Front. Microbiol. 2014, 5, 405. [Google Scholar] [CrossRef]
- Ergaieg, K.; Chevanne, M.; Cillard, J.; Seux, R. Involvement of both Type I and Type II mechanisms in Gram-positive and Gram-negative bacteria photosensitization by a meso-substituted cationic porphyrin. Sol. Energy 2008, 82, 1107–1117. [Google Scholar] [CrossRef]
- DeRosa, M.; Crutchley, R.J. Photosensitized singlet oxygen and its applications. Coord. Chem. Rev. 2002, 233, 351–371. [Google Scholar] [CrossRef]
- Schlenke, P. Pathogen inactivation technologies for cellular blood components: An update. Transfus. Med. Hemother. 2014, 41, 309–325. [Google Scholar] [CrossRef] [PubMed]
- Dean, C.L.; Wade, J.; Roback, J.D. Transfusion-Transmitted Infections: An Update on Product Screening, Diagnostic Techniques, and the Path Ahead. J. Clin. Microbiol. 2018, 56, e00352-18. [Google Scholar] [CrossRef] [PubMed]
- Lozano, M.; Cid, J.; Müller, T.H. Plasma Treated with Methylene Blue and Light: Clinical Efficacy and Safety Profile. Transfus. Med. Rev. 2013, 27, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Solheim, B.G. Pathogen reduction of blood components. Transfus. Apher. Sci. 2008, 39, 75–82. [Google Scholar] [CrossRef]
- Baptista, M.S.; Wainwright, M. Photodynamic antimicrobial chemotherapy (PACT) for the treatment of malaria, leishmaniasis and trypanosomiasis. Braz. J. Med. Biol. Res. 2011, 44, 1–10. [Google Scholar] [CrossRef]
- Wagner, S.; Storry, J.; Mallory, D.; Stromberg, R.; Benade, L.; Friedman, L. Red cell alterations associated with virucidal methylene blue phototreatment. Transfusion 1993, 33, 30–36. [Google Scholar] [CrossRef]
- Bryant, B.J.; Klein, H.G. The Definitive Safeguard for the Blood Supply. Arch. Pathol. Lab. Med. 2007, 131, 719–733. [Google Scholar]
- Williamson, L.M.; Cardigan, R.; Prowse, C.V. Methylene blue-treated fresh-frozen plasma: What is its contribution to blood safety? Transfusion 2003, 43, 1322–1329. [Google Scholar] [CrossRef]
- Del Río-Garma, J.; Alvarez-Larrán, A.; Martínez, C.; Muncunill, J.; Castellà, D.; De La Rubia, J.; Zamora, C.; Corral, M.; Viejo, A.; Peña, F.; et al. Methylene blue-photoinactivated plasma versus quarantine fresh frozen plasma in thrombotic thrombocytopenic purpura: A multicentric, prospective cohort study. Br. J. Haematol. 2008, 143, 39–45. [Google Scholar] [CrossRef]
- Benjamin, R.J.; McCullough, J.; Mintz, P.D.; Snyder, E.; Spotnitz, W.D.; Rizzo, R.J.; Wages, D.; Lin, J.-S.; Wood, L.; Corash, L.; et al. Therapeutic efficacy and safety of red blood cells treated with a chemical process (S-303) for pathogen inactivation: A Phase III clinical trial in cardiac surgery patients. Transfusion 2005, 45, 1739–1749. [Google Scholar] [CrossRef]
- Cancelas, J.A.; Gottschall, J.L.; Rugg, N.; Graminske, S.; Schott, M.A.; North, A.; Huang, N.; Mufti, N.; Erickson, A.; Rico, S.; et al. Red blood cell concentrates treated with the amustaline (S-303) pathogen reduction system and stored for 35 days retain post-transfusion viability: Results of a two-centre study. Vox Sang. 2017, 112, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.; Cunha, A.; Faustino, M.A.; Tomé, A.C.; Neves, M.G. Porphyrins as Antimicrobial Photosensitizing Agents. Photodynamic Inactivation of Microbial Pathogens: Medical and Environmental Applications; Royal Society of Chemistry: Cambridge, UK, 2011; pp. 83–160. [Google Scholar]
