Comparative Effects of Intermittent vs. Constant Ceftiofur Hydrochloride Exposure on Staphylococcus aureus In Vitro


Abstract
1. Introduction
2. Results
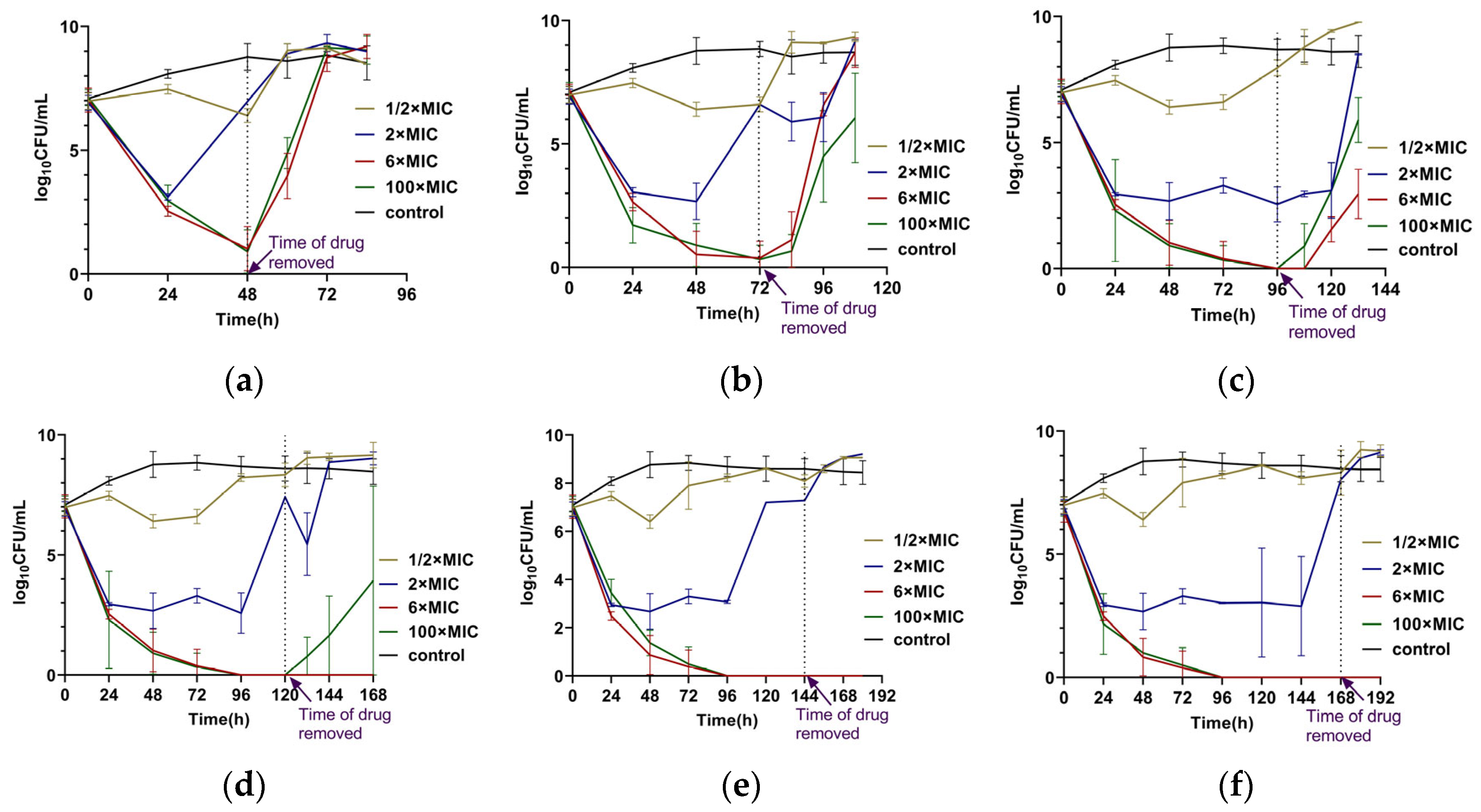
2.1. Results of Intermittent Exposure to CEF at 1/2 × MIC, 2 × MIC, 6× MIC, and 100 × MIC
2.2. Results of Single Prolonged Exposure to CEF for 48, 72, 96, 120, 144, and 168 h
2.3. Results of Genome Resequencing Analysis
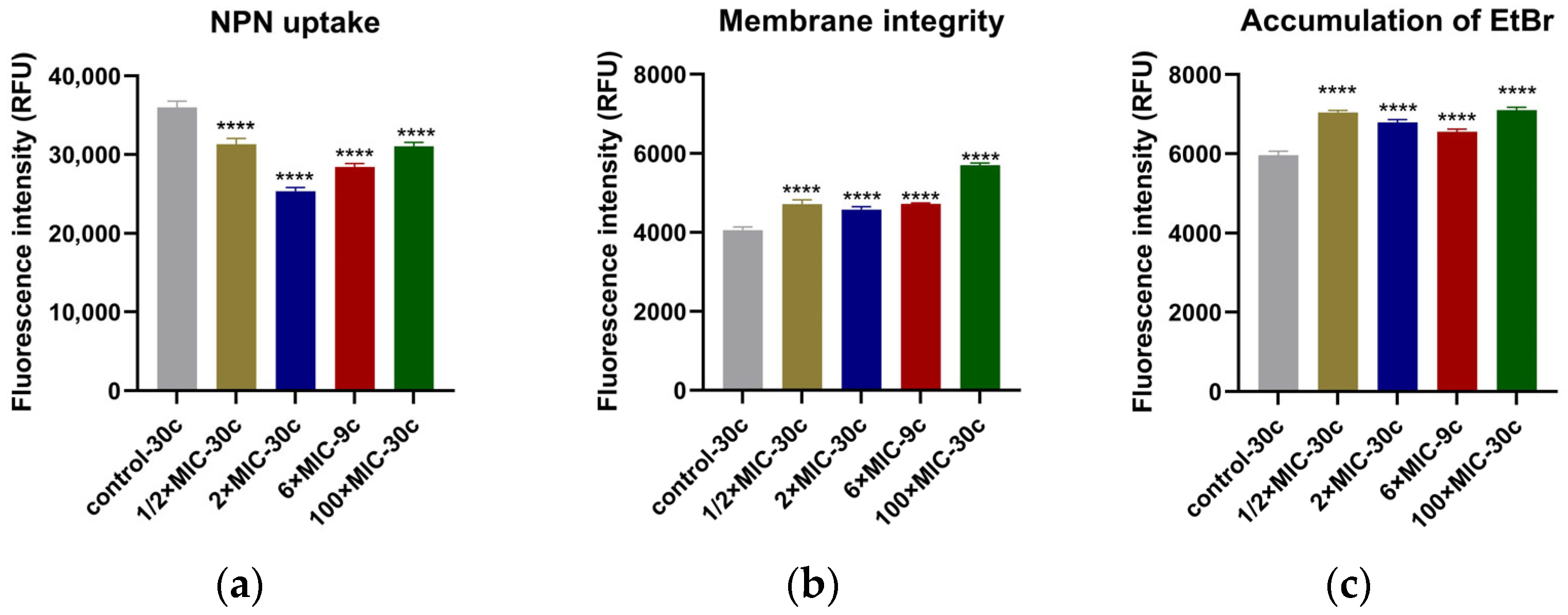
2.4. Results of Membrane Permeability, Membrane Integrity, and Efflux Pump Assays
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Culture Conditions, and Reagents
4.2. MIC, MBC, and MPC Determination
4.3. Intermittent CEF Exposure with Different Concentrations by In Vitro Adaptive Evolution Experiments
4.4. Single-Dose Administration of CEF with Different Durations and Concentrations
4.5. CFU Count and Survival Rate
4.6. Growth Curve Determination
4.7. Time–Kill Assay
4.8. Whole-Genome Resequencing Analysis
4.9. Membrane Permeability, Membrane Integrity, and Efflux Pump Assay
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CEF | Ceftiofur hydrochloride |
| BW | Body weight |
| S. aureus | Staphylococcus aureus |
| PBPs | Penicillin-binding proteins |
| MIC | Minimum inhibitory concentration |
| MBC | Minimum bactericidal concentration |
