Protein Expression Modifications in Phage-Resistant Mutants of Aeromonas salmonicida after AS-A Phage Treatment

 ,
, 

Abstract
1. Introduction
2. Results
2.1. Detection of Host Sensitivity to Phages after Phage Contact
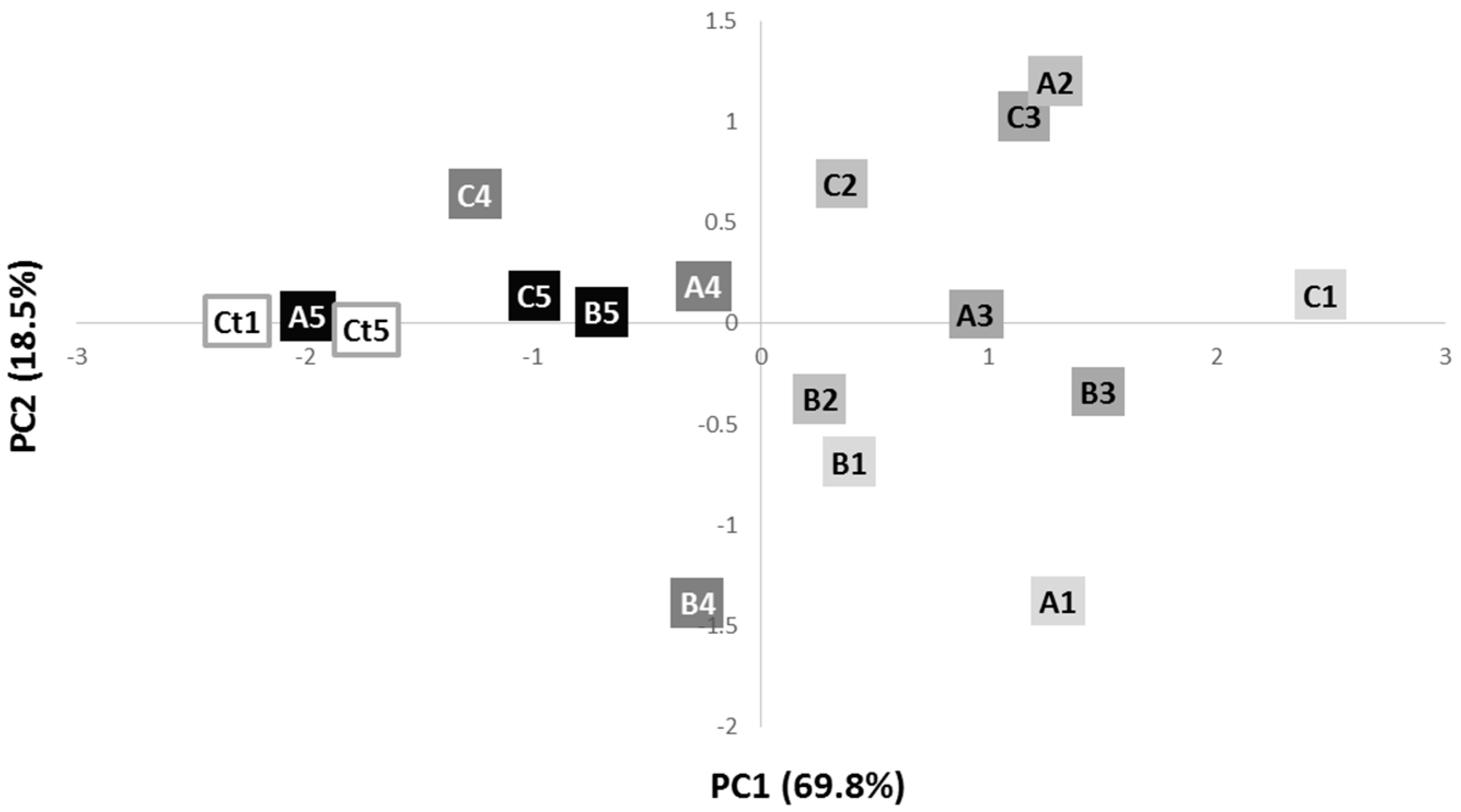
2.2. Infrared Spectroscopy of Whole Cells
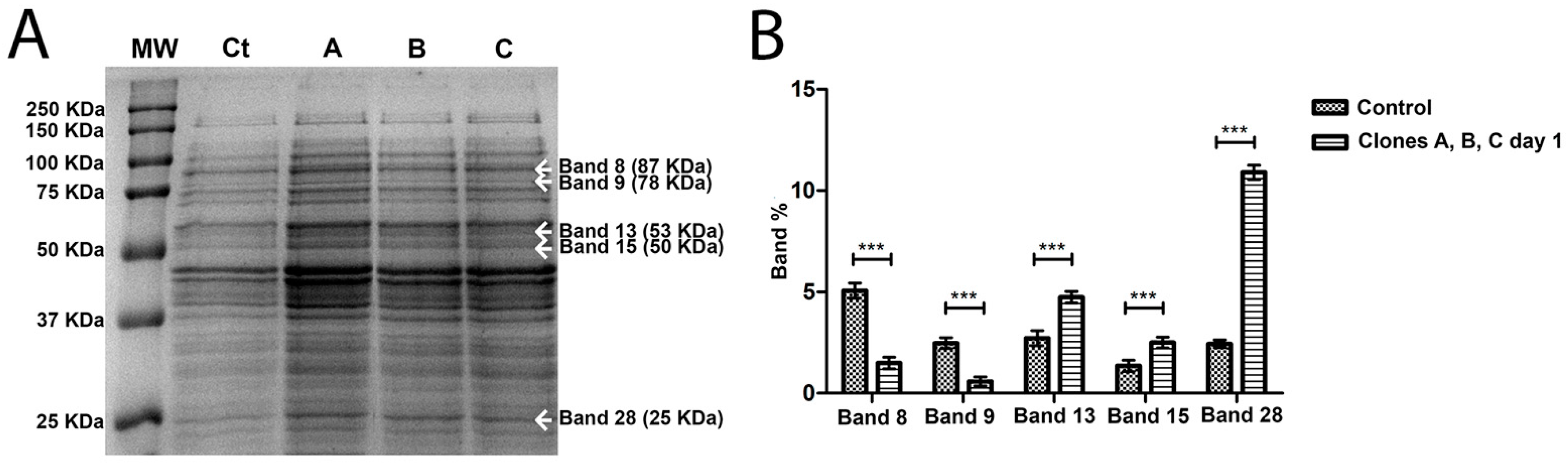
2.3. Differential Expression of the Proteins of the Phage-Resistant Clones (First Streak-Plating and Fifth Streak-Plating)
3. Discussion
4. Materials and Methods
4.1. Bacteria and Phage
4.2. Isolation of A. salmonicida Phage-Resistant Mutants
4.3. Detection of Bacteria Sensitivity to the Phage after One Cycle of Phage Contact
4.4. Efficiency of Plating (EOP)
4.5. Phage Adsorption
4.6. Infrared Spectroscopy
4.7. Extraction and Quantification of Intracellular Proteins from Phage-Sensitive and Phage-Resistant Bacteria
4.8. Protein Separation by 1-D Electrophoresis
4.9. Presumptive Identification of the Proteins in Differentially Expressed Bands
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture; FAO: Rome, Italy, 2014. [Google Scholar]
- Iwama, G.K.; Pickering, A.D.; Sumpter, J.P.; Schreck, C.B. Fish Stress and Health in Aquaculture; Cambridge University Press: Cambridge, UK, 2011; Volume 62, p. 279. [Google Scholar]
- Furushita, M.; Shiba, T.; Maeda, T.; Yahata, M.; Kaneoka, A.; Takahashi, Y.; Torii, K.; Hasegawa, T.; Ohta, M. Similarity of Tetracycline Resistance Genes Isolated from Fish Farm Bacteria to Those from Clinical Isolates. Appl. Environ. Microbiol. 2003, 69, 5336–5342. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Use of Antimicrobials Outside Medicine and Resultant Antimicrobial Resistance in Humans; WHO: Geneva, Switzerland, 2002; Volume 268, p. 2. [Google Scholar]
- Duckworth, D.H.; Gulig, P.A. Bacteriophages. BioDrugs 2002, 16, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Vadstein, O. The use of immunostimulation in marine larviculture: Possibilities and challenges. Aquaculture 1997, 155, 401–417. [Google Scholar] [CrossRef]
- Wiklund, T.; Dalsgaard, I. Occurrence and significance of atypical Aeromonas salmonicida in non-salmonid and salmonid fish species: A review. Dis. Aquat. Organ. 1998, 32, 49–69. [Google Scholar] [CrossRef] [PubMed]
- Uhland, F.C.; Martineau, D.; Mikaelian, I.; Canada, S.-L.V. Maladies des Poissons D’eau Douce du Québec: Guide de Diagnostic; University Press of Montreal: Montreal, QC, Canada, 2000; ISBN 2760617785. [Google Scholar]
- Boyd, J.; Williams, J.; Curtis, B.; Kozera, C.; Singh, R.; Reith, M. Three small, cryptic plasmids from Aeromonas salmonicida subsp. salmonicida A449. Plasmid 2003, 50, 131–144. [Google Scholar] [CrossRef]
- Burr, S.E.; Pugovkin, D.; Wahli, T.; Segner, H.; Frey, J. Attenuated virulence of an Aeromonas salmonicida subsp. salmonicida type III secretion mutant in a rainbow trout model. Microbiology 2005, 151, 2111–2118. [Google Scholar]
