Degradation Kinetics of Clavulanic Acid in Fermentation Broths at Low Temperatures

 , ,
, , 

Abstract
1. Introduction
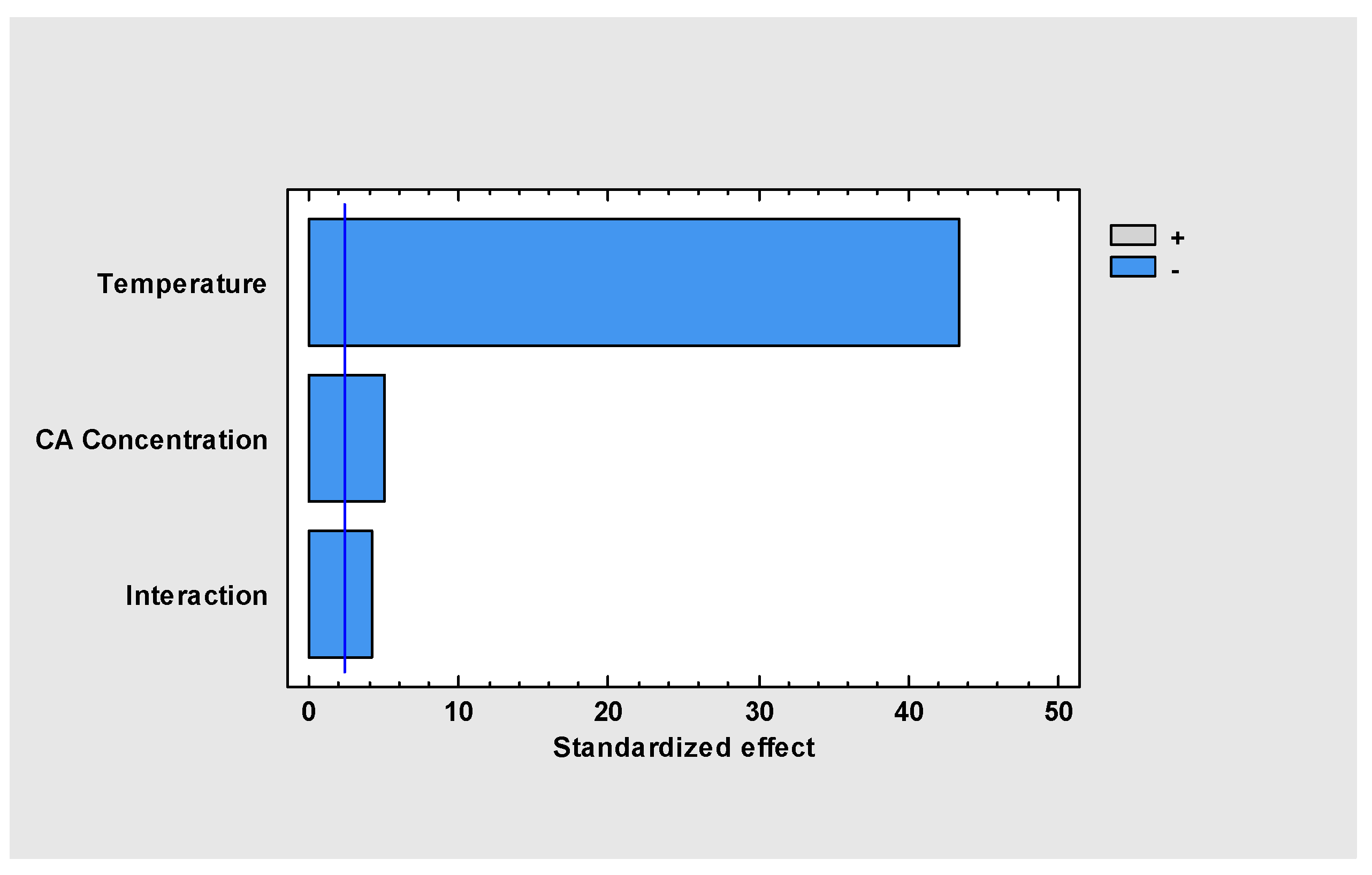
2. Results and Discussion
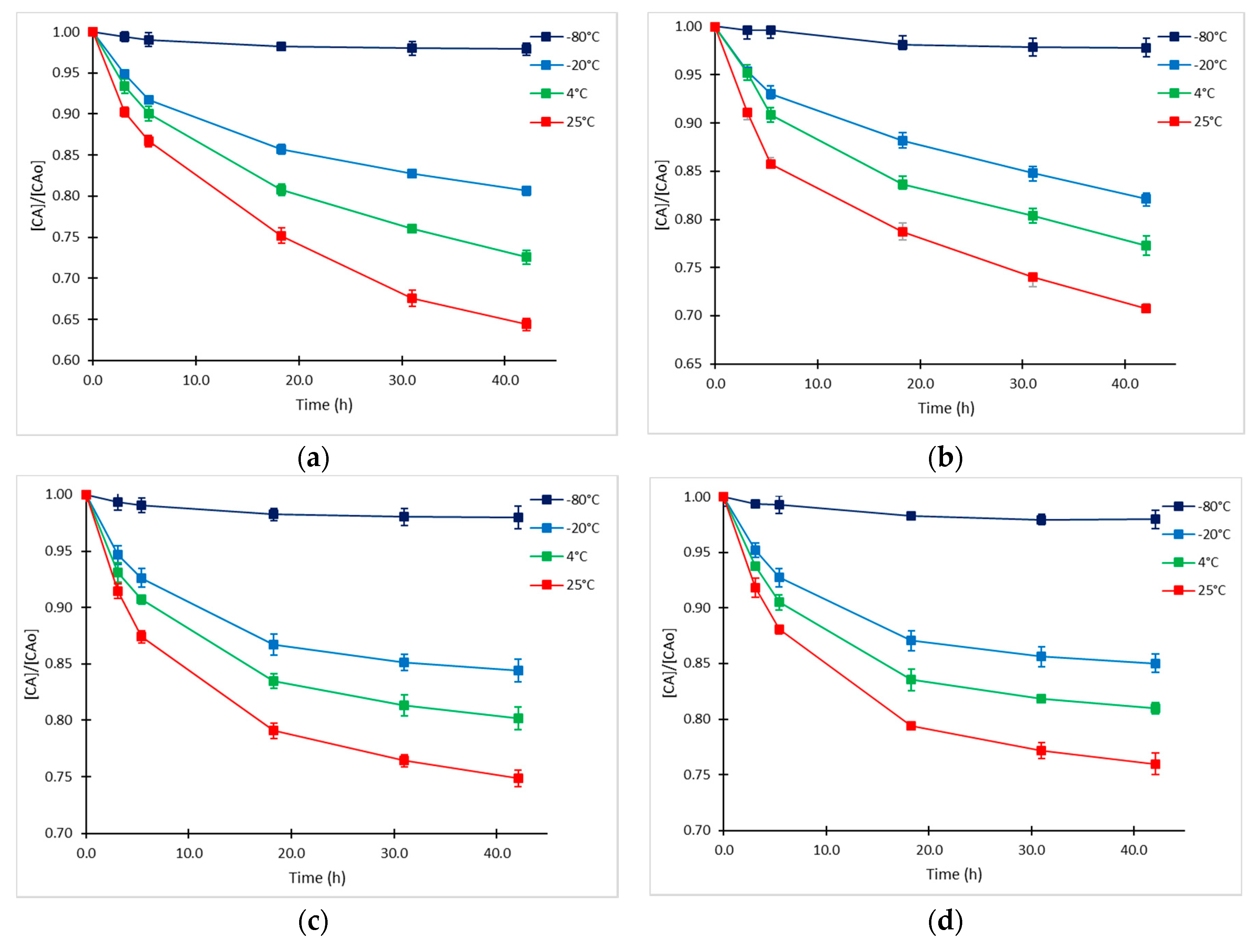
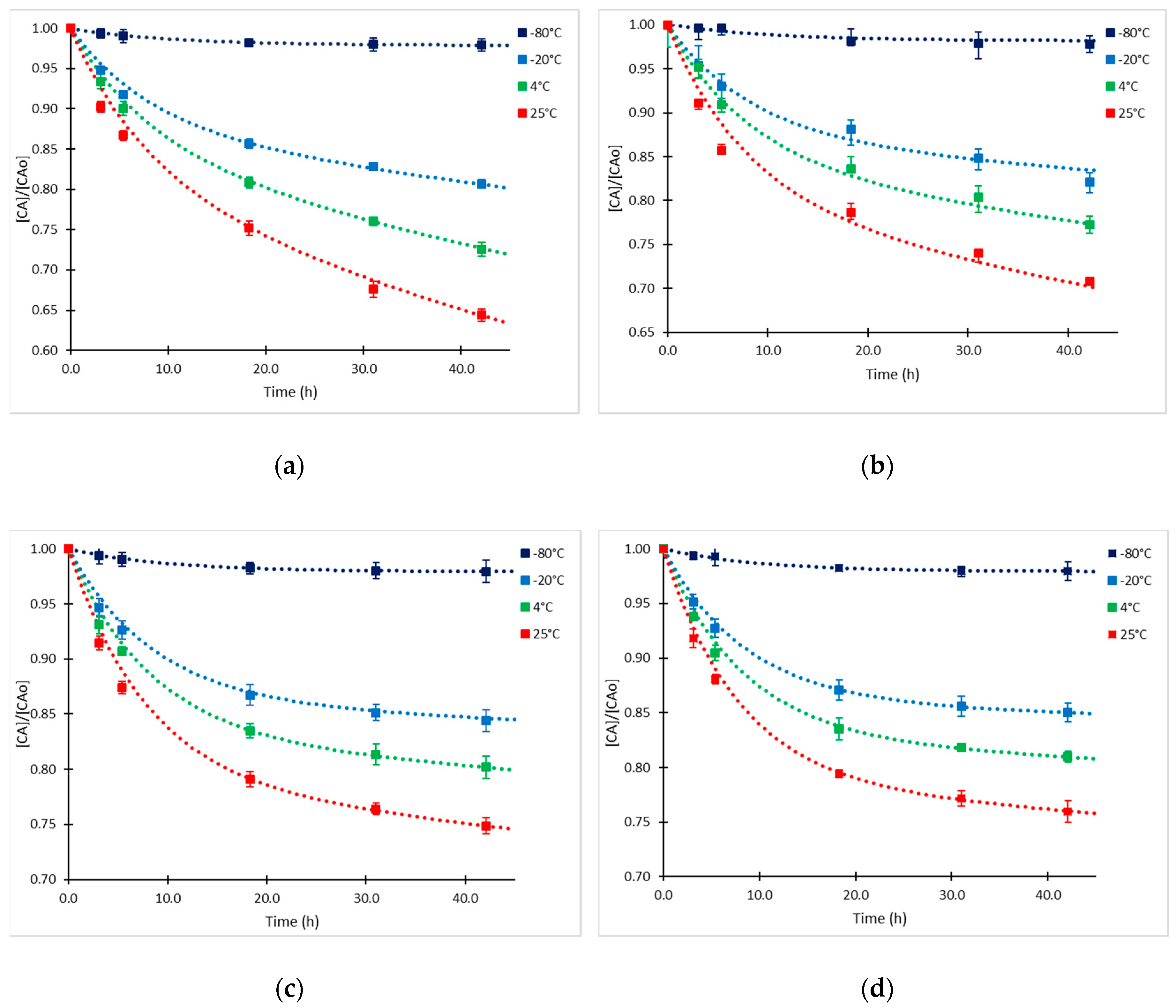
2.1. Clavulanic Acid Degradation
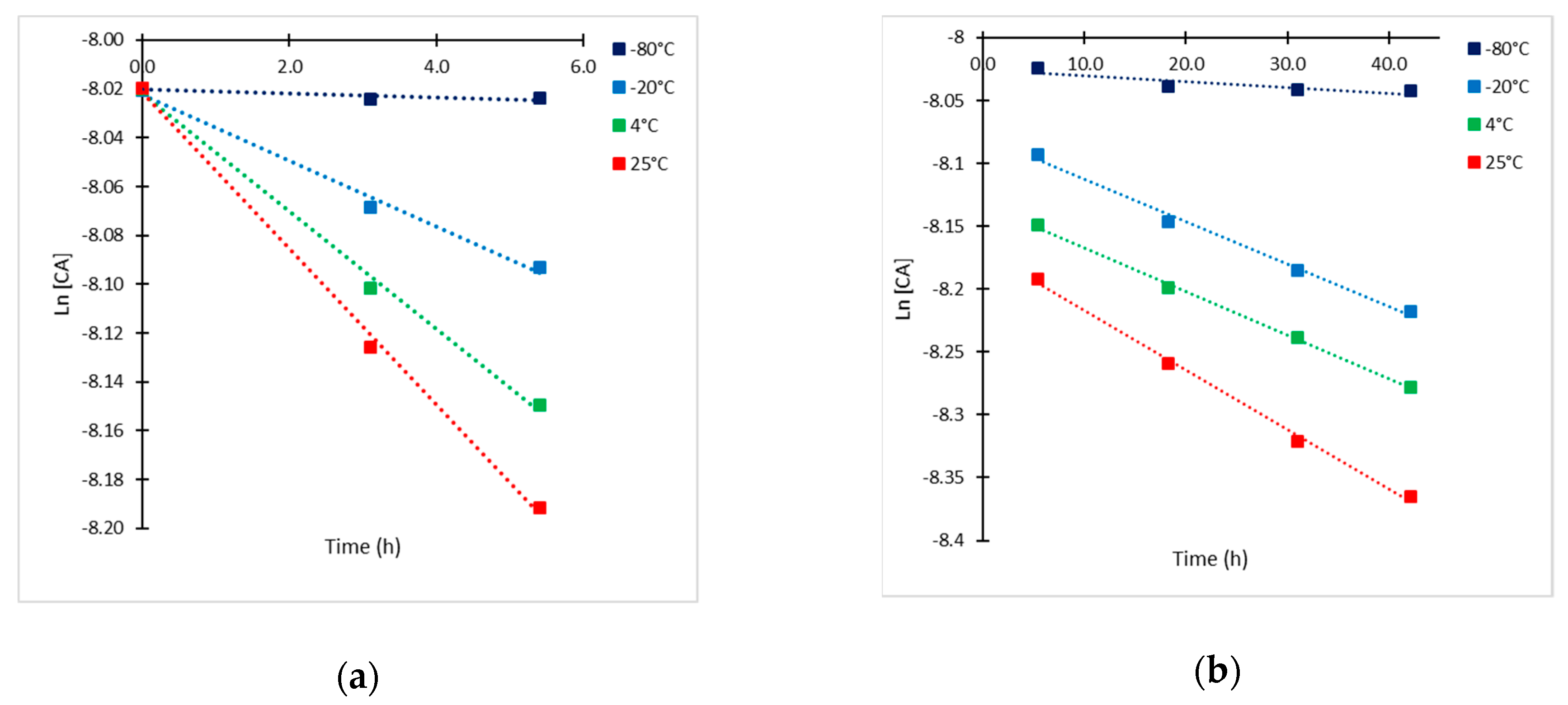
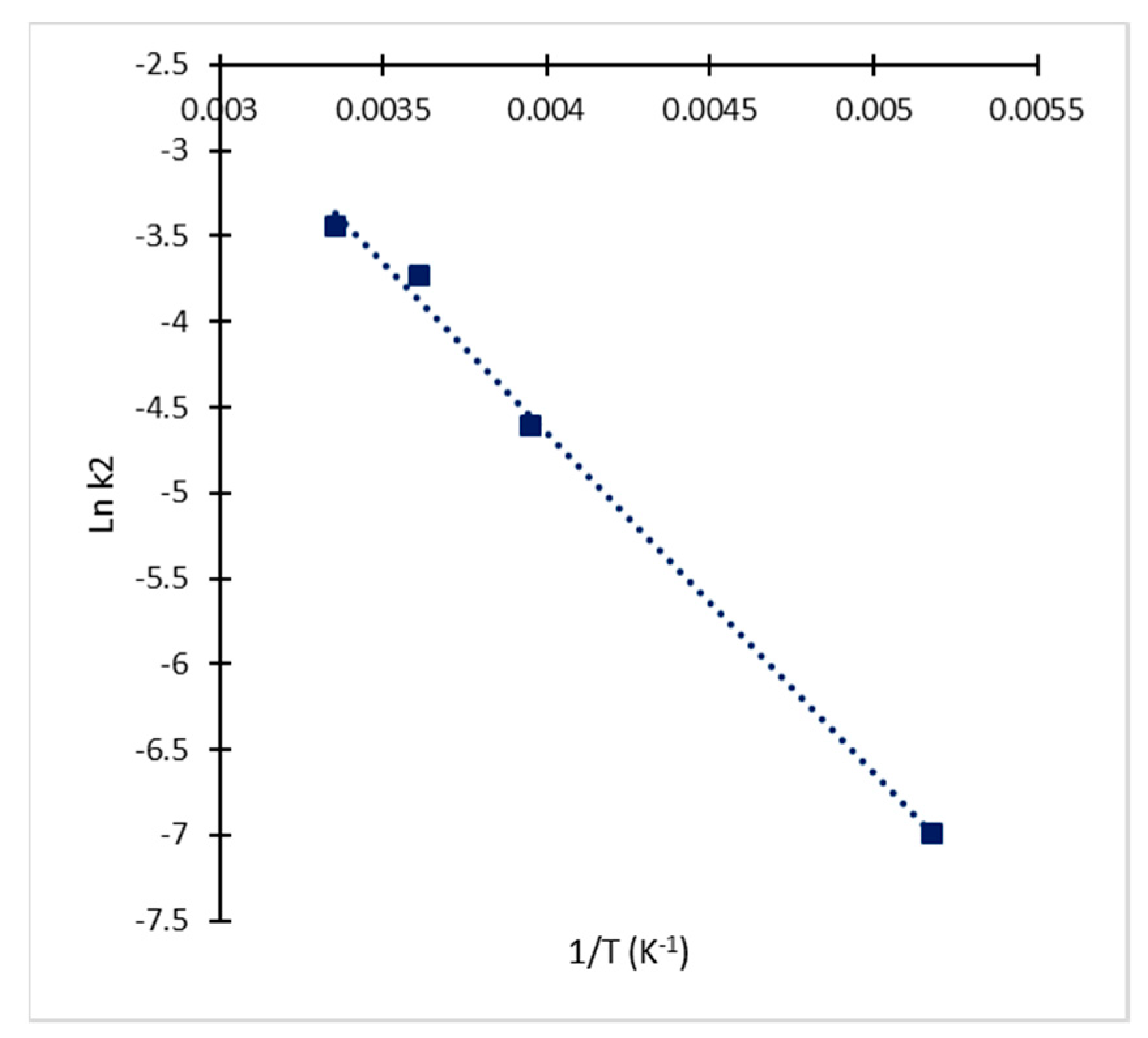
2.2. Kinetic Approach of CA Degradation
2.3. Kinetic Model Validation
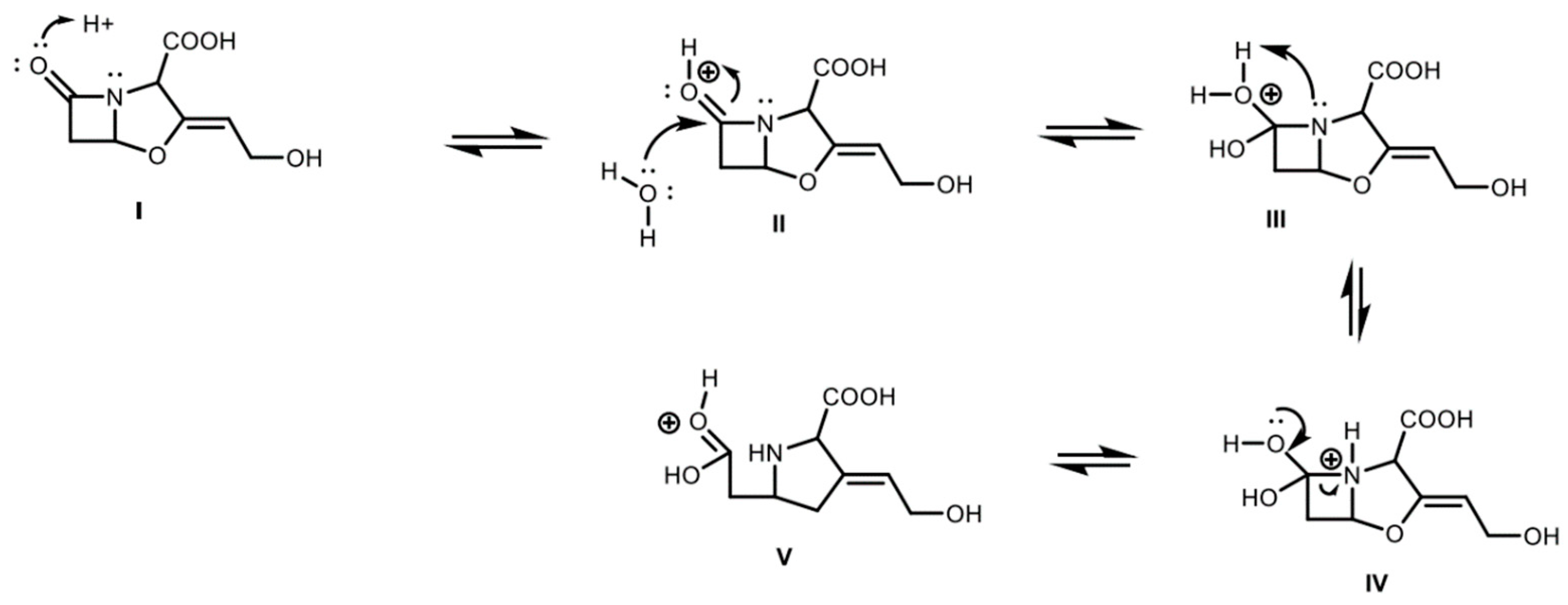
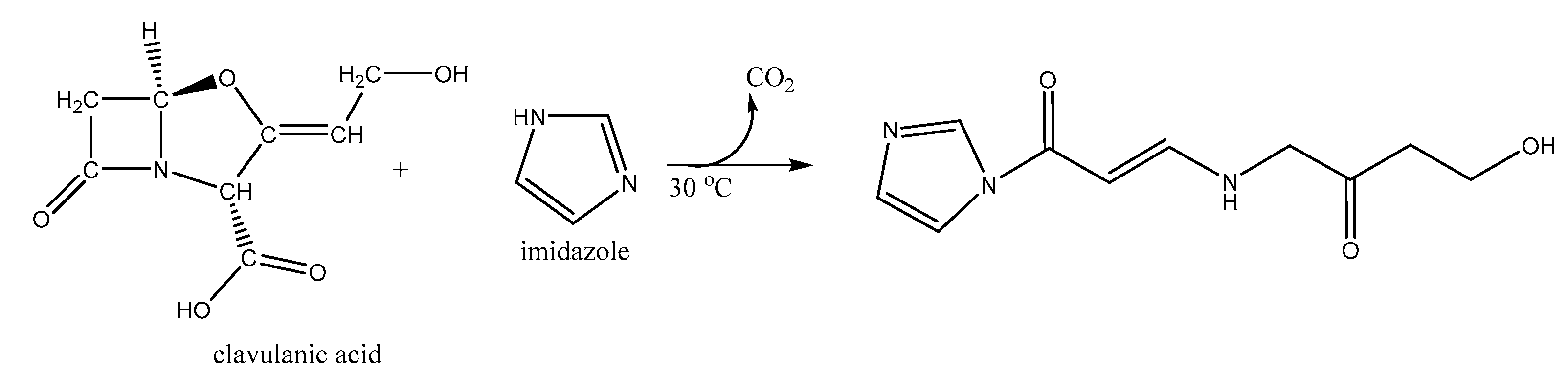
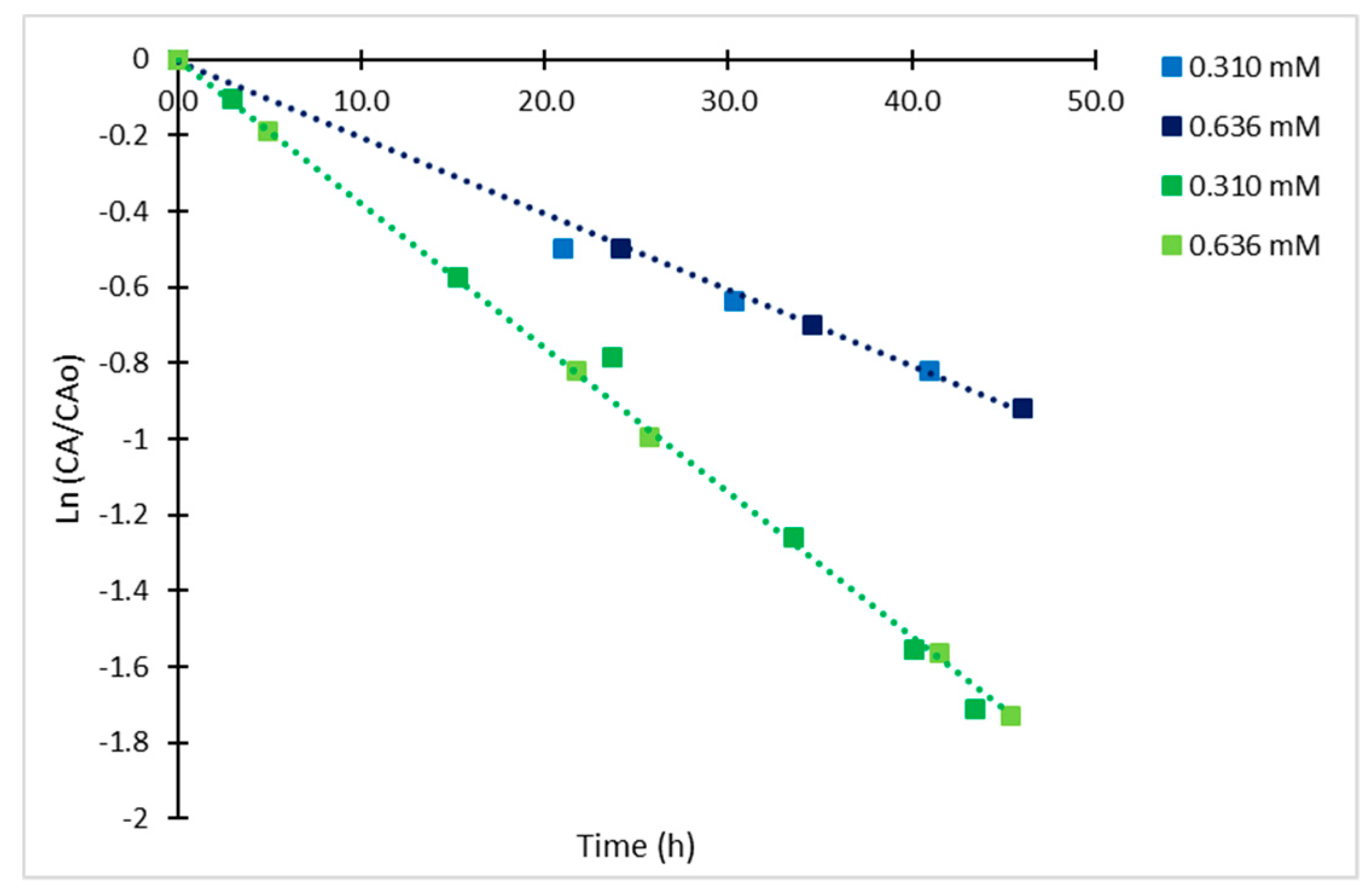
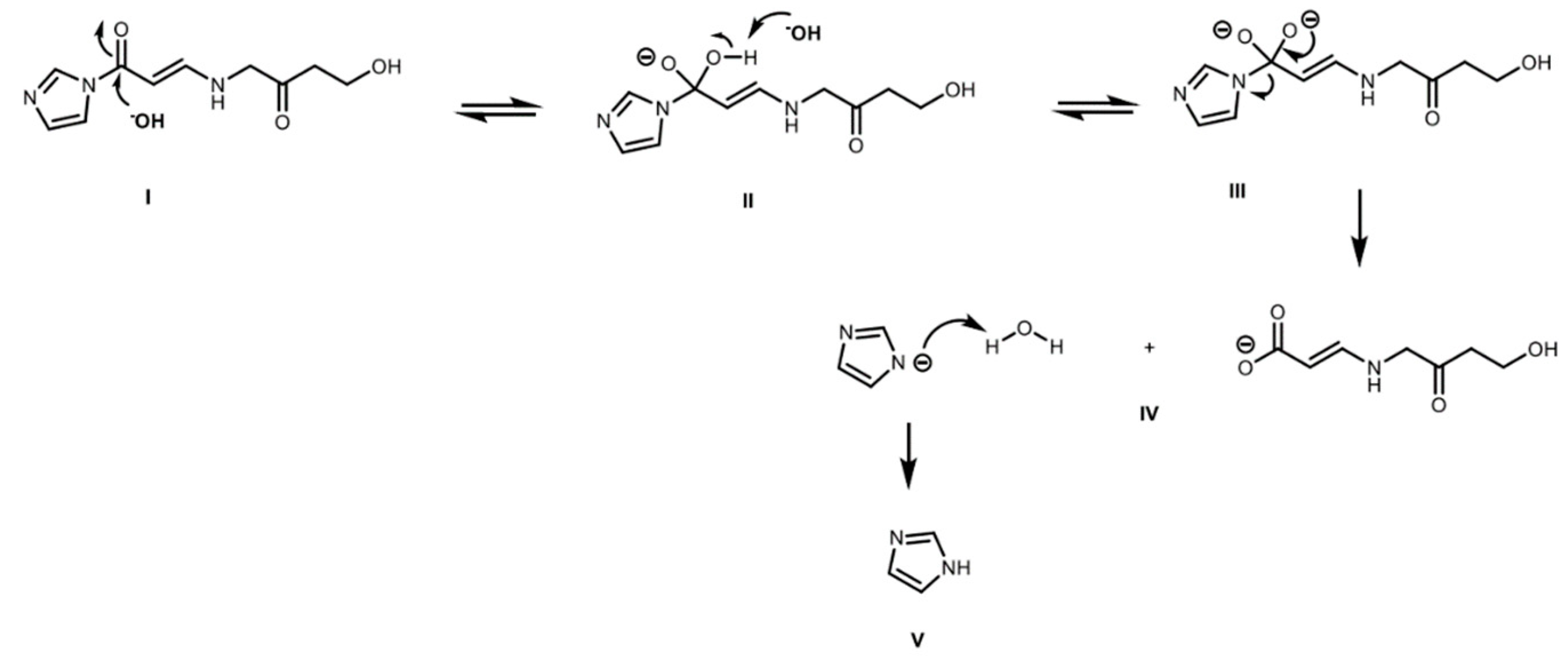
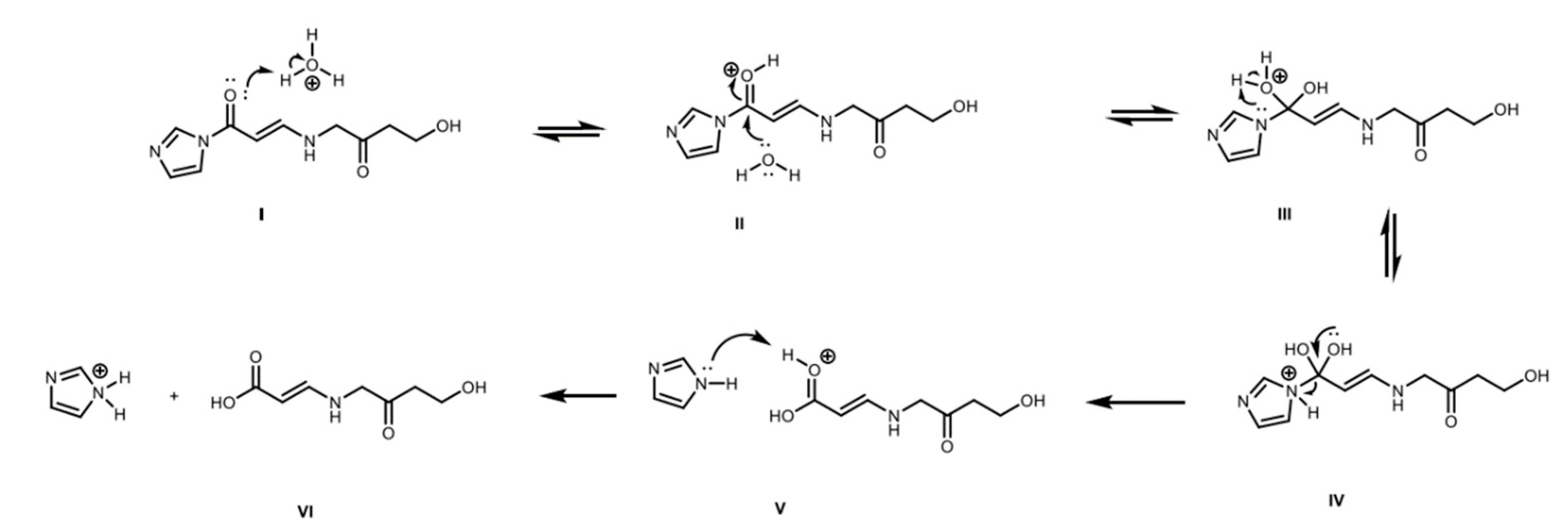
2.4. Derivatized Clavulanate-Imidazole Degradation Kinetics
3. Materials and Methods
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ramirez-Malule, H. Bibliometric analysis of global research on clavulanic acid. Antibiotics 2018, 7, 102. [Google Scholar] [CrossRef] [PubMed]
- De Araujo Viana Marques, D.; Feitosa Machado, S.E.; Santos Ebinuma, V.C.; De Albuquerque Lima Duarte, C.; Converti, A.; Porto, A.L.F. Production of β-lactamase inhibitors by Streptomyces species. Antibiotics 2018, 7, 61. [Google Scholar]
- Takahashi, Y.; Nakashima, T. Actinomycetes, an inexhaustible source of naturally occurring antibiotics. Antibiotics 2018, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Manteca, Á.; Yagüe, P. Streptomyces differentiation in liquid cultures as a trigger of Secondary metabolism. Antibiotics 2018, 7, 41. [Google Scholar] [CrossRef] [PubMed]
