
Finding the Missing IMP Gene: Overcoming the Imipenemase IMP Gene Drop-Out in Automated Molecular Testing for Carbapenem-Resistant Bacteria Circulating in Latin America

 ,
,  ,
,  , , , , ,
, , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Identification and Antimicrobial Susceptibility Testing
2.3. Automated qPCR Analysis for Carbapenemase Gene Detection
2.4. Genomic DNA Extraction and Quantification
2.5. Redesign of Primers to Extend Detection of IMP Alleles
2.6. Tailored qPCR
2.7. Whole-Genome Sequencing, Assembly, and Annotation
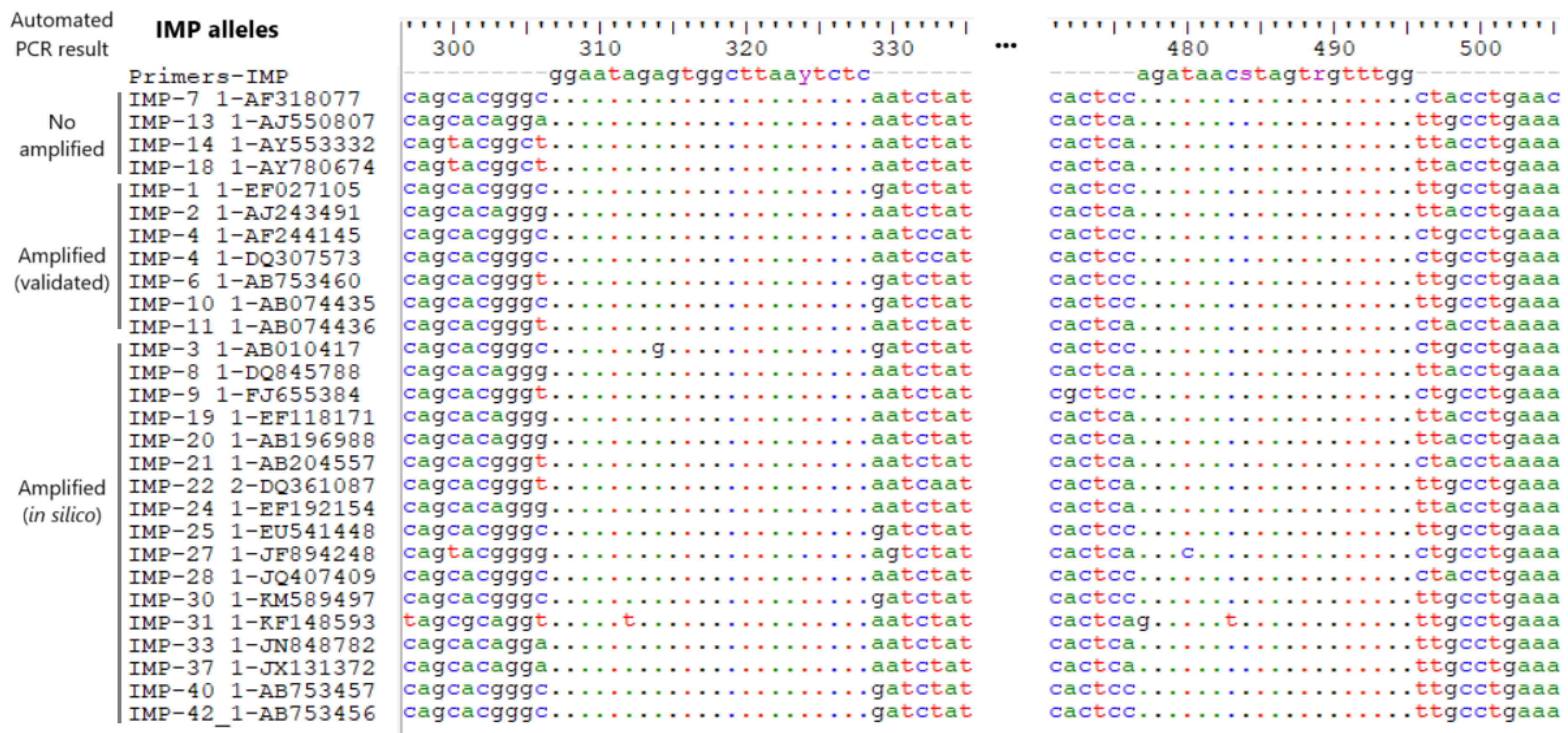
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| IMP | Imipenemase IMP |
| PCR | Polymerase Chain Reaction |
| qPCR | Real-time PCR |
References
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, G.; Midiri, A.; Gerace, E.; Biondo, C. Bacterial antibiotic resistance: The most critical pathogens. Pathogens 2021, 10, 1310. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for the Prevention and Control of Carbapenem-Resistant Enterobacteriaceae, Acinetobacter baumannii and Pseudomonas aeruginosa in Health Care Facilities; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- World Health Organization. WHO Bacterial Priority Pathogens List. 2024. Available online: https://iris.who.int/bitstream/handle/10665/376776/9789240093461-eng.pdf?sequence=1 (accessed on 6 June 2025).