- Almeida, A.; Cunha, A.; Gomes, N.C.M.; Alves, E.; Costa, L.; Faustino, M.A.F. Phage therapy and photodynamic therapy: Low environmental impact approaches to inactivate microorganisms in fish farming plants. Mar. Drugs 2009, 7, 268–313. [Google Scholar] [CrossRef] [PubMed]
- Vieira, A.; Silva, Y.J.; Cunha, A.; Gomes, N.C.M.; Ackermann, H.-W.W.; Almeida, A.; Cunha, A.; Gomes, N.C.M.; Ackermann, H.-W.W.; Almeida, A. Phage therapy to control multidrug-resistant Pseudomonas aeruginosa skin infections: In vitro and ex vivo experiments. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 3241–3249. [Google Scholar] [CrossRef] [PubMed]
- Gill, J.; Hyman, P. Phage Choice, Isolation, and Preparation for Phage Therapy. Curr. Pharm. Biotechnol. 2010, 11, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.J.; Costa, L.; Pereira, C.; Mateus, C.; Cunha, A.; Calado, R.; Gomes, N.C.M.; Pardo, M.A.; Hernandez, I.; Almeida, A. Phage Therapy as an Approach to Prevent Vibrio anguillarum Infections in Fish Larvae Production. PLoS ONE 2014, 9, e114197. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Moreirinha, C.; Lewickab, M.; Almeida, P.; Clemente, C.; Delgadillo, I.; Romalde, J.L.; Nunes, M.L.; Lewicka, M.; Almeida, P.; et al. Bacteriophages with potential to inactivate Salmonella Typhimurium: Use of single phage suspensions and phage cocktails. Virus Res. 2016, 220, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Moreirinha, C.; Lewicka, M.; Almeida, P.; Clemente, C.; Romalde, J.L.; Nunes, M.; Almeida, A. Characterization and in vitro evaluation of new bacteriophages for the biocontrol of Escherichia coli. Virus Res. 2017, 227, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Heller, K.J. Molecular interaction between bacteriophage and the gram-negative cell envelope. Arch. Microbiol. 1992, 158, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Bull, J.J.; Vegge, C.S.; Schmerer, M.; Chaudhry, W.N.; Levin, B.R. Phenotypic Resistance and the Dynamics of Bacterial Escape from Phage Control. PLoS ONE 2014, 9, e94690. [Google Scholar] [CrossRef] [PubMed]
- Laanto, E.; Bamford, J.J.K.H.; Laakso, J.; Sundberg, L.L.-R. Phage-driven loss of virulence in a fish pathogenic bacterium. PLoS ONE 2012, 7, e53157. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.L.; Moreirinha, C.; Lopes, D.; Esteves, A.C.; Henriques, I.; Almeida, A.; Domingues, M.R.M.; Delgadillo, I.; Correia, A.; Cunha, A.; et al. Effects of UV Radiation on the Lipids and Proteins of Bacteria Studied by Mid-Infrared Spectroscopy. Environ. Sci. Technol. 2013, 47, 6306–6315. [Google Scholar] [CrossRef] [PubMed]
- Taillandier, E.; Liquier, J. DNA Structures Part A: Synthesis and Physical Analysis of DNA; Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1992; Volume 211, ISBN 9780121821128. [Google Scholar]