- Ser, H.-L.; Law, J.W.-F.; Chaiyakunapruk, N.; Jacob, S.A.; Palanisamy, U.D.; Chan, K.-G.; Goh, B.-H.; Lee, L.-H. Fermentation conditions that affect clavulanic acid production in Streptomyces clavuligerus: A systematic review. Front. Microbiol. 2016, 7, 522. [Google Scholar] [CrossRef] [PubMed]
- Saudagar, P.S.; Singhal, R.S. Optimization of nutritional requirements and feeding strategies for clavulanic acid production by Streptomyces clavuligerus. Bioresour. Technol. 2007, 98, 2010–2017. [Google Scholar] [CrossRef]
- Bellão, C.; Antonio, T.; Araujo, M.L.G.C.; Badino, A.C. Production of clavulanic acid and cephamycin c by Streptomyces clavuligerus under different fed-batch conditions. Braz. J. Chem. Eng. 2013, 30, 257–266. [Google Scholar] [CrossRef]
- Teodoro, J.C.; Baptista-Neto, A.; Araujo, M.L.G.C.; Hokka, C.O.; Badino, A.C. Influence of glycerol and ornithine feeding on clavulanic acid production by Streptomyces clavuligerus. Braz. J. Chem. Eng. 2010, 27, 499–506. [Google Scholar] [CrossRef]
- Neto, A.B.; Hirata, D.B.; Cassiano Filho, L.C.M.; Bellão, C.; Badino, A.C.; Hokka, C.O. A study on clavulanic acid production by Streptomyces clavuligerus in batch, FED-batch and continuous processes. Braz. J. Chem. Eng. 2005, 22, 557–563. [Google Scholar] [CrossRef]