- Almeida, A.; Faustino, M.A.; Tomé, J.P. Photodynamic inactivation of bacteria: Finding the effective targets. Future Med. Chem. 2015, 7, 1221–1224. [Google Scholar] [CrossRef] [PubMed]
- Simões, J.; Bezerra, A.R.; Moura, G.R.; Araújo, H.; Gut, I.; Bayes, M.; Santos, M.A.S. The Fungus Candida albicans Tolerates Ambiguity at Multiple Codons. Front. Microbiol. 2016, 7, 583. [Google Scholar] [CrossRef] [PubMed]
- Burghi, G.; Ortiz, G.; Bagnulo, H. Blood transfusions: An independent risk factor for the development of Candida infections in critically ill surgical patients. Crit. Care 2011, 15, P237. [Google Scholar] [CrossRef][Green Version]
- Troiano, G.; Mercurio, I.; Nante, N.; Lancia, M.; Bacci, M. Candida autovaccination: A new strategy to prevent antifungal resistance? J. Infect. Prev. 2018, 19, 201–202. [Google Scholar] [CrossRef]
- Douglas, L.J. Candida biofilms and their role in infection. Trends Microbiol. 2003, 11, 30–36. [Google Scholar] [CrossRef]
- Spampinato, C.; Leonardi, D. Candida Infections, Causes, Targets, and Resistance Mechanisms: Traditional and Alternative Antifungal Agents. BioMed Res. Int. 2013, 2013, 204237. [Google Scholar] [CrossRef]
- Brock, M. Fungal metabolism in host niches. Curr. Opin. Microbiol. 2009, 12, 371–376. [Google Scholar] [CrossRef]
- Azizi, A.; Amirzadeh, Z.; Rezai, M.; Lawaf, S.; Rahimi, A. Effect of photodynamic therapy with two photosensitizers on Candida albicans. J. Photochem. Photobiol. B Biol. 2016, 158, 267–273. [Google Scholar] [CrossRef]
- Vieira, C.; Santos, A.; Mesquita, M.Q.; Gomes, A.T.P.C.; Neves, M.G.P.M.S.; Faustino, M.A.F.; Almeida, A. Advances in aPDT based on the combination of a porphyrinic formulation with potassium iodide: Effectiveness on bacteria and fungi planktonic/biofilm forms and viruses. J. Porphyr. Phthalocyanines 2019, 23, 534–545. [Google Scholar] [CrossRef]
- Silva, M.P.; Dos Santos, T.A.; De Barros, P.P.; Ribeiro, F.D.C.; Junqueira, J.C.; Jorge, A.O.C. Action of antimicrobial photodynamic therapy on heterotypic biofilm: Candida albicans and Bacillus atrophaeus. Lasers Med. Sci. 2016, 31, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Carmello, J.C.; Alves, F.; Basso, F.G.; Costa, C.A.D.S.; Tedesco, A.C.; Primo, F.L.; Mima, E.G.D.O.; Pavarina, A.C. Antimicrobial photodynamic therapy reduces adhesion capacity and biofilm formation of Candida albicans from induced oral candidiasis in mice. Photodiagnosis Photodyn. Ther. 2019, 27, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Bertoloni, G.; Zambotto, F.; Conventi, L.; Reddi, E.; Jori, G. Role of specific cellular targets in the hematoporphy rin-sensitized photoinactivation of microbial cells. Photochem. Photobiol. 1987, 46, 695–698. [Google Scholar] [CrossRef] [PubMed]