- Helm, D.; Naumann, D. Identification of some bacterial cell components by FT-IR spectroscopy. FEMS Microbiol. Lett. 1995, 126, 75–79. [Google Scholar] [CrossRef]
- Moreirinha, C.; Nunes, A.; Barros, A.A.; Almeida, A.; Delgadillo, I. Evaluation of the potential of Mid-infrared spectroscopy to assess the microbiological quality of ham. J. Food Saf. 2015, 35, 270–275. [Google Scholar] [CrossRef]
- Blanco, M.; Villarroya, I. NIR spectroscopy: A rapid-response analytical tool. TrAC Trends Anal. Chem. 2002, 21, 240–250. [Google Scholar] [CrossRef]
- Brereton, R. Chemometrics: Data Analysis for the Laboratoty and Chemical Plant; Wiley: London, UK, 2003. [Google Scholar]
- Barth, A. Infrared spectroscopy of proteins. Biochim. Biophys. Acta 2007, 1767, 1073–1101. [Google Scholar] [CrossRef] [PubMed]
- Alves, E.; Moreirinha, C.; Faustino, M.A.; Cunha, Â.; Delgadillo, I.; Neves, M.G.; Almeida, A. Overall biochemical changes in bacteria photosensitized with cationic porphyrins monitored by infrared spectroscopy. Future Med. Chem. 2016, 8, 613–628. [Google Scholar] [CrossRef] [PubMed]
- Pudziuvyte, B.; Bakiene, E.; Bonnett, R.; Shatunov, P.A.; Magaraggia, M.; Jori, G. Alterations of Escherichia coli envelope as a consequence of photosensitization with tetrakis(N-ethylpyridinium-4-yl)porphyrin tetratosylate. Photochem. Photobiol. Sci. 2011, 10, 1046. [Google Scholar] [CrossRef] [PubMed]
- Naumann, D. Infrared and NIR Raman Spectroscopy in Medical Microbiology. Proc. SPIE 1998, 3257. [Google Scholar] [CrossRef]
- Dovbeshko, G.I.; Gridina, N.Y.; Kruglova, E.B.; Pashchuk, O.P. FTIR spectroscopy studies of nucleic acid damage. In Talanta; Elsevier: Amsterdam, The Netherlands, 2000; Volume 53, pp. 233–246. [Google Scholar]
- Gasper, R.; Dewelle, J.; Kiss, R.; Mijatovic, T.; Goormaghtigh, E. IR spectroscopy as a new tool for evidencing antitumor drug signatures. Biochim. Biophys. Acta Biomembr. 2009, 1788, 1263–1270. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.C. Infrared Spectral Interpretation: A Systematic Approach; CRC Press: Boca Raton, FL, USA, 1999; ISBN 9780849324635. [Google Scholar]
- Huffman, S.W.; Lukasiewicz, K.; Geldart, S.; Elliott, S.; Sperry, J.F.; Brown, C.W. Analysis of Microbial Components Using LC-IR. Anal. Chem. 2003, 75, 4606–4611. [Google Scholar] [CrossRef] [PubMed]
- Salzer, R.; Siesler, H.W. Infrared and Raman Spectroscopic Imaging; Wiley-VCH: Weinheim, Germany, 2014; ISBN 3527336524. [Google Scholar]
- Stuart, B. Infrared Spectroscopy: Fundamentals and Applications, 2nd ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2004. [Google Scholar]
- Kudva, I.T.; Jelacic, S.; Tarr, P.I.; Hovde, C.J.; Youderian, P. Biocontrol of Escherichia coli O157 with Biocontrol of Escherichia coli O157 with O157-Specific Bacteriophages. Appl. Environ. Microbiol. 1999, 65, 3767–3773. [Google Scholar] [PubMed]