- Bersanetti, P.A.; Almeida, R.M.R.G.; Barboza, M.; Araújo, M.L.G.C.; Hokka, C.O. Kinetic studies on clavulanic acid degradation. Biochem. Eng. J. 2005, 23, 31–36. [Google Scholar] [CrossRef]
- Marques, D.A.V.; Oliveira, R.P.S.; Perego, P.; Porto, A.L.F.; Pessoa, A.; Converti, A. Kinetic and thermodynamic investigation on clavulanic acid formation and degradation during glycerol fermentation by Streptomyces DAUFPE 3060. Enzyme Microb. Technol. 2009, 45, 169–173. [Google Scholar] [CrossRef]
- Rosa, J.C.; Baptista Neto, A.; Hokka, C.O.; Badino, A.C. Influence of dissolved oxygen and shear conditions on clavulanic acid production by Streptomyces clavuligerus. Bioprocess Biosyst. Eng. 2005, 27, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Teodoro, J.C.; Baptista-Neto, A.; Cruz-Hernández, I.L.; Hokka, C.O.; Badino, A.C. Influence of feeding conditions on clavulanic acid production in fed-batch cultivation with medium containing glycerol. Appl. Microbiol. Biotechnol. 2006, 72, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Bushell, M.E.; Kirk, S.; Zhao, H.; Avignone-rossa, C.A. Manipulation of the physiology of clavulanic acid biosynthesis with the aid of metabolic flux analysis. Enzyme Microb. Technol. 2006, 39, 149–157. [Google Scholar] [CrossRef]