- Bliss, J.M.; Bigelow, C.E.; Foster, T.H.; Haidaris, C.G. Susceptibility of Candida Species to Photodynamic Effects of Photofrin. Antimicrob. Agents Chemother. 2004, 48, 2000–2006. [Google Scholar] [CrossRef] [PubMed]
- Chabrier-Roselló, Y.; Foster, T.H.; Pérez-Nazario, N.; Mitra, S.; Haidaris, C.G. Sensitivity of Candida albicans Germ Tubes and Biofilms to Photofrin-Mediated Phototoxicity. Antimicrob. Agents Chemother. 2005, 49, 4288–4295. [Google Scholar] [CrossRef]
- Philippova, T.O.; Galkin, B.N.; Zinchenko, O.Y.; Rusakova, M.Y.; Ivanitsa, V.A.; Zhilina, Z.I.; Vodzinskii, S.V.; Ishkov, Y.V. The antimicrobial properties of new synthetic porphyrins. J. Porphyr. Phthalocyanines 2003, 7, 755–760. [Google Scholar] [CrossRef]
- Yu, K.G.; Li, D.H.; Zhou, C.H.; Diao, J.L. Study on the synthesis and antimicrobial activity of novel cationic porphyrins. Chin. Chem. Lett. 2009, 20, 411–414. [Google Scholar] [CrossRef]
- Mora, S.J.; Cormick, M.P.; Milanesio, M.E.; Durantini, E.N. The photodynamic activity of a novel porphyrin derivative bearing a fluconazole structure in different media and against Candida albicans. Dyes Pigments 2010, 87, 234–240. [Google Scholar] [CrossRef]
- Monfrecola, G.; Procaccini, E.M.; Bevilacqua, M.; Manco, A.; Calabrò, G.; Santoianni, P. In vitro effect of 5-aminolaevulinic acid plus visible light on Candida albicans. Photochem. Photobiol. Sci. 2003, 3, 419–422. [Google Scholar] [CrossRef]
- Vieira, C.; Gomes, A.T.P.C.; Mesquita, M.Q.; Moura, N.M.M.; Neves, M.G.P.M.S.; Faustino, M.A.F.; Almeida, A. An Insight into the Potentiation Effect of Potassium Iodide on aPDT Efficacy. Front. Microbiol. 2018, 9, 2665. [Google Scholar] [CrossRef]
- Marciel, L.; Mesquita, M.Q.; Ferreira, R.; Moreira, B.; Neves, M.G.P.; Faustino, M.A.F.; Almeida, A. An efficient formulation based on cationic porphyrins to photoinactivate Staphylococcus aureus and Escherichia coli. Future Med. Chem. 2018, 10, 1821–1833. [Google Scholar] [CrossRef] [PubMed]
- Martins, D.; Mesquita, M.Q.; Neves, M.G.P.M.S.; Faustino, M.A.F.; Reis, L.; Figueira, E.; Almeida, A. Photoinactivation of Pseudomonas syringae pv. actinidiae in kiwifruit plants by cationic porphyrins. Planta 2018, 248, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Kibbler, C.; Seaton, S.; Barnes, R.; Gransden, W.; Holliman, R.; Johnson, E.; Perry, J.; Sullivan, D.; Wilson, J. Management and outcome of bloodstream infections due to Candida species in England and Wales. J. Hosp. Infect. 2003, 54, 18–24. [Google Scholar] [CrossRef]
- Diogo, P.; Fernandes, C.; Caramelo, F.; Mota, M.; Miranda, I.M.; Faustino, M.A.F.; Neves, M.G.P.M.S.; Uliana, M.P.; De Oliveira, K.T.; Santos, J.M.; et al. Antimicrobial Photodynamic Therapy against Endodontic Enterococcus faecalis and Candida albicans Mono and Mixed Biofilms in the Presence of Photosensitizers: A Comparative Study with Classical Endodontic Irrigants. Front. Microbiol. 2017, 8, 626. [Google Scholar] [CrossRef]
