
Can Clays in Livestock Feed Promote Antibiotic Resistance and Virulence in Pathogenic Bacteria?

 and
and
Abstract
:1. Introduction

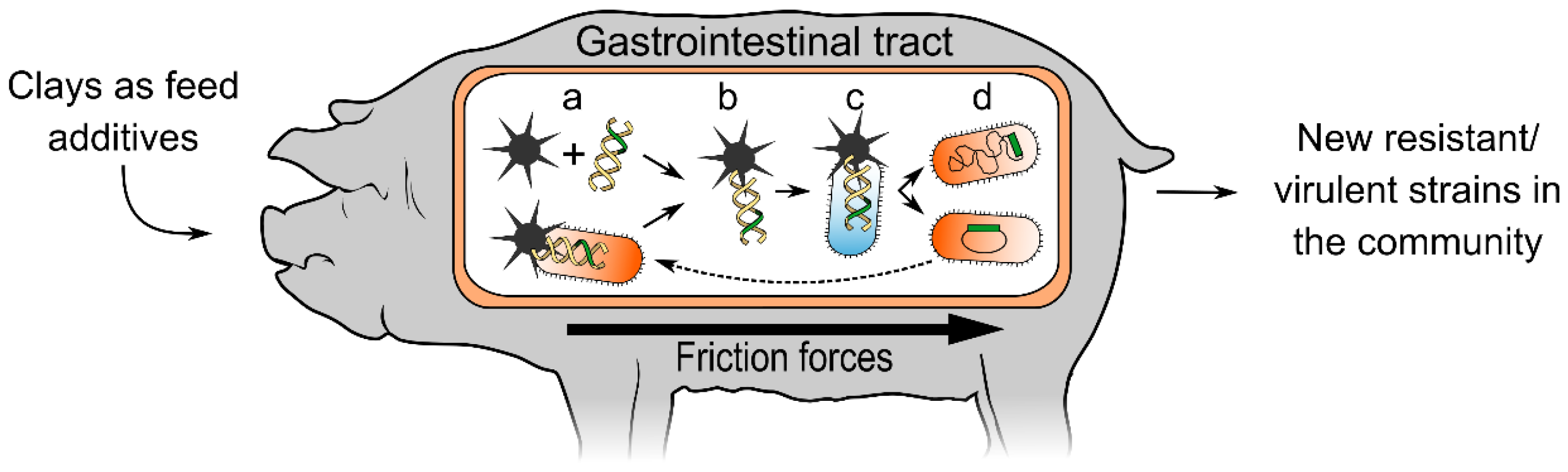
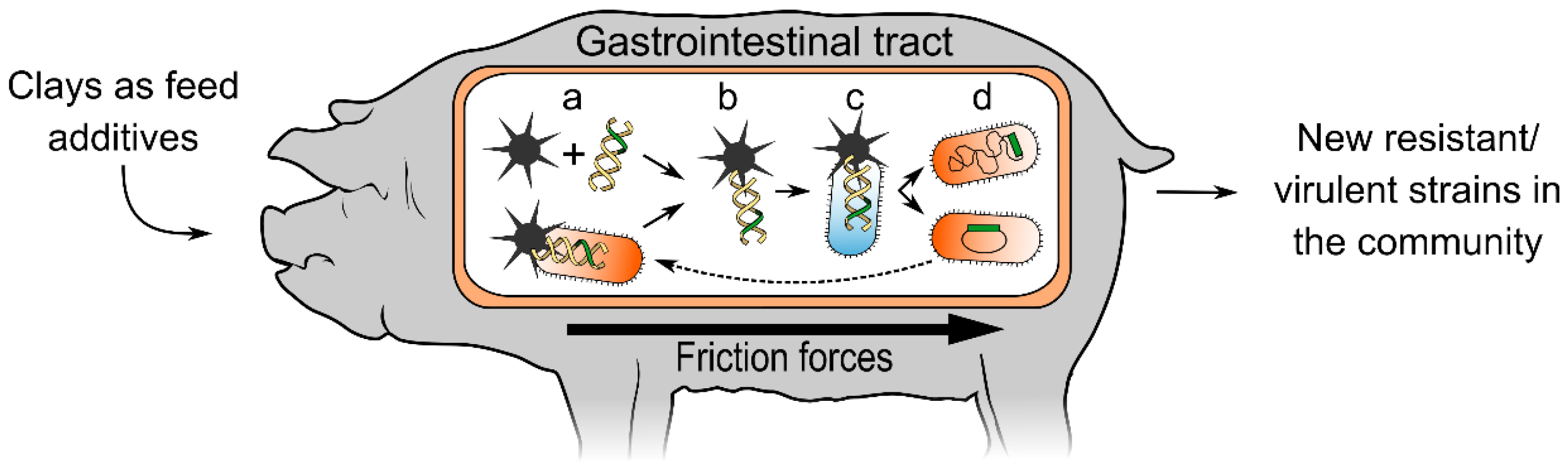{kind=link}
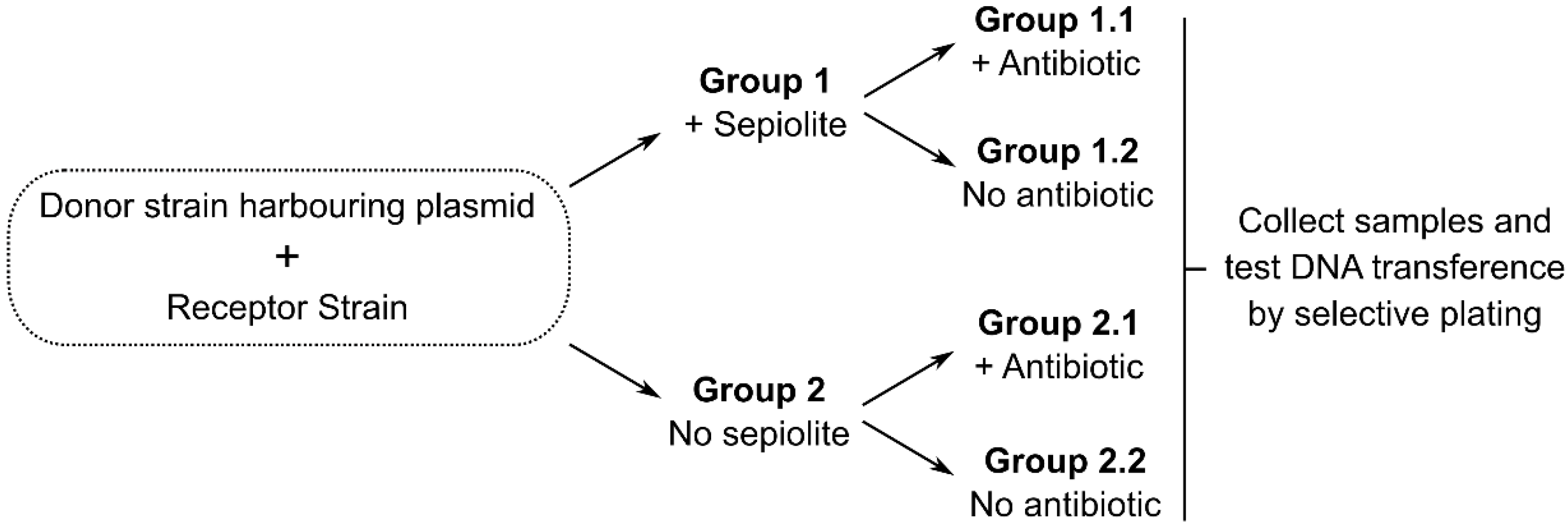
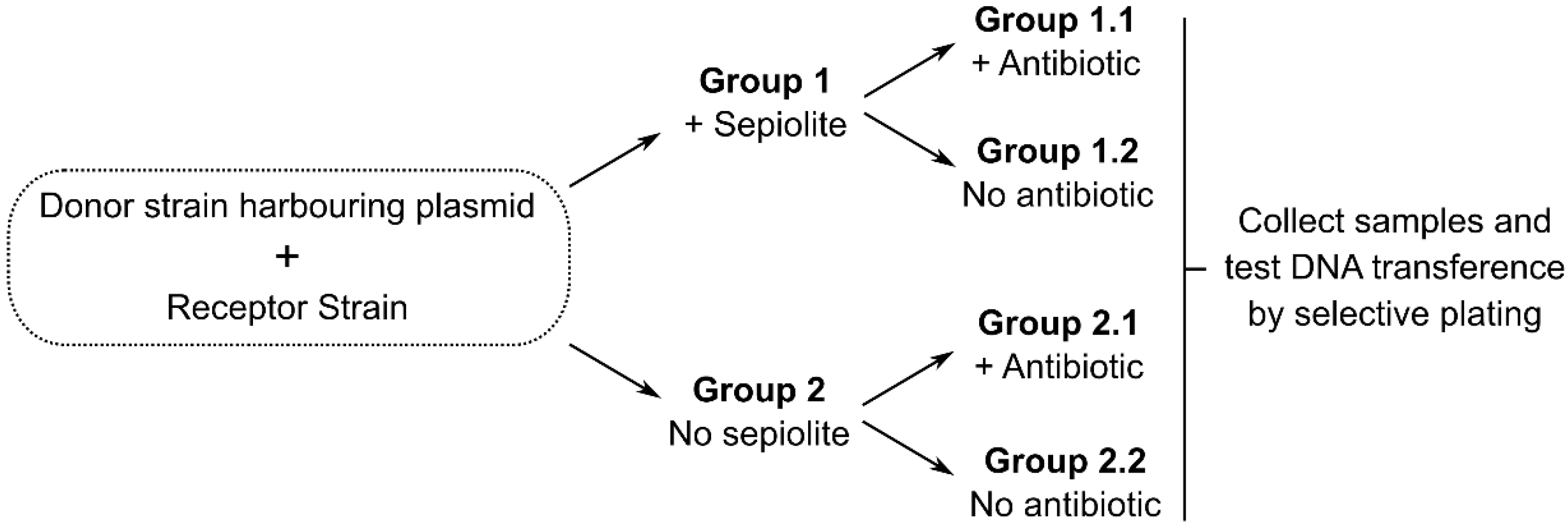{kind=link}
| Bacterial Species | References |
|---|---|
| Pseudomonas aeruginosa | [20] |
| Salmonella enterica Serovar Typhimurium | [20] |
| Mycobacterium smegmatis | [20] |
| Pseudomonas putida | [21] |
| Escherichia coli (several strains) | [21,22,23] |
| Yersinia enterocolitica | [23] |
| Acinetobacter baumannii | [23] |
| Bacillus subtilis | [16] |
| Pseudomonas elodea | [16] |
2. The Hypothesis
| Average Pressure of Digestive Tract (kPa) | Species | Digestive Tract Section | References |
|---|---|---|---|
| 2.7 | humans | jejunum | [28] |
| 2.7 | humans | pylorus | [29] |
| 10.7 | humans | oesophagus | [30] |
| 37.3 | geese | gizzard | [31] |
| 3.1 | turkeys | muscular stomach | [32] |
| 0.5–39.9 | chickens | gizzard | [33] |
| 3.9 | chickens | muscular stomach | [32] |
| 4.7 | pigs | stomach | [34] |
3. Testing the Hypothesis
4. Conclusions
Acknowledgments
Authors Contributions
Conflicts of Interest
References
- Blázquez, J.; Couce, A.; Rodríguez-Beltrán, J.; Rodríguez-Rojas, A. Antimicrobials as promoters of genetic variation. Curr. Opin. Microbiol. 2012, 15, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rojas, A.; Rodríguez-Beltrán, J.; Couce, A.; Blázquez, J. Antibiotics and antibiotic resistance: A bitter fight against evolution. Int. J. Med. Microbiol. 2013, 303, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Beaber, J.W.; Hochhut, B.; Waldor, M.K. SOS response promotes horizontal dissemination of antibiotic resistance genes. Nature 2004, 427, 72–74. [Google Scholar] [CrossRef] [PubMed]
- Couce, A.; Blazquez, J. Side effects of antibiotics on genetic variability. FEMS Microbiol. Rev. 2009, 33, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Ferber, D. Antibiotic resistance. WHO advises kicking the livestock antibiotic habit. Science 2003, 301. [Google Scholar] [CrossRef] [PubMed]