- Mancuso, G.; De Gaetano, S.; Midiri, A.; Zummo, S.; Biondo, C. The Challenge of Overcoming Antibiotic Resistance in Carbapenem-Resistant Gram-Negative Bacteria: “Attack on Titan”. Microorganisms 2023, 11, 1912. [Google Scholar] [CrossRef]
- Logan, L.K.; Weinstein, R.A. The epidemiology of Carbapenem-resistant enterobacteriaceae: The impact and evolution of a global menace. J. Infect. Dis. 2017, 215, S28–S36. [Google Scholar] [CrossRef]
- Budia-Silva, M.; Kostyanev, T.; Ayala-Montaño, S.; Bravo-Ferrer Acosta, J.; Garcia-Castillo, M.; Cantón, R.; Goossens, H.; Rodriguez-Baño, J.; Grundmann, H.; Reuter, S. International and regional spread of carbapenem-resistant Klebsiella pneumoniae in Europe. Nat. Commun. 2024, 15, 5092. [Google Scholar] [CrossRef]
- Baeza, L.L.; Pfennigwerth, N.; Greissl, C.; Göttig, S.; Saleh, A.; Stelzer, Y.; Gatermann, S.G.; Hamprecht, A. Comparison of five methods for detection of carbapenemases in Enterobacterales with proposal of a new algorithm. Clin. Microbiol. Infect. 2019, 25, 1286.e9–1286.e15. [Google Scholar] [CrossRef] [PubMed]
- Gomes Chagas, T.P.; Rangel, K.; De-Simone, S. Carbapenem-resistant Acinetobacter baumannii in Latin America. In Acinetobacter baumannii—The Rise of a Resistant Pathogen; IntechOpen: London, UK, 2024; ISBN 978-0-85466-370-5. [Google Scholar]
- Monteiro, J.; Widen, R.H.; Pignatari, A.C.C.; Kubasek, C.; Silbert, S. Rapid detection of carbapenemase genes by multiplex real-time PCR. J. Antimicrob. Chemother. 2012, 67, 906–909. [Google Scholar] [CrossRef]
- Hong, D.J.; Bae, I.K.; Jang, I.-H.; Jeong, S.H.; Kang, H.-K.; Lee, K. Epidemiology and characteristics of metallo-ß-lactamase-producing Pseudomonas aeruginosa. Infect. Chemother. 2015, 47, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Meletis, G.; Exindari, M.; Vavatsi, N.; Sofianou, D.; Diza, E. Mechanisms responsible for the emergence of carbapenem resistance in Pseudomonas aeruginosa. Hippokratia 2012, 16, 303–307. [Google Scholar]
- Molina-Mora, J.A.; Chinchilla-Montero, D.; García-Batán, R.; García, F. Genomic context of the two integrons of ST-111 Pseudomonas aeruginosa AG1: A VIM-2-carrying old-acquaintance and a novel IMP-18-carrying integron. Infect. Genet. Evol. 2021, 89, 104740. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, K.L.; Meunier, D.; Naas, T.; Volland, H.; Woodford, N. Evaluation of the NG-Test CARBA 5 multiplex immunochromatographic assay for the detection of KPC, OXA-48-like, NDM, VIM and IMP carbapenemases. J. Antimicrob. Chemother. 2018, 73, 3523–3526. [Google Scholar] [CrossRef]
- Traczewski, M.M.; Carretto, E.; Canton, R.; Moore, N.M.; Brovarone, F.; Nardini, P.; Visiello, R.; García-Castillo, M.; Ruiz-Garbajosa, P.; Tato, M. Multicenter evaluation of the xpert carba-R assay for detection of carbapenemase genes in gram-negative isolates. J. Clin. Microbiol. 2018, 56, e00272-18. [Google Scholar] [CrossRef] [PubMed]
- Walkty, A.; Adam, H.; Tran, V.; Alexander, D.C.; Boyd, D.; Bharat, A.; Karlowsky, J. Failure of a multiplex polymerase chain reaction assay to detect IMP-27 in a clinical isolate of Morganella morganii. Diagn. Microbiol. Infect. Dis. 2018, 92, 194–195. [Google Scholar] [CrossRef]
- de Mello Malta, F.; Amgarten, D.; Marra, A.R.; Petroni, R.C.; da Silva Nali, L.H.; Siqueira, R.A.; Neto, M.C.; Oler, S.C.; Pinho, J.R.R. Nucleocapsid single point-mutation associated with drop-out on RT-PCR assay for SARS-CoV-2 detection. BMC Infect. Dis. 2023, 23, 714. [Google Scholar] [CrossRef]
- Blais, J.; Lavoie, S.B.; Giroux, S.; Bussières, J.; Lindsay, C.; Dionne, J.; Laroche, M.; Giguère, Y.; Rousseau, F. Risk of Misdiagnosis Due to Allele Dropout and False-Positive PCR Artifacts in Molecular Diagnostics: Analysis of 30,769 Genotypes. J. Mol. Diagn. 2015, 17, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Almotiri, M.; Alzeyadi, A.; Antimicrobial, Z.A.; Irfan, M.; Almotiri, A.; Alzeyadi, Z.A. Antimicrobial Resistance and Its Drivers—A Review. Antibiot 2022, 11, 1362. [Google Scholar]
- Yang, L.; Ding, F.; Lin, Q.; Xie, J.; Fan, W.; Dai, F.; Cui, P.; Liu, W. A tool to automatically design multiplex PCR primer pairs for specific targets using diverse templates. Sci. Rep. 2023, 13, 16451. [Google Scholar] [CrossRef]