| MPC | Mutant prevention concentration |
| MDK99 | Minimum duration required to kill 99% of bacterial cells in the population |
| MDK99.99 | Minimum duration required to kill 99.99% of bacterial cells in the population |
| CDS | Coding sequence |
| SNP | Single-nucleotide polymorphism |
| SV | Structural variation |
| NPN | 1-N-phenyl-naphthylamine |
| PI | Propidium iodide |
| EtBr | Ethidium bromide |
| TSB | Tryptic soy broth |
| TSA | Tryptic soy agar |
| MH | Mueller–Hinton |
| MPCpr | Provisional mutant prevention concentration |
References
- Chenault, J.R.; McAllister, J.F.; Chester, S.T., Jr.; Dame, K.J.; Kausche, F.M.; Robb, E.J. Efficacy of ceftiofur hydrochloride sterile suspension administered parenterally for the treatment of acute postpartum metritis in dairy cows. J. Am. Vet. Med. Assoc. 2004, 224, 1634–1639. [Google Scholar] [CrossRef] [PubMed]
- An, B.; Xu, X.; Ma, W.; Huo, M.; Wang, H.; Liu, Z.; Cheng, G.; Huang, L. The adsorption-desorption characteristics and degradation kinetics of ceftiofur in different agricultural soils. Ecotoxicol. Environ. Saf. 2021, 222, 112503. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration. Available online: https://animaldrugsatfda.fda.gov/adafda/views/#/home/searchResult (accessed on 10 May 2025).