- Cerri, M.O.; Badino, A.C. Shear conditions in clavulanic acid production by Streptomyces clavuligerus in stirred tank and airlift bioreactors. Bioprocess Biosyst. Eng. 2012, 35, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.L.L.; Badino, A.C. Production of clavulanic acid by Streptomyces clavuligerus in batch cultures without and with glycerol pulses under different temperature conditions. Biochem. Eng. J. 2012, 69, 1–7. [Google Scholar] [CrossRef]
- Carvalho, V.; Brandão, J.F.; Brandão, R.; Rangel-yagui, C.O.; Couto, J.A.; Converti, A.; Pessoa, A. Stability of clavulanic acid under variable pH, ionic strength and temperature conditions. A new kinetic approach. Biochem. Eng. J. 2009, 45, 89–93. [Google Scholar]
- Ishida, K.; Hung, T.V.; Lee, H.C.; Liou, K.K.; Shin, C.H.; Yoon, Y.J.; Sohng, J.K. Degradation of clavulanic Acid During the cultivation of Streptomyces clavuligerus; Instability of clavulanic Acid by Metabolites and Proteins from the Strain. J. Microbiol. Biotechnol. 2006, 16, 590–596. [Google Scholar]
- Brethauer, S.; Held, M.; Panke, S. Clavulanic acid decomposition is catalyzed by the compound itself and by its decomposition products. J. Pharm. Sci. 2008, 97, 3451–3455. [Google Scholar] [CrossRef]
- Roubos, J.A.; Krabben, P.; De Laat, W.; Heijnen, J.J. Clavulanic acid degradation in Streptomyces clavuligerus fed-batch cultivations. Biotechnol. Prog. 2002, 18, 451–457. [Google Scholar] [CrossRef]
- Nickolai, D.J.; Lammel, C.J.; Byford, B.A.; Morris, J.H.; Kaplan, E.B.; Hadley, W.K.; Brooks, G.F. Effects of storage temperature and pH on the stability of eleven b-lactam antibiotics in MIC trays. J. Clin. Microbiol. 1985, 21, 366–370. [Google Scholar] [PubMed]