- Huang, J.; Lv, C.; Li, M.; Rahman, T.; Chang, Y.F.; Guo, X.; Song, Z.; Zhao, Y.; Li, Q.; Ni, P.; et al. Carbapenem-resistant Escherichia coli exhibit diverse spatiotemporal epidemiological characteristics across the globe. Commun. Biol. 2024, 7, 51. [Google Scholar] [CrossRef]
- García-Betancur, J.C.; Appel, T.M.; Esparza, G.; Gales, A.C.; Levy-Hara, G.; Cornistein, W.; Vega, S.; Nuñez, D.; Cuellar, L.; Bavestrello, L.; et al. Update on the epidemiology of carbapenemases in Latin America and the Caribbean. Expert Rev. Anti. Infect. Ther. 2021, 19, 197–213. [Google Scholar] [CrossRef]
- Molina-Mora, J.A.; García, F. Molecular Determinants of Antibiotic Resistance in the Costa Rican Pseudomonas aeruginosa AG1 by a Multi-omics Approach: A Review of 10 Years of Study. Phenomics 2021, 1, 3. [Google Scholar] [CrossRef]
- dos Santos, L.A.; Cayô, R.; Valiatti, T.B.; Gales, A.C.; de Araújo, L.F.B.; Rodrigues, F.M.; de Carvalho, T.S.; Vaz, M.A.B.; Campanharo, M. Biodiversity of carbapenem-resistant bacteria in clinical samples from the Southwest Amazon region (Rondônia/Brazil). Sci. Rep. 2024, 14, 9383. [Google Scholar] [CrossRef] [PubMed]
- Garza-Ramos, U.; Rodríguez-Medina, N.; Córdova-Fletes, C.; Rubio-Mendoza, D.; Alonso-Hernández, C.J.; Esaú López-Jácome, L.; Morfín-Otero, R.; Rodríguez-Noriega, E.; Rojas-Larios, F.; del Rosario Vázquez-Larios, M.; et al. Whole genome analysis of Gram-negative bacteria using the EPISEQ CS application and other bioinformatic platforms. J. Glob. Antimicrob. Resist. 2023, 33, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez-Prada, E.D.; Bustos, I.G.; Gamboa-Silva, E.; Josa, D.F.; Mendez, L.; Fuentes, Y.V.; Serrano-Mayorga, C.C.; Baron, O.; Ruiz-Cuartas, A.; Silva, E.; et al. Molecular characterization and descriptive analysis of carbapenemase-producing Gram-negative rod infections in Bogota, Colombia. Microbiol. Spectr. 2024, 12, e0171423. [Google Scholar] [CrossRef]
- Universidad de Costa Rica iPAT: Plataforma Genómica, Bioinformática y de Inteligencia Artificial para la Vigilancia de Patógenos. Available online: https://vinv.ucr.ac.cr/sigpro/web/projects/C4604 (accessed on 6 June 2025).
- Florensa, A.F.; Kaas, R.S.; Clausen, P.T.L.C.; Aytan-Aktug, D.; Aarestrup, F.M. ResFinder—An open online resource for identification of antimicrobial resistance genes in next-generation sequencing data and prediction of phenotypes from genotypes. Microb. Genom. 2022, 8, 000748. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Cepheid Xpert® Carba-R 2020. Available online: https://www.cepheid.com/es-ES/tests/hai-other-infectious-diseases/xpert-carba-r.html (accessed on 28 March 2025).
- Deshpande, L.M.; Vega, S.; Tinoco, J.C.; Castanheira, M. Endemicity of Pseudomonas aeruginosa producing IMP-18 and/or VIM-2 MBLs from the high-risk clone ST111 in Central America. JAC-Antimicrob. Resist. 2023, 5, dlad092. [Google Scholar] [CrossRef]
- Toval, F.; Guzmán-Marte, A.; Madriz, V.; Somogyi, T.; Rodríguez, C.; García, F. Predominance of carbapenem-resistant Pseudomonas aeruginosa isolates carrying blaIMP and blaVIM metallo-β-lactamases in a major hospital in Costa Rica. J. Med. Microbiol. 2015, 64, 37–43. [Google Scholar] [CrossRef]
- Mentasti, M.; Prime, K.; Sands, K.; Khan, S.; Wootton, M. Rapid detection of IMP, NDM, VIM, KPC and OXA-48-like carbapenemases from Enterobacteriales and Gram-negative non-fermenter bacteria by real-time PCR and melt-curve analysis. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 2029–2036. [Google Scholar] [CrossRef] [PubMed]
- Sheh, A.; Artim, S.C.; Burns, M.A.; Molina-Mora, J.A.; Lee, M.A.; Dzink-Fox, J.; Muthupalani, S.; Fox, J.G. Analysis of gut microbiome profiles in common marmosets (Callithrix jacchus) in health and intestinal disease. Sci. Rep. 2022, 12, 4430. [Google Scholar] [CrossRef]
- Molina-Mora, J.A.; Sibaja-Amador, M.; Rivera-Montero, L.; Chacón-Arguedas, D.; Guzmán, C.; García, F. Assessment of Mathematical Approaches for the Estimation and Comparison of Efficiency in qPCR Assays for a Prokaryotic Model. DNA 2024, 4, 189–200. [Google Scholar] [CrossRef]
- Andrews, S. FastQC A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 10 April 2018).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, C.-M.; Luo, R.; Sadakane, K.; Lam, T.-W. MEGAHIT: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Molina-Mora, J.A.; Garcia, F. The 3C criterion: Contiguity, Completeness and Correctness to assess de novo genome assemblies. BMC Bioinform. Bioinforma. Algorithms Appl. 2020, 21, 5. [Google Scholar]