- von Krueger, X.; Scherpenisse, P.; Roiger, S.; Heuwieser, W. Determination of ceftiofur derivatives in serum, endometrial tissue, and lochia in puerperal dairy cows with fever or acute puerperal metritis after subcutaneous administration of ceftiofur crystalline free acid. J. Dairy Sci. 2013, 96, 1054–1062. [Google Scholar] [CrossRef]
- Goranova, M.; Ochoa, G.; Maier, P.; Hoyle, A. Evolutionary optimisation of antibiotic dosing regimens for bacteria with different levels of resistance. Artif. Intell. Med. 2022, 133, 102405. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.N.; Papich, M.G.; Drusano, G.L. Dosing regimen matters: The importance of early intervention and rapid attainment of the pharmacokinetic/pharmacodynamic target. Antimicrob. Agents Chemother. 2012, 56, 2795–2805. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Z.; Mehrotra, M.; Ghimire, S.; Adewoye, L. beta-Lactam resistance and beta-lactamases in bacteria of animal origin. Vet. Microbiol. 2007, 121, 197–214. [Google Scholar] [CrossRef]
- Burgess, D.S. Pharmacodynamic principles of antimicrobial therapy in the prevention of resistance. Chest 1999, 115, 19s–23s. [Google Scholar] [CrossRef]
- McNabb, J.J.; Nightingale, C.H.; Quintiliani, R.; Nicolau, D.P. Cost-effectiveness of ceftazidime by continuous infusion versus intermittent infusion for nosocomial pneumonia. Pharmacotherapy 2001, 21, 549–555. [Google Scholar] [CrossRef]
- Lewis, K. Persister cells. Annu. Rev. Microbiol. 2010, 64, 357–372. [Google Scholar] [CrossRef]
- Fisher, R.A.; Gollan, B.; Helaine, S. Persistent bacterial infections and persister cells. Nat. Rev. Microbiol. 2017, 15, 453–464. [Google Scholar] [CrossRef]
- Levin-Reisman, I.; Ronin, I.; Gefen, O.; Braniss, I.; Shoresh, N.; Balaban, N.Q. Antibiotic tolerance facilitates the evolution of resistance. Science 2017, 355, 826–830. [Google Scholar] [CrossRef]
- LeBlanc, S.J.; Duffield, T.F.; Leslie, K.E.; Bateman, K.G.; Keefe, G.P.; Walton, J.S.; Johnson, W.H. The effect of treatment of clinical endometritis on reproductive performance in dairy cows. J. Dairy Sci. 2002, 85, 2237–2249. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Pei, L.; Shen, Y.; Wu, J.; Qian, Y.; Zhang, N.; Mao, W.; Cao, J. Prostaglandin E2 accumulation is closely associated with S. aureus-infected bovine endometritis. Cytokine 2024, 175, 156498. [Google Scholar] [CrossRef]
- Freitas, C.H.; Mendes, J.F.; Villarreal, P.V.; Santos, P.R.; Gonçalves, C.L.; Gonzales, H.L.; Nascente, P.S. Identification and antimicrobial suceptibility profile of bacteria causing bovine mastitis from dairy farms in Pelotas, Rio Grande do Sul. Braz. J. Biol. Rev. Brasleira Biol. 2018, 78, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Wilke, M.S.; Lovering, A.L.; Strynadka, N.C. Beta-lactam antibiotic resistance: A current structural perspective. Curr. Opin. Microbiol. 2005, 8, 525–533. [Google Scholar] [CrossRef]
- Fridman, O.; Goldberg, A.; Ronin, I.; Shoresh, N.; Balaban, N.Q. Optimization of lag time underlies antibiotic tolerance in evolved bacterial populations. Nature 2014, 513, 418–421. [Google Scholar] [CrossRef] [PubMed]
- Balaban, N.Q.; Helaine, S.; Lewis, K.; Ackermann, M.; Aldridge, B.; Andersson, D.I.; Brynildsen, M.P.; Bumann, D.; Camilli, A.; Collins, J.J.; et al. Definitions and guidelines for research on antibiotic persistence. Nat. Rev. Microbiol. 2019, 17, 441–448. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Park, I.; Khan, I.; Alshammari, F.H.; Kumar, P.; Chen, L.; Na, M.; Huh, Y.S.; Han, Y.-K.; Shukla, S. (−)-Tetrahydroberberrubine∙acetate accelerates antioxidant potential and inhibits food associated Bacillus cereus in rice. Food Chem. 2021, 339, 127902. [Google Scholar] [CrossRef]