- Alcorn, T. Antibiotic use in livestock production in the USA. Lancet Infect. Dis. 2012, 12, 273–274. [Google Scholar] [CrossRef]
- Rizzi, A.; Raddadi, N.; Sorlini, C.; Nordgrd, L.; Nielsen, K.M.; Daffonchio, D. The stability and degradation of dietary DNA in the gastrointestinal tract of mammals: Implications for horizontal gene transfer and the biosafety of GMOs. Crit. Rev. Food Sci. Nutr. 2012, 52, 142–161. [Google Scholar] [CrossRef] [PubMed]
- Authority, E.F.S. Consolidated Presentation of the Joint Scientific Opinion of the GMO and BIOHAZ Panels. EFSA J. 2009, 1108, 1–8. [Google Scholar]
- Ouhida, I.; Pérez, J.F.; Piedrafita, J.; Gasa, J. The effects of sepiolite in broiler chicken diets of high, medium and low viscosity. Productive performance and nutritive value. Anim. Feed Sci. Technol. 2000, 85, 183–194. [Google Scholar] [CrossRef]
- Parisini, P.; Martelli, G.; Sardi, L.; Escribano, F. Protein and energy retention in pigs fed diets containing sepiolite. Anim. Feed Sci. Technol. 1999, 79, 155–162. [Google Scholar] [CrossRef]
- Carretero, M.I. Clay minerals and their beneficial effects upon human health. A review. Appl. Clay Sci. 2002, 21, 155–163. [Google Scholar] [CrossRef]
- Porubcan, L.S.; Serna, C.J.; White, J.L.; Hem, S.L. Mechanism of adsorption of clindamycin and tetracycline by montmorillonite. J. Pharm. Sci. 1978, 67, 1081–1087. [Google Scholar] [CrossRef] [PubMed]
- Otto, C.C.; Haydel, S.E. Exchangeable ions are responsible for the in vitro antibacterial properties of natural clay mixtures. PLoS ONE 2013, 8, e64068. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.B.; Metge, D.W.; Eberl, D.D.; Harvey, R.W.; Turner, A.G.; Prapaipong, P.; Poret-Peterson, A.T. What makes a natural clay antibacterial? Environ. Sci. Technol. 2011, 45, 3768–3773. [Google Scholar] [CrossRef] [PubMed]