- Molina-Mora, J.-A.; Campos-Sánchez, R.; Rodríguez, C.; Shi, L.; García, F. High quality 3C de novo assembly and annotation of a multidrug resistant ST-111 Pseudomonas aeruginosa genome: Benchmark of hybrid and non-hybrid assemblers. Sci. Rep. 2020, 10, 1392. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Molina-Mora, J.A.; Chinchilla, D.; Chavarría, M.; Ulloa, A.; Campos-Sanchez, R.; Mora-Rodríguez, R.A.; Shi, L.; García, F. Transcriptomic determinants of the response of ST-111 Pseudomonas aeruginosa AG1 to ciprofloxacin identified by a top-down systems biology approach. Sci. Rep. 2020, 10, 13717. [Google Scholar] [CrossRef] [PubMed]
- Molina-Mora, J.A.; Chinchilla-Montero, D.; Castro-Peña, C.; García, F. Two-dimensional gel electrophoresis (2D-GE) image analysis based on CellProfiler. Medicine 2020, 99, e23373. [Google Scholar] [CrossRef]
- Cason, C.; D’Accolti, M.; Soffritti, I.; Mazzacane, S.; Comar, M.; Caselli, E. Next-generation sequencing and PCR technologies in monitoring the hospital microbiome and its drug resistance. Front. Microbiol. 2022, 13, 969863. [Google Scholar] [CrossRef]
- Zurita, J.; Sevillano, G.; Solís, M.B.; Paz y Miño, A.; Alves, B.R.; Changuan, J.; González, P. Pseudomonas aeruginosa epidemic high-risk clones and their association with multidrug-resistant. J. Glob. Antimicrob. Resist. 2024, 38, 332–338. [Google Scholar] [CrossRef]
- Wolter, D.J.; Khalaf, N.; Robledo, I.E.; Vázquez, J.G.; Santé, I.M.; Aquino, E.E.; Goering, R.V.; Hanson, N.D. Surveillance of Carbapenem-Resistant Pseudomonas aeruginosa Isolates from Puerto Rican Medical Center Hospitals: Dissemination of KPC and IMP-18 β-Lactamases. Antimicrob. Agents Chemother. 2009, 53, 1660. [Google Scholar] [CrossRef]
- Borgianni, L.; Prandi, S.; Salden, L.; Santella, G.; Hanson, N.D.; Rossolini, G.M.; Docquier, J.D. Genetic context and biochemical characterization of the IMP-18 metallo-β-lactamase identified in a Pseudomonas aeruginosa isolate from the United States. Antimicrob. Agents Chemother. 2011, 55, 140–145. [Google Scholar] [CrossRef]
- Kazmierczak, K.M.; Karlowsky, J.A.; de Jonge, B.L.M.; Stone, G.G.; Sahm, D.F. Epidemiology of Carbapenem Resistance Determinants Identified in Meropenem-Nonsusceptible Enterobacterales Collected as Part of a Global Surveillance Program, 2012 to 2017. Antimicrob. Agents Chemother. 2021, 65, e02000-20. [Google Scholar] [CrossRef]
- Yoon, E.J.; Jeong, S.H. Mobile Carbapenemase Genes in Pseudomonas aeruginosa. Front. Microbiol. 2021, 12, 614058. [Google Scholar] [CrossRef]
- Duze, S.T.; Thomas, T.; Pelego, T.; Jallow, S.; Perovic, O.; Duse, A. Evaluation of Xpert Carba-R for detecting carbapenemase-producing organisms in South Africa. Afr. J. Lab. Med. 2023, 12, 1898. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Park, J.Y.; Kim, J.H.; Lee, Y.H.; Yang, H.Y.; Yoo, J.S. Outbreak of Imipenemase-1-Producing Carbapenem-Resistant Klebsiella pneumoniae in an Intensive Care Unit. Acute Crit. Care 2016, 32, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Koh, T.H.; Babini, G.S.; Woodford, N.; Sng, L.H.; Hall, L.M.C.; Livermore, D.M. Carbapenem-hydrolysing IMP-1 β-lactamase in Klebsiella pneumoniae from Singapore. Lancet 1999, 353, 2162. [Google Scholar] [CrossRef]
- Cheng, Z.; Bethel, C.R.; Thomas, P.W.; Shurina, B.A.; Alao, J.P.; Thomas, C.A.; Yang, K.; Marshall, S.H.; Zhang, H.; Sturgill, A.M.; et al. Carbapenem Use Is Driving the Evolution of Imipenemase 1 Variants. Antimicrob. Agents Chemother. 2021, 65, e01714-20. [Google Scholar] [CrossRef] [PubMed]
- Wink, P.L.; Almeida, E.K.; Crispim, M.N.; De Lima-Morales, D.; Zavascki, A.P.; Barth, A.L. First report of IMP-1 in a clinical isolate of Escherichia coli in Latin America. Infect. Control Hosp. Epidemiol. 2020, 41, 997–998. [Google Scholar] [CrossRef]
- Ramos, A.C.; Cayô, R.; Carvalhaes, C.G.; Jové, T.; Da Silva, G.P.; Sancho, F.M.P.; Chagas-Neto, T.; Medeiros, E.A.S.; Gales, A.C. Dissemination of multidrug-resistant proteus mirabilis clones carrying a novel integron-borne bla IMP-1 in a tertiary hospital. Antimicrob. Agents Chemother. 2018, 62, e01321-17. [Google Scholar] [CrossRef]