- Tomat, D.; Mercanti, D.; Balague, C.; Quiberoni, A. Phage biocontrol of enteropathogenic and shiga toxin-producing escherichia coli during milk fermentation. Lett. Appl. Microbiol. 2013, 57, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Nakai, T. Bacteriophage control of Pseudomonas plecoglossicida infection in ayu. Dis. Aquat. Organ. 2003, 53, 33–39. [Google Scholar] [CrossRef] [PubMed]
- O’Flynn, G.; Coffey, A.; Fitzgerald, G.; Ross, R. The newly isolated lytic bacteriophages st104a and st104b are highly virulent against Salmonella enterica. J. Appl. Microbiol. 2006, 101, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.J.; Moreirinha, C.; Pereira, C.; Costa, L.; Rocha, R.J.M.; Cunha, Â.; Gomes, N.C.M.; Calado, R.; Almeida, A. Biological control of Aeromonas salmonicida infection in juvenile Senegalese sole (Solea senegalensis) with Phage AS-A. Aquaculture 2016, 450, 225–233. [Google Scholar] [CrossRef]
- Filippov, A.A.; Sergueev, K.V.; He, Y.; Huang, X.-Z.; Gnade, B.T.; Mueller, A.J.; Fernandez-Prada, C.M.; Nikolich, M.P. Bacteriophage-resistant mutants in Yersinia pestis: Identification of phage receptors and attenuation for mice. PLoS ONE 2011, 6, e25486. [Google Scholar] [CrossRef] [PubMed]
- Levin, B.R.; Bull, J.J. Population and evolutionary dynamics of phage therapy. Nat. Rev. Microbiol. 2004, 2, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Koskella, B.; Brockhurst, M.A. Bacteria-phage coevolution as a driver of ecological and evolutionary processes in microbial communities. FEMS Microbiol. Rev. 2014, 38, 916–931. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, P.D.; Buckling, A.; Hall, A.R. Experimental evolution and bacterial resistance: (Co)evolutionary costs and trade-offs as opportunities in phage therapy research. Bacteriophage 2015, 5, e1050153. [Google Scholar] [CrossRef] [PubMed]
- Joly, N.; Schumacher, J.; Buck, M. Heterogeneous Nucleotide Occupancy Stimulates Functionality of Phage Shock Protein F, an AAA+ Transcriptional Activator. J. Biol. Chem. 2006, 281, 34997–35007. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, L.J.; Jones, S.E.; Jovanovic, G.; Gyaneshwar, P.; Rolfe, M.D.; Thompson, A.; Hinton, J.C.; Buck, M. Identification of a New Member of the Phage Shock Protein Response in Escherichia coli, the Phage Shock Protein G (PspG). J. Biol. Chem. 2004, 279, 55707–55714. [Google Scholar] [CrossRef] [PubMed]
- Darwin, A.J. The phage-shock-protein response. Mol. Microbiol. 2005, 57, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Brissette, J.L.; Russel, M.; Weiner, L.; Model, P. Phage shock protein, a stress protein of Escherichia coli. Proc. Natl. Acad. Sci. USA 1990, 87, 862–866. [Google Scholar] [CrossRef] [PubMed]