- Tong, G.; Yulong, M.; Peng, G.; Zirong, X. Antibacterial effects of the Cu(II)-exchanged montmorillonite on Escherichia coli K88 and Salmonella choleraesuis. Vet. Microbiol. 2005, 105, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, N.; Fujiura, N. Earthquakes promote bacterial genetic exchange in serpentinite crevices. Astrobiology 2009, 9, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, N.; Sato, M. Plasmid uptake by bacteria: A comparison of methods and efficiencies. Appl. Microbiol. Biotechnol. 2009, 83, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Johnson, D.T. Adsorption of acicular particles at liquid-fluid interfaces and the influence of the line tension. Langmuir 2005, 21, 3838–3849. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, N.; Ide, K. Plasmid DNA is released from nanosized acicular material surface by low molecular weight oligonucleotides: Exogenous plasmid acquisition mechanism for penetration intermediates based on the Yoshida effect. Appl. Microbiol. Biotechnol. 2008, 80, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Beltrán, J.; Rodríguez-Rojas, A.; Yubero, E.; Blázquez, J. The animal food supplement sepiolite promotes a direct horizontal transfer of antibiotic resistance plasmids between bacterial species. Antimicrob. Agents Chemother. 2013, 57, 2651–2653. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Beltran, J.; Elabed, H.; Gaddour, K.; Blazquez, J.; Rodriguez-Rojas, A. Simple DNA transformation in Pseudomonas based on the Yoshida effect. J. Microbiol. Methods 2012, 89, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, N.; Nakajima-Kambe, T.; Matsuki, K.; Shigeno, T. Novel plasmid transformation method mediated by chrysotile, sliding friction, and elastic body exposure. Anal. Chem. Insights 2007, 2, 9–15. [Google Scholar] [PubMed]
- Wilharm, G.; Lepka, D.; Faber, F.; Hofmann, J.; Kerrinnes, T.; Skiebe, E. A simple and rapid method of bacterial transformation. J. Microbiol. Methods 2010, 80, 215–216. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Beltrán, J.; Rodríguez-Rojas, A.; Yubero, E.; Blázquez, J. The animal food supplement sepiolite promotes a direct horizontal transfer of antibiotic resistance plasmids between bacterial species. Antimicrob. Agents Chemother. 2013, 57, 2651–2653. [Google Scholar] [CrossRef] [PubMed]