- Picão, R.C.; Andrade, S.S.; Nicoletti, A.G.; Campana, E.H.; Moraes, G.C.; Mendes, R.E.; Gales, A.C. Metallo-β-Lactamase Detection: Comparative Evaluation of Double-Disk Synergy versus Combined Disk Tests for IMP-, GIM-, SIM-, SPM-, or VIM-Producing Isolates. J. Clin. Microbiol. 2008, 46, 2028. [Google Scholar] [CrossRef]
- García-González, N.; Beamud, B.; Sevilla-Fortuny, J.; Sánchez-Hellín, V.; Vidal, I.; Rodríguez, J.C.; Fuster, B.; Tormo, N.; Salvador, C.; Gimeno, C.; et al. Genomic surveillance reveals different transmission patterns between third-generation cephalosporin and carbapenem resistance in Klebsiella pneumoniae in the Comunidad Valenciana (Spain), 2018–2020. Antimicrob. Resist. Infect. Control 2025, 14, 44. [Google Scholar] [CrossRef]
- McMillen, T.; Jani, K.; Robilotti, E.V.; Kamboj, M.; Babady, N.E. The spike gene target failure (SGTF) genomic signature is highly accurate for the identification of Alpha and Omicron SARS-CoV-2 variants. Sci. Rep. 2022, 12, 18968. [Google Scholar] [CrossRef] [PubMed]
- Pasteran, F.; de Mendieta, J.M.; Menocal, M.A.; Marchetti, P.; Olivieri, L.; Appendino, A.; Pereda, R.; Sangoy, A.; Kuzawka, M.; Tocho, E.; et al. Evaluation of the ability of the Gene Xpert Carba-R system to detect KPC and IMP carbapenemases variants. In Proceedings of the European Congress of Clinical Microbiology & Infectious Diseases, Barcelona, Spain, 27–30 April 2024; pp. 1–14. [Google Scholar]
- Escandón-Vargas, K.; Reyes, S.; Gutiérrez, S.; Villegas, M.V. The epidemiology of carbapenemases in Latin America and the Caribbean. Expert Rev. Anti. Infect. Ther. 2017, 15, 277–297. [Google Scholar] [CrossRef] [PubMed]
- van Duin, D.; Doi, Y. The global epidemiology of carbapenemase-producing Enterobacteriaceae. Virulence 2017, 8, 460–469. [Google Scholar] [CrossRef]
- Zeng, M.; Xia, J.; Zong, Z.; Shi, Y.; Ni, Y.; Hu, F.; Chen, Y.; Zhuo, C.; Hu, B.; Lv, X.; et al. Guidelines for the diagnosis, treatment, prevention and control of infections caused by carbapenem-resistant gram-negative bacilli. J. Microbiol. Immunol. Infect. 2023, 56, 653–671. [Google Scholar] [CrossRef] [PubMed]
- Wise, M.G.; Karlowsky, J.A.; Mohamed, N.; Hermsen, E.D.; Kamat, S.; Townsend, A.; Brink, A.; Soriano, A.; Paterson, D.L.; Moore, L.S.P.; et al. Global trends in carbapenem- and difficult-to-treat-resistance among World Health Organization priority bacterial pathogens: ATLAS surveillance program 2018–2022. J. Glob. Antimicrob. Resist. 2024, 37, 168–175. [Google Scholar] [CrossRef]
- Miranda Padilla, A.; Gómez Murillo, I.; González Solórzano, B.; Morales Rodríguez, Y. Primer hallazgo en Costa Rica de Enterobacteral portador de doble carbapenemasa: Un análisis desde el Laboratorio Clínico del Hospital México y su impacto para Latinoamérica—Revista del Colegio de Microbiólogos y. Químicos Clínicos de Costa Rica. Rev. Del Col. Microbiólogos Y Químicos Clínicos 2024, 29, 1-6a. [Google Scholar]
- Algarni, S.; Gudeta, D.D.; Han, J.; Nayak, R.; Foley, S.L. Genotypic analyses of IncHI2 plasmids from enteric bacteria. Sci. Rep. 2024, 14, 9802. [Google Scholar] [CrossRef]
- Lim, C.; Ashley, E.A.; Hamers, R.L.; Turner, P.; Kesteman, T.; Akech, S.; Corso, A.; Mayxay, M.; Okeke, I.N.; Limmathurotsakul, D.; et al. Surveillance strategies using routine microbiology for antimicrobial resistance in low- and middle-income countries. Clin. Microbiol. Infect. 2021, 27, 1391–1399. [Google Scholar] [CrossRef]
- Alamri, A.M.; Alkhilaiwi, F.A.; Ullah Khan, N. Era of Molecular Diagnostics Techniques before and after the COVID-19 Pandemic. Curr. Issues Mol. Biol. 2022, 44, 4769–4789. [Google Scholar] [CrossRef] [PubMed]
- Dien Bard, J.; Babady, N.E. The Successes and Challenges of SARS-CoV-2 Molecular Testing in the United States. Clin. Lab. Med. 2022, 42, 147. [Google Scholar] [CrossRef] [PubMed]
- Molina-Mora, J.A.; Cordero-Laurent, E.; Godínez, A.; Calderón-Osorno, M.; Brenes, H.; Soto-Garita, C.; Pérez-Corrales, C.; Drexler, J.F.; Moreira-Soto, A.; Corrales-Aguilar, E.; et al. SARS-CoV-2 genomic surveillance in Costa Rica: Evidence of a divergent population and an increased detection of a spike T1117I mutation. Infect. Genet. Evol. 2021, 92, 104872. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Strain | Immuno-Chromatography | MBL Based on Automated qPCR | Whole-Genome Sequencing |
|---|---|---|---|