- Fumes, A.C.; Telles, P.D.D.S.; Corona, S.A.M.; Borsatto, M.C. Effect of aPDT on Streptococcus mutans and Candida albicans present in the dental biofilm: Systematic review. Photodiagnosis Photodyn. Ther. 2018, 21, 363–366. [Google Scholar] [CrossRef]
- Carvalho, G.G.; Felipe, M.P.; Costa, M.S. The photodynamic effect of methylene blue and toluidine blue on Candida albicans is dependent on medium conditions. J. Microbiol. 2009, 47, 619–623. [Google Scholar] [CrossRef]
- Leonel, L.D.C.; Carvalho, M.L.; Da Silva, B.M.; Zamuner, S.; Alberto-Silva, C.; Costa, M.S. Photodynamic Antimicrobial Chemotherapy (PACT) using methylene blue inhibits the viability of the biofilm produced by Candida albicans. Photodiagnosis Photodyn. Ther. 2019, 26, 316–323. [Google Scholar] [CrossRef]
- Can Ince, A.B. Johan Groeneveld. The case for 0.9% NaCl: Is the undefendable, defensible? Kidney Int. 2014, 86, 1087–1095. [Google Scholar] [CrossRef]
- Alves, E.; Rodrigues, J.M.; Faustino, M.A.; Neves, M.G.; Cavaleiro, J.A.; Lin, Z.; Cunha, Â.; Nadais, M.H.; Tomé, J.P.; Almeida, A.; et al. A new insight on nanomagnet–porphyrin hybrids for photodynamic inactivation of microorganisms. Dyes Pigments 2014, 110, 80–88. [Google Scholar] [CrossRef]
- Simões, C.; Gomes, M.C.; Neves, M.G.; Cunha, Â.; Tomé, J.P.; Tomé, A.C.; Cavaleiro, J.A.; Almeida, A.; Faustino, M.A.; Neves, M.D.G.P.M.S.; et al. Photodynamic inactivation of Escherichia coli with cationic meso-tetraarylporphyrins—The charge number and charge distribution effects. Catal. Today 2016, 266, 197–204. [Google Scholar] [CrossRef]
- Mundt, J.M.; Rouse, L.; Van den Bossche, J.; Goodrich, R.P. Chemical and Biological Mechanisms of Pathogen Reduction Technologies. Photochem. Photobiol. 2014, 90, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Azeez, O.I.; Oyagbemi, A.A.; Iji, O.T. Haematology and Erythrocyte Osmotic Fragility Indices in Domestic Chicken Following Exposure to a Polyvalent Iodophorous Disinfectant. Jordan J. Biol. Sci. 2012, 5, 99–103. [Google Scholar]











© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa, V.; Gomes, A.T.P.C.; Freitas, A.; Faustino, M.A.F.; Neves, M.G.P.M.S.; Almeida, A. Photodynamic Inactivation of Candida albicans in Blood Plasma and Whole Blood. Antibiotics 2019, 8, 221. https://doi.org/10.3390/antibiotics8040221
Sousa V, Gomes ATPC, Freitas A, Faustino MAF, Neves MGPMS, Almeida A. Photodynamic Inactivation of Candida albicans in Blood Plasma and Whole Blood. Antibiotics. 2019; 8(4):221. https://doi.org/10.3390/antibiotics8040221
Chicago/Turabian StyleSousa, Vera, Ana T. P. C. Gomes, Américo Freitas, Maria A. F. Faustino, Maria G. P. M. S. Neves, and Adelaide Almeida. 2019. "Photodynamic Inactivation of Candida albicans in Blood Plasma and Whole Blood" Antibiotics 8, no. 4: 221. https://doi.org/10.3390/antibiotics8040221
APA StyleSousa, V., Gomes, A. T. P. C., Freitas, A., Faustino, M. A. F., Neves, M. G. P. M. S., & Almeida, A. (2019). Photodynamic Inactivation of Candida albicans in Blood Plasma and Whole Blood. Antibiotics, 8(4), 221. https://doi.org/10.3390/antibiotics8040221