- Guerin, E.; Cambray, G.; Sanchez-Alberola, N.; Campoy, S.; Erill, I.; da Re, S.; Gonzalez-Zorn, B.; Barbé, J.; Ploy, M.-C.; Mazel, D. The SOS response controls integron recombination. Science 2009. [Google Scholar] [CrossRef] [PubMed]
- López, E.; Elez, M.; Matic, I.; Blázquez, J. Antibiotic-mediated recombination: Ciprofloxacin stimulates SOS-independent recombination of divergent sequences in Escherichia coli. Mol. Microbiol. 2007, 64, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Khanna, M.; Stotzky, G. Transformation of Bacillus subtilis by DNA bound on montmorillonite and effect of DNase on the transforming ability of bound DNA. Appl. Environ. Microbiol. 1992, 58, 1930–1939. [Google Scholar] [PubMed]
- Scott, S.M.; Knowles, C.H.; Wang, D.; Yazaki, E.; Picon, L.; Wingate, D.L.; Lindberg, G. The nocturnal jejunal migrating motor complex: Defining normal ranges by study of 51 healthy adult volunteers and meta-analysis. Neurogastroenterol. Motil. 2006, 18, 927–935. [Google Scholar] [CrossRef] [PubMed]
- McShane, A.J.; O’Morain, C.; Lennon, J.R.; Coakley, J.B.; Alton, B.G. Atraumatic non-distorting pyloric sphincter pressure studies. Gut 1980, 21, 826–828. [Google Scholar] [CrossRef] [PubMed]
- Pope, C.E.; Horton, P.F. Intraluminal force transducer measurements of human oesophageal peristalsis. Gut 1972, 13, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.J. The comparative functional gizzard morphology of several species of birds. Aust. J. Zool. 1998. [Google Scholar] [CrossRef]
- Duke, G.E.; Dziuk, H.E.; Evanson, O.A. Gastric pressure and smooth muscle electrical potential changes in turkeys. Am. J. Physiol. 1972, 222, 167–173. [Google Scholar] [PubMed]