| E. cloacae complex R36 | IMP-positive | IMP-negative | IMP-18-positive |
| P. aeruginosa R59 | IMP-positive | IMP-negative | IMP-18-positive |
| P. aeruginosa AG1 | Not tested | IMP-negative | IMP-18-positive |
| Gene | Name | Sequence | Length (pb) |
|---|---|---|---|
| IMP | IMP-F | GGAATAGAGTGGCTTAAYTCTC | 22 |
| IMP-R | CCAAACYACTASGTTATCT | 19 | |
| VIM | VIM-F | CGAAAAACACAGCGGCMCTTCT | 22 |
| VIM-R | GTGGAGACTGCACGCGTTAC | 20 | |
| 16S rRNA | 16S-F515 | GTGYCAGCMGCCGCGGTAA | 19 |
| 16S-R926 | CCGYCAATTYMTTTRAGTTT | 20 |
| Study ID | Species | Reception Date | Hospital and Region | MBL-Automated qPCR * | Custom qPCR | ||
|---|---|---|---|---|---|---|---|
| Ct IMP | Ct VIM | 16S rRNA | |||||
| AG1 | P. aeruginosa | 2010 | H1 metropolitan | VIM | 16.9 | 13.36 | + |
| RA | P. aeruginosa | 2015 | H1 metropolitan | VIM | 17.57 | 14.23 | + |
| RB | P. aeruginosa | 2015 | H1 metropolitan | VIM | 16.36 | 13.95 | + |
| R1 | K. pneumoniae | dic-21 | H7 metropolitan | NDM | - | - | + |
| R2 | A. baumannii | jul-22 | H7 metropolitan | NDM | - | - | + |
| R3 | S. fonticola | jul-22 | H7 metropolitan | NDM | - | - | + |
| R4 | P. putida | nov-22 | H1 metropolitan | VIM | 19.01 | 12.65 | + |
| R5 | S. marcescens | sept-23 | H1 metropolitan | NDM | - | - | + |
| R6 | K. pneumoniae | sept-23 | H1 metropolitan | NDM | - | - | + |
| R7 | K. pneumoniae | sept-23 | H1 metropolitan | NDM | - | - | + |
| R8 | E. coli | sept-23 | H1 metropolitan | NDM | - | - | + |
| R9 | P. mirabilis | sept-23 | H1 metropolitan | NDM | - | - | + |
| R10 | P. mirabilis | sept-23 | H1 metropolitan | NDM | - | - | + |
| R11 | S. marcescens | sept-23 | H1 metropolitan | NDM | - | - | + |
| R12 | P. aeruginosa | sept-23 | H1 metropolitan | VIM | 19.62 | 15.57 | + |
| R13 | K. pneumoniae | sept-23 | H1 metropolitan | NDM | - | - | + |
| R14 | K. pneumoniae | sept-23 | H1 metropolitan | NDM | - | - | + |
| R15 | K. pneumoniae | sept-23 | H1 metropolitan | NDM | - | - | + |
| R16 | P. aeruginosa | oct-23 | H1 metropolitan | VIM | 15.99 | 13.46 | + |
| R17 | P. aeruginosa | oct-23 | H1 metropolitan | VIM | 15.97 | 15.97 | + |
| R18 | A. baumannii | oct-23 | H1 metropolitan | NDM | - | - | + |
| R19 | K. pneumoniae | dic-23 | H7 metropolitan | No MBL | - | - | + |
| R20 | K. pneumoniae | ene-24 | H2 regional | No MBL | - | - | + |
| R21 | M. morganii | ene-24 | H2 regional | No MBL | - | - | + |
| R22 | E. coli | ene-24 | H2 regional | No MBL | - | - | + |
| R23 | E. coli | ene-24 | H2 regional | No MBL | - | - | + |
| R24 | E. cloacae complex | ene-24 | H2 regional | No MBL | - | - | + |
| R25 | P. mirabilis | ene-24 | H2 regional | No MBL | - | - | + |
| R26 | E. coli | ene-24 | H2 regional | No MBL | - | - | + |
| R27 | E. coli | ene-24 | H2 regional | No MBL | - | - | + |
| R28 | P. aeruginosa | feb-24 | H2 regional | No MBL | - | - | + |
| R29 | E. coli | feb-24 | H2 regional | No MBL | - | - | + |
| R30 | E. coli | feb-24 | H2 regional | No MBL | - | - | + |
| R31 | K. pneumoniae | feb-24 | H2 regional | No MBL | - | - | + |
| R32 | K. pneumoniae | feb-24 | H2 regional | No MBL | - | - | + |
| R33 | E. cloacae | mar-24 | H2 regional | No MBL | - | - | + |
| R34 | P. aeruginosa | jun-24 | H3 metropolitan | No MBL | - | - | + |
| R35 | P. aeruginosa | jun-24 | H3 metropolitan | No MBL | - | - | + |
| R36 | E. cloacae complex | jun-24 | H4 metropolitan | No MBL (IC: IMP-pos) | 18.24 | - | + |
| R37 | S. marcescens | jun-24 | H4 metropolitan | NDM | - | - | + |
| R38 | C. youngae | jun-24 | H4 metropolitan | NDM | - | - | + |
| R39 | S. marcescens | jun-24 | H4 metropolitan | NDM | - | - | + |
| R40 | K. aerogenes | jun-24 | H4 metropolitan | NDM | - | - | + |
| R41 | E. coli | jun-24 | H4 metropolitan | NDM | - | - | + |
| R42 | E. coli | jun-24 | H4 metropolitan | NDM | - | - | + |
| R43 | C. freundii | jun-24 | H4 metropolitan | NDM | - | - | + |
| R44 | E. cloacae complex | jun-24 | H4 metropolitan | NDM | - | - | + |
| R45 | K. pneumoniae | jun-24 | H4 metropolitan | NDM | - | - | + |
| R46 | K. pneumoniae | jun-24 | H4 metropolitan | NDM | - | - | + |
| R47 | K. aerogenes | jun-24 | H4 metropolitan | NDM | - | - | + |
| R48 | K. pneumoniae | jun-24 | H4 metropolitan | NDM | - | - | + |
| R49 | K. aerogenes | jun-24 | H4 metropolitan | NDM | - | - | + |