- Okerman, L.; Van Hende, J.; De Zutter, L. Stability of frozen stock solutions of beta-lactam antibiotics, cephalosporins, tetracyclines and quinolones used in antibiotic residue screening and antibiotic susceptibility testing. Anal. Chim. Acta 2007, 586, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Vahdat, L.; Sunderland, V.B. Kinetics of amoxicillin and clavulanate degradation alone and in combination in aqueous solution under frozen conditions. Int. J. Pharm. 2007, 342, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Vetráková, L. Study of Processes Associated with Freezing of Aqueous Solutions. Ph.D. Thesis, Masaryk University, Brno, Czech Republic, 2017. [Google Scholar]
- Vahdat, L.; Sunderland, B. The influence of potassium clavulanate on the rate of amoxicillin sodium degradation in phosphate and acetate buffers in the liquid state. Drug Dev. Ind. Pharm. 2009, 35, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Jerzsele, Á.; Nagy, G. The stability of amoxicillin trihydrate and potassium clavulanate combination in aqueous solutions. Acta Vet. Hung. 2009, 57, 485–493. [Google Scholar] [CrossRef]
- Nur, A.O.; Hassan, A.A.; Gadkariem, E.A.; Osman, Z. Stability of co-amoxiclav reconstituted injectable solution. Eur. J. Pharm. Med. Res. 2015, 2, 109–123. [Google Scholar]