- Lentle, R.G.; Reynolds, G.; de Loubens, C.; Hulls, C.; Janssen, P.W.M.; Ravindran, V. Spatiotemporal mapping of the muscular activity of the gizzard of the chicken (Gallus domesticus). Poult. Sci. 2013, 92, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Desilets, D.J.; Mader, T.J.; Romanelli, J.R.; Earle, D.B. Gastric transmural pressure measurements in vivo: Implications for natural orifice transluminal endoscopic surgery (NOTES). Gastrointest. Endosc. 2010, 71, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Merchant, H.A.; McConnell, E.L.; Liu, F.; Ramaswamy, C.; Kulkarni, R.P.; Basit, A.W.; Murdan, S. Assessment of gastrointestinal pH, fluid and lymphoid tissue in the guinea pig, rabbit and pig, and implications for their use in drug development. Eur. J. Pharm. Sci. 2011, 42, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Mabelebele, M.; Alabi, O.J.; Ngambi, J.W.; Norris, D.; Ginindza, M.M. Comparison of gastrointestinal tracts and pH values of digestive organs of ross 308 broiler and indigenous venda chickens fed the same diet. Asian J. Anim. Vet. Adv. 2014, 9, 71–76. [Google Scholar] [CrossRef]
- Sleator, R.D.; Hill, C. Compatible solutes: The key to Listeria’s success as a versatile gastrointestinal pathogen? Gut Pathog. 2010. [Google Scholar] [CrossRef] [PubMed]
- Demanèche, S.; Jocteur-Monrozier, L.; Quiquampoix, H.; Simonet, P. Evaluation of biological and physical protection against nuclease degradation of clay-bound plasmid DNA. Appl. Environ. Microbiol. 2001, 67, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Huddleston, J.R. Horizontal gene transfer in the human gastrointestinal tract: Potential spread of antibiotic resistance genes. Infect. Drug Resist. 2014, 7, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Hausner, M.; Wuertz, S. High rates of conjugation in bacterial biofilms as determined by quantitative in situ analysis. Appl. Environ. Microbiol. 1999, 65, 3710–3713. [Google Scholar] [PubMed]
- Licht, T.R.; Christensen, B.B.; Krogfelt, K.A.; Molin, S. Plasmid transfer in the animal intestine and other dynamic bacterial populations: The role of community structure and environment. Microbiology 1999, 145, 2615–2622. [Google Scholar] [PubMed]