- Fröbel, J.; Rose, P.; Müller, M. Early Contacts between Substrate Proteins and TatA Translocase Component in Twin-arginine Translocation. J. Biol. Chem. 2011, 286, 43679–43689. [Google Scholar] [CrossRef] [PubMed]
- Bageshwar, U.K.; VerPlank, L.; Baker, D.; Dong, W.; Hamsanathan, S.; Whitaker, N.; Sacchettini, J.C.; Musser, S.M. High Throughput Screen for Escherichia coli Twin Arginine Translocation (Tat) Inhibitors. PLoS ONE 2016, 11, e0149659. [Google Scholar] [CrossRef] [PubMed]
- Friman, V.-P.; Hiltunen, T.; Jalasvuori, M.; Lindstedt, C.; Laanto, E.; Örmälä, A.-M.; Laakso, J.; Mappes, J.; Bamford, J.K.H. High Temperature and Bacteriophages Can Indirectly Select for Bacterial Pathogenicity in Environmental Reservoirs. PLoS ONE 2011, 6, e17651. [Google Scholar] [CrossRef] [PubMed]
- Domingues, S.; Harms, K.; Fricke, W.F.; Johnsen, P.J.; da Silva, G.J.; Nielsen, K.M. Natural Transformation Facilitates Transfer of Transposons, Integrons and Gene Cassettes between Bacterial Species. PLoS Pathog. 2012, 8, e1002837. [Google Scholar] [CrossRef] [PubMed]
- Schuster, C.F.; Mechler, L.; Nolle, N.; Krismer, B.; Zelder, M.-E.; Götz, F.; Bertram, R. The MazEF Toxin-Antitoxin System Alters the β-Lactam Susceptibility of Staphylococcus aureus. PLoS ONE 2015, 10, e0126118. [Google Scholar] [CrossRef] [PubMed]
- Magnuson, R.D. Hypothetical functions of toxin-antitoxin systems. J. Bacteriol. 2007, 189, 6089–6092. [Google Scholar] [CrossRef] [PubMed]
- Dy, R.L.; Przybilski, R.; Semeijn, K.; Salmond, G.P.C.; Fineran, P.C. A widespread bacteriophage abortive infection system functions through a Type IV toxin-antitoxin mechanism. Nucleic Acids Res. 2014, 42, 4590–4605. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, K.M.; Nilsson, A.S. Isolation of Phages for Phage Therapy: A Comparison of Spot Tests and Efficiency of Plating Analyses for Determination of Host Range and Efficacy. PLoS ONE 2015, 10, e0118557. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T. Lysis from without. Bacteriophage 2011, 1, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.H. Bacteriophages; John Wiley and Sons Inc.: New York, NY, USA, 1959. [Google Scholar]
- Stuer-Lauridsen, B.; Janzen, T.; Schnabl, J.; Johansen, E. Identification of the host determinant of two prolate-headed phages infecting lactococcus lactis. Virology 2003, 309, 10–17. [Google Scholar] [CrossRef]
- Laemli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Neuhoff, V.; Arold, N.; Taube, D.; Ehrhardt, W. Improved staining of proteins in polyacrylamide gels including isoelectric focusing gels with clear background at nanogram sensitivity using Coomassie Brilliant Blue G-250 and R-250. Electrophoresis 1988, 9, 255–262. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PC1 − (cm−1) | PC1 + (cm−1) | Assignment | Reference |
|---|---|---|---|