| R50 | C. youngae | jun-24 | H4 metropolitan | NDM | - | - | + |
| R51 | E. coli | jun-24 | H4 metropolitan | NDM | - | - | + |
| R52 | K. aerogenes | jun-24 | H4 metropolitan | NDM | - | - | + |
| R53 | K. aerogenes | jun-24 | H4 metropolitan | NDM | - | - | + |
| R54 | K. aerogenes | jun-24 | H4 metropolitan | NDM | - | - | + |
| R55 | K. oxytoca | jun-24 | H4 metropolitan | NDM | - | - | + |
| R56 | K. aerogenes | jun-24 | H4 metropolitan | NDM | - | - | + |
| R57 | E. coli | jun-24 | H3 metropolitan | No MBL | - | - | + |
| R58 | P. mirabilis | jun-24 | H3 metropolitan | No MBL | - | - | + |
| R59 | P. aeruginosa | jul-24 | H5 metropolitan | No MBL (IC: IMP-pos) | 18.16 | - | + |
| R60 | P. aeruginosa | jul-24 | H5 metropolitan | NT | 20.17 | - | + |
| R61 | P. aeruginosa | jul-24 | H3 metropolitan | VIM | 18.13 | 11.88 | + |
| R62 | P. aeruginosa | jul-24 | H3 metropolitan | No MBL | - | - | + |
| R63 | P. aeruginosa | jul-24 | H3 metropolitan | VIM | - | - | + |
| R64 | K. pneumoniae | ago-24 | H6 metropolitan | NT | - | - | + |
| R65 | P. aeruginosa | ago-24 | H6 metropolitan | NT | - | - | + |
| R66 | P. aeruginosa | ago-24 | H6 metropolitan | NT | - | - | + |
| R67 | K. pneumoniae | ago-24 | H6 metropolitan | NT | - | - | + |
| R68 | Salmonella sp | ago-24 | H6 metropolitan | NT | - | - | + |
| R69 | K. pneumoniae | ago-24 | H6 metropolitan | NT | - | - | + |
| R70 | K. aerogenes | ago-24 | H6 metropolitan | NT | - | - | + |
| R71 | P. aeruginosa | ago-24 | H3 metropolitan | No MBL | - | - | + |
| R72 | P. aeruginosa | sept-24 | H3 metropolitan | No MBL | - | - | + |
| R73 | P. aeruginosa | sept-24 | H3 metropolitan | No MBL | - | - | + |
| R74 | P. aeruginosa | sept-24 | H3 metropolitan | No MBL | - | - | + |
| R75 | E. coli | sept-24 | H7 metropolitan | No MBL | - | - | + |
| R76 | E. aerogenes | sept-24 | H7 metropolitan | No MBL | - | - | + |
| R77 | E. coli | sept-24 | H7 metropolitan | NDM | - | - | + |
| R78 | E. cloacae complex | oct-24 | H7 metropolitan | No MBL | - | - | + |
| R79 | K. aerogenes | oct-24 | H7 metropolitan | No MBL | - | - | + |
| R80 | E. coli | oct-24 | H8 regional | No MBL | - | - | + |
| R81 | K. pneumoniae | oct-24 | H8 regional | NDM | - | - | + |
| R82 | S. marcescens | oct-24 | H5 metropolitan | NDM | - | - | + |
| R83 | S. marcescens | oct-24 | H5 metropolitan | NDM | - | - | + |
| R84 | E. coli | oct-24 | H8 regional | NDM | - | - | + |
| R85 | K. pneumoniae | oct-24 | H8 regional | NDM | - | - | + |
| R86 | P. aeruginosa | nov-24 | H7 metropolitan | VIM (IC: VIM and IMP-pos) | 18.01 | 12.71 | + |
| R87 | E. cloacae | nov-24 | H6 metropolitan | NT | - | - | + |
| R88 | P. aeruginosa | nov-24 | H6 metropolitan | NT | 21.76 | 11.31 | + |
| R89 | K. pneumoniae | nov-24 | H5 metropolitan | NDM | - | - | + |
| R90 | P. aeruginosa | nov-24 | H5 metropolitan | VIM | 21.11 | 13.62 | + |
| R91 | M. morganii | nov-24 | H8 regional | NDM | - | - | + |
| R92 | S. marcescens | nov-24 | H8 regional | NDM | - | - | + |
| R93 | K. pneumoniae | nov-24 | H8 regional | NDM | - | - | + |
| R94 | E. coli | nov-24 | H8 regional | NDM | - | - | + |
| R95 | E. coli | nov-24 | H5 metropolitan | NDM | - | - | + |
| R96 | K. pneumoniae | nov-24 | H5 metropolitan | NDM | - | - | + |
| R97 | C. freundii | dic-24 | H5 metropolitan | NDM | - | - | + |
| R98 | E. coli | dic-24 | H5 metropolitan | NDM | - | - | + |
| R99 | K. aerogenes | ene-25 | H7 metropolitan | No MBL | - | - | + |
| R100 | C. freundii | ene-25 | H7 metropolitan | NDM | - | - | + |
| R101 | E. aerogenes | ene-25 | H7 metropolitan | NDM | - | - | + |
| R102 | S. fonticola | feb-25 | H7 metropolitan | NDM | - | - | + |
| R103 | E. coli | mar-25 | H7 metropolitan | NDM | - | - | + |
| R104 | K. pneumoniae | abr-25 | H8 regional | NDM | - | - | + |
| R105 | K. pneumoniae | abr-25 | H8 regional | NDM | - | - | + |
| R106 | E. coli | abr-25 | H8 regional | NDM | - | - | + |
| R107 | C. freundii | abr-25 | H8 regional | NDM | - | - | + |
| R108 | K. pneumoniae | abr-25 | H8 regional | NDM | - | - | + |
| R109 | K. pneumoniae | abr-25 | H8 regional | NDM | - | - | + |
| R110 | A. baumannii | may-25 | H6 metropolitan | NDM | - | - | + |
| R111 | A. baumannii | may-25 | H6 metropolitan | NDM | - | - | + |
| R112 | A. baumannii | may-25 | H6 metropolitan | NDM | - | - | + |