- Faure, S.; Perrin-Guyomard, A.; Delmas, J.M.; Chatre, P.; Laurentie, M. Transfer of plasmid-mediated CTX-M-9 from Salmonella enterica serotype virchow to enterobacteriaceae in human flora-associated rats treated with cefixime. Antimicrob. Agents Chemother. 2009, 54, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, A.-K.; Bøhn, T.; Primicerio, R.; Shorten, P.R.; Soboleva, T.K.; Nielsen, K.M. Modeling suggests frequency estimates are not informative for predicting the long-term effect of horizontal gene transfer in bacteria. Environ. Biosafety Res. 2005, 4, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.M.; Townsend, J.P. Monitoring and modeling horizontal gene transfer. Nat. Biotechnol. 2004, 22, 1110–1114. [Google Scholar] [CrossRef] [PubMed]
- Mater, D.D.G. Evidence of vancomycin resistance gene transfer between enterococci of human origin in the gut of mice harbouring human microbiota. J. Antimicrob. Chemother. 2005, 56, 975–978. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Rojas, A.; Rodríguez-Beltrán, J.; Valverde, J.R.; Blázquez, J. Can Clays in Livestock Feed Promote Antibiotic Resistance and Virulence in Pathogenic Bacteria? Antibiotics 2015, 4, 299-308. https://doi.org/10.3390/antibiotics4030299
Rodríguez-Rojas A, Rodríguez-Beltrán J, Valverde JR, Blázquez J. Can Clays in Livestock Feed Promote Antibiotic Resistance and Virulence in Pathogenic Bacteria? Antibiotics. 2015; 4(3):299-308. https://doi.org/10.3390/antibiotics4030299
Chicago/Turabian StyleRodríguez-Rojas, Alexandro, Jerónimo Rodríguez-Beltrán, José Ramón Valverde, and Jesús Blázquez. 2015. "Can Clays in Livestock Feed Promote Antibiotic Resistance and Virulence in Pathogenic Bacteria?" Antibiotics 4, no. 3: 299-308. https://doi.org/10.3390/antibiotics4030299
APA StyleRodríguez-Rojas, A., Rodríguez-Beltrán, J., Valverde, J. R., & Blázquez, J. (2015). Can Clays in Livestock Feed Promote Antibiotic Resistance and Virulence in Pathogenic Bacteria? Antibiotics, 4(3), 299-308. https://doi.org/10.3390/antibiotics4030299