- Haginaka, J.; Yasuda, H.; Uno, T.; Nakagawa, T. Degradation of clavulanic acid in aqueous alkaline solution: isolation and structural investigation of degradation products. Chem. Pharm. Bull. 1985, 33, 218–224. [Google Scholar] [CrossRef]
- Finn, M.J.; Harris, M.A.; Hunt, E. Studies on the hydrolysis of clavulanic. J. Chem. Soc. Perkin Trans. 1984, 10, 1345–1349. [Google Scholar] [CrossRef]
- Zhong, C.; Cao, G.; Jin, X.; Wang, F. Studies on the formation and forming mechanism of the related substance E in potassium clavulanate production by HPLC-MS/MS. Braz. J. Pharm. Sci. 2014, 50, 391–399. [Google Scholar] [CrossRef]
- Soroka, D.; De La Sierra-Gallay, I.L.; Dubée, V.; Triboulet, S.; Van Tilbeurgh, H.; Compain, F.; Ballell, L.; Barros, D.; Mainardi, J.L.; Hugonnet, J.E.; et al. Hydrolysis of clavulanate by Mycobacterium tuberculosis β-lactamase BlaC harboring a canonical SDN motif. Antimicrob. Agents Chemother. 2015, 59, AAC-00598. [Google Scholar] [CrossRef] [PubMed]
- Peace, N.; Olubukola, O.; Moshood, A. Stability of reconstituted amoxicillin clavulanate potassium under simulated in-home storage conditions. J. Appl. Pharm. Sci. 2012, 2, 28–31. [Google Scholar]
- Gour-Salin, B.J. Hydrolysis rates of some acetylimidazole derivatives. Can. J. Chem. 1983, 61, 2059–2061. [Google Scholar] [CrossRef]