| R113 | A. baumannii | may-25 | H6 metropolitan | NDM | - | - | + |
| R114 | A. baumannii | may-25 | H6 metropolitan | NDM | - | - | + |
| R115 | K. pneumoniae | may-25 | H6 metropolitan | NT | - | - | + |
| R116 | E. cloacae complex | may-25 | H8 regional | NDM | - | - | + |
| Isolates | Phenotypic Profile * | Genomic Profile | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Species (Strain/ID) | Hospital (H) | Meropenem | Imipenem | qPCR | Whole-Genome Sequencing Analyses | ||||
| MIC (µg/mL) | Profile | MIC (µg/mL) | Profile | Ct IMP | Ct VIM | MLST | bla Genes | ||
| P. aeruginosa AG1 | H1 metropolitan | ≥16 | R | ≥16 | R | 16.90 | 13.36 | ST-111 | IMP-18, VIM-2, OXA-2, OXA-395, PDC-55 |
| P. aeruginosa RA | H1 metropolitan | ≥16 | R | ≥16 | R | 17.57 | 14.23 | ST-111 | IMP-18, VIM-2, OXA-2, OXA-395, PDC-55 |
| P. aeruginosa RB | H1 metropolitan | ≥16 | R | ≥16 | R | 16.36 | 13.95 | ST-111 | IMP-18, VIM-62, OXA-2, OXA-395, PDC-55 |
| P. putida R4 | H1 metropolitan | ≥16 | R | ≥16 | R | 19.01 | 12.65 | Unassigned | IMP-18, VIM-2 |
| P. aeruginosa R12 | H1 metropolitan | ≥16 | R | NT | NT | 19.62 | 15.57 | ST-111 | IMP-18, VIM-2, OXA-2, OXA-395, PDC-55 |
| P. aeruginosa R16 | H1 metropolitan | ≥16 | R | ≥16 | R | 15.99 | 13.46 | ST-111 | IMP-18, VIM-2, OXA-2, OXA-395, PDC-55 |
| P. aeruginosa R17 | H1 metropolitan | ≥16 | R | ≥16 | R | 15.97 | 15.97 | ST-111 | IMP-18, VIM-2, OXA-2, OXA-395, PDC-55 |
| E. cloacae complex R36 | H4 metropolitan | 8 | R | ≥16 | R | 18.24 | - | ST-90 | IMP-18, OXA-2, SHV-12 |
| P. aeruginosa R59 | H5 metropolitan | 8 | R | ≥16 | R | 18.16 | - | ST-179 | IMP-18, OXA-2, OXA-396, PDC-374 |
| P. aeruginosa R60 | H5 metropolitan | 2 | S | ≥16 | R | 20.17 | - | ST-179 | IMP-18, OXA-2, OXA-396, PDC-374 |
| P. aeruginosa R61 | H3 metropolitan | ≥16 | R | ≥16 | R | 18.13 | 11.88 | ST-111 | IMP-18, VIM-2, OXA-2, OXA-395, PDC-55 |
| P. aeruginosa R86 | H7 metropolitan | 4 | I | ≥16 | R | 18.01 | 12.71 | ST-111 | IMP-18, VIM-2, OXA-2, OXA-395, PDC-55 |
| P. aeruginosa R88 | H6 metropolitan | ≥16 | R | ≥16 | R | 21.76 | 11.31 | ST-111 | IMP-18, VIM-2, OXA-2, OXA-395, PDC-55 |
| P. aeruginosa R90 | H5 metropolitan | ≥16 | R | ≥16 | R | 21.11 | 13.62 | ST-111 | IMP-18, VIM-2, OXA-2, OXA-395, PDC-55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molina-Mora, J.A.; Rojas-Varela, Á.; Martínez-Arana, C.; Portilla-Victor, L.; Quirós-Fallas, I.; Sánchez-Fonseca, M.; Araya, X.; Cascante-Serrano, D.; Segura-Retana, E.; Espinoza-Solís, C.; et al. Finding the Missing IMP Gene: Overcoming the Imipenemase IMP Gene Drop-Out in Automated Molecular Testing for Carbapenem-Resistant Bacteria Circulating in Latin America. Antibiotics 2025, 14, 772. https://doi.org/10.3390/antibiotics14080772
Molina-Mora JA, Rojas-Varela Á, Martínez-Arana C, Portilla-Victor L, Quirós-Fallas I, Sánchez-Fonseca M, Araya X, Cascante-Serrano D, Segura-Retana E, Espinoza-Solís C, et al. Finding the Missing IMP Gene: Overcoming the Imipenemase IMP Gene Drop-Out in Automated Molecular Testing for Carbapenem-Resistant Bacteria Circulating in Latin America. Antibiotics. 2025; 14(8):772. https://doi.org/10.3390/antibiotics14080772
Chicago/Turabian StyleMolina-Mora, Jose Arturo, Ángel Rojas-Varela, Christopher Martínez-Arana, Lucia Portilla-Victor, Isaac Quirós-Fallas, Miryana Sánchez-Fonseca, Xavier Araya, Daniel Cascante-Serrano, Elvira Segura-Retana, Carlos Espinoza-Solís, and et al. 2025. "Finding the Missing IMP Gene: Overcoming the Imipenemase IMP Gene Drop-Out in Automated Molecular Testing for Carbapenem-Resistant Bacteria Circulating in Latin America" Antibiotics 14, no. 8: 772. https://doi.org/10.3390/antibiotics14080772
APA StyleMolina-Mora, J. A., Rojas-Varela, Á., Martínez-Arana, C., Portilla-Victor, L., Quirós-Fallas, I., Sánchez-Fonseca, M., Araya, X., Cascante-Serrano, D., Segura-Retana, E., Espinoza-Solís, C., Uribe-Calvo, M. J., Villalobos-Alfaro, V., Estrada-Murillo, H., Montoya-Madriz, S., Madrigal, W., Lizano, M., Lozada-Alvarado, S., Alvarado-Rodríguez, M., Bolaños-Muñoz, M., ... García-Santamaría, F. (2025). Finding the Missing IMP Gene: Overcoming the Imipenemase IMP Gene Drop-Out in Automated Molecular Testing for Carbapenem-Resistant Bacteria Circulating in Latin America. Antibiotics, 14(8), 772. https://doi.org/10.3390/antibiotics14080772