| 1695 | Amide I—proteins (β-sheet) | [27] | |
| 1650 | Amide I—proteins (α-helix) | [23,27] | |
| 1590, 1570, 1560 | Amide II—proteins | [27] | |
| 1510 | Amide II—proteins | [27] | |
| 1440 | CH3 bending—proteins (methyl groups) | [28] | |
| 1380 | COO−—acids and methyl groups from proteins/CO bonds or deformation of C-H or N-H bonds of proteins | [28,29] | |
| 1250 | Amide III—proteins/PO2−—phospholipids | [30,31] | |
| 1175 | C-O—proteins and glycomaterials | [32,33] | |
| 1150 | C-O carbohydrates | [33] | |
| 1070 | PO2−—nucleotides | [34] | |
| 1025 | Carbohydrates | [35] | |
| 980 | OCH3—polysaccharides | [36] |
| Band | MW (KDa) | Protein/Gene | Molecular Function |
|---|---|---|---|
| Band 8 | 87 | Phage transcriptional protein (ASA_3866) | Interacts selectively and non-covalently with the DNA with a specific nucleotide composition or with a specific sequence motif or type of DNA. |
| Band 9 | 78 | Phage shock protein B (pspB, ASA_2424) | Response of the bacteria to a variety of stimuli, including phage infection. It is involved in bacterial protection mechanisms. |
| Band 13 | 53 | Sec-independent protein translocase proteinTatA (tatA, ASA_3970) | Biological process: controlled liberation of proteins from a cell. |
| Band 15 | 50 | ASA_P5G151 | Unknown function. |
| Band 16 | 45 | Transposase (VO70_17345, VO70_21745) | Facilitates the transference of genetic material between organisms. |
| Band 18 | 40 | Toxin-antitoxin system, toxin component (VO68_18510, VO70_09250) | Plasmid maintenance, stress regulation and adaptation, growth control and programmed cellular death. |
| Band 28 | 25 | Q70WF0, Q70WF0_AERSA | Unknown function. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreirinha, C.; Osório, N.; Pereira, C.; Simões, S.; Delgadillo, I.; Almeida, A. Protein Expression Modifications in Phage-Resistant Mutants of Aeromonas salmonicida after AS-A Phage Treatment. Antibiotics 2018, 7, 21. https://doi.org/10.3390/antibiotics7010021
Moreirinha C, Osório N, Pereira C, Simões S, Delgadillo I, Almeida A. Protein Expression Modifications in Phage-Resistant Mutants of Aeromonas salmonicida after AS-A Phage Treatment. Antibiotics. 2018; 7(1):21. https://doi.org/10.3390/antibiotics7010021
Chicago/Turabian StyleMoreirinha, Catarina, Nádia Osório, Carla Pereira, Sara Simões, Ivonne Delgadillo, and Adelaide Almeida. 2018. "Protein Expression Modifications in Phage-Resistant Mutants of Aeromonas salmonicida after AS-A Phage Treatment" Antibiotics 7, no. 1: 21. https://doi.org/10.3390/antibiotics7010021
APA StyleMoreirinha, C., Osório, N., Pereira, C., Simões, S., Delgadillo, I., & Almeida, A. (2018). Protein Expression Modifications in Phage-Resistant Mutants of Aeromonas salmonicida after AS-A Phage Treatment. Antibiotics, 7(1), 21. https://doi.org/10.3390/antibiotics7010021