- Joule, J.A.; Mills, K. 1,3-Azoles: Imidazoles, thiazoles and oxazoles: Reactions and synthesis. In Heterocyclic Chemistry; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; pp. 461–467. ISBN 978-1-4051-3300-5. [Google Scholar]
- Ramirez-malule, H.; Junne, S.; Cruz-bournazou, M.N.; Neubauer, P. Streptomyces clavuligerus shows a strong association between TCA cycle intermediate accumulation and clavulanic acid biosynthesis. Appl. Microbiol. Biotechnol. 2018, 102, 4009–4023. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Malule, H.; Junne, S.; López, C.; Zapata, J.; Sáez, A.; Neubauer, P.; Rios-Estepa, R. An improved HPLC-DAD method for clavulanic acid quantification in fermentation broths of Streptomyces clavuligerus. J. Pharm. Biomed. Anal. 2016, 120, 241–247. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature | t < 5.5 h | t > 5.5 h | ||
|---|---|---|---|---|
| kobs,1 (h−1) | r2 | kobs,2 (h−1) | r2 | |
| −80 °C | 0.0009 | 0.969 | 0.0005 | 0.924 |
| −20 °C | 0.0135 | 0.991 | 0.0034 | 0.992 |
| 4 °C | 0.0241 | 0.996 | 0.0035 | 0.998 |
| 25 °C | 0.0320 | 0.998 | 0.0047 | 0.997 |
| Temperature | Equilibrium Constant Keq | Irreversible Rate Constant k2 (h−1) |
|---|---|---|
| −80 °C | 0.018 | 0.0009 |
| −20 °C | 0.162 | 0.0135 |
| 4 °C | 0.210 | 0.0241 |
| 25 °C | 0.280 | 0.0320 |
| Concentration (mg/L) | Temperature (°C) | NRMSE (%) | Shelf Life (t90) |
|---|---|---|---|
| 126.7 | −80 | 1.87 | 5.1 months |
| 126.7 | −20 | 3.22 | 9.0 h |
| 126.7 | 4 | 2.18 | 6.5 h |
| 126.7 | 25 | 3.78 | 4.5 h |
| 65.5 | −80 | 1.95 | 9.8 months |
| 65.5 | −20 | 4.98 | 9.7 h |
| 65.5 | 4 | 4.93 | 6.6 h |
| 65.5 | 25 | 4.45 | 4.6 h |
| 25.3 | −80 | 1.92 | 2.1 years |
| 25.3 | −20 | 3.36 | 9.9 h |
| 25.3 | 4 | 3.27 | 6.7 h |
| 25.3 | 25 | 3.40 | 4.7 h |
| 16.3 | −80 | 3.89 | 3.1 years |
| 16.3 | −20 | 1.98 | 10 h |
| 16.3 | 4 | 2.53 | 6.8 h |
| 16.3 | 25 | 2.42 | 4.8 h |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Ríos, D.; Ramírez-Malule, H.; Neubauer, P.; Junne, S.; Ríos-Estepa, R. Degradation Kinetics of Clavulanic Acid in Fermentation Broths at Low Temperatures. Antibiotics 2019, 8, 6. https://doi.org/10.3390/antibiotics8010006
Gómez-Ríos D, Ramírez-Malule H, Neubauer P, Junne S, Ríos-Estepa R. Degradation Kinetics of Clavulanic Acid in Fermentation Broths at Low Temperatures. Antibiotics. 2019; 8(1):6. https://doi.org/10.3390/antibiotics8010006
Chicago/Turabian StyleGómez-Ríos, David, Howard Ramírez-Malule, Peter Neubauer, Stefan Junne, and Rigoberto Ríos-Estepa. 2019. "Degradation Kinetics of Clavulanic Acid in Fermentation Broths at Low Temperatures" Antibiotics 8, no. 1: 6. https://doi.org/10.3390/antibiotics8010006
APA StyleGómez-Ríos, D., Ramírez-Malule, H., Neubauer, P., Junne, S., & Ríos-Estepa, R. (2019). Degradation Kinetics of Clavulanic Acid in Fermentation Broths at Low Temperatures. Antibiotics, 8(1), 6. https://doi.org/10.3390/antibiotics8010006