- Ahmed, M.N.; Porse, A.; Sommer, M.O.A.; Høiby, N.; Ciofu, O. Evolution of Antibiotic Resistance in Biofilm and Planktonic Pseudomonas aeruginosa Populations Exposed to Subinhibitory Levels of Ciprofloxacin. Antimicrob. Agents Chemother. 2018, 62, e00320-18. [Google Scholar] [CrossRef]
- Santos-Lopez, A.; Marshall, C.W.; Scribner, M.R.; Snyder, D.J.; Cooper, V.S. Evolutionary pathways to antibiotic resistance are dependent upon environmental structure and bacterial lifestyle. eLife 2019, 8, e47612. [Google Scholar] [CrossRef]
- Trampari, E.; Holden, E.R.; Wickham, G.J.; Ravi, A.; Martins, L.O.; Savva, G.M.; Webber, M.A. Exposure of Salmonella biofilms to antibiotic concentrations rapidly selects resistance with collateral tradeoffs. NPJ Biofilms Microbiomes 2021, 7, 3. [Google Scholar] [CrossRef]
- Miller, C.; Kong, J.; Tran, T.T.; Arias, C.A.; Saxer, G.; Shamoo, Y. Adaptation of Enterococcus faecalis to daptomycin reveals an ordered progression to resistance. Antimicrob. Agents Chemother. 2013, 57, 5373–5383. [Google Scholar] [CrossRef] [PubMed]
- Scribner, M.R.; Santos-Lopez, A.; Marshall, C.W.; Deitrick, C.; Cooper, V.S. Parallel Evolution of Tobramycin Resistance across Species and Environments. mBio 2020, 11, e00932-20. [Google Scholar] [CrossRef]
- France, M.T.; Cornea, A.; Kehlet-Delgado, H.; Forney, L.J. Spatial structure facilitates the accumulation and persistence of antibiotic-resistant mutants in biofilms. Evol. Appl. 2019, 12, 498–507. [Google Scholar] [CrossRef]
- Usui, M.; Yoshii, Y.; Thiriet-Rupert, S.; Ghigo, J.M.; Beloin, C. Intermittent antibiotic treatment of bacterial biofilms favors the rapid evolution of resistance. Commun. Biol. 2023, 6, 275. [Google Scholar] [CrossRef] [PubMed]
- The Upjohn Company; EXCENEL® Sterile Suspension; U.S. Food and Drug Administration. 1996. Available online: https://animaldrugsatfda.fda.gov/adafda/app/search/public/document/downloadFoi/516 (accessed on 10 May 2025).
- Roberts, J.A.; Kruger, P.; Paterson, D.L.; Lipman, J. Antibiotic resistance—What’s dosing got to do with it? Crit. Care Med. 2008, 36, 2433–2440. [Google Scholar] [CrossRef]
- Michiels, J.E.; Van den Bergh, B.; Verstraeten, N.; Fauvart, M.; Michiels, J. In Vitro Emergence of High Persistence upon Periodic Aminoglycoside Challenge in the ESKAPE Pathogens. Antimicrob. Agents Chemother. 2016, 60, 4630–4637. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X. Clarification of MPC and the mutant selection window concept. J. Antimicrob. Chemother. 2003, 52, 731. [Google Scholar] [CrossRef]
- Hu, Y.; Zhu, H.; Zhang, X.; Wu, Y.; Li, J.; Li, N.; Cai, Z.; Yang, Y. Adaptive Resistance of Staphylococcus aureus to Cefquinome Sulfate in an In Vitro Pharmacokinetic Model with Transcriptomic Insights. Microorganisms 2025, 13, 329. [Google Scholar] [CrossRef]
- Mouton, J.W.; den Hollander, J.G. Killing of Pseudomonas aeruginosa during continuous and intermittent infusion of ceftazidime in an in vitro pharmacokinetic model. Antimicrob. Agents Chemother. 1994, 38, 931–936. [Google Scholar] [CrossRef]
- Vasco, K.A.; Carbonell, S.; Sloup, R.E.; Bowcutt, B.; Colwell, R.R.; Graubics, K.; Erskine, R.; Norby, B.; Ruegg, P.L.; Zhang, L.; et al. Persistent effects of intramammary ceftiofur treatment on the gut microbiome and antibiotic resistance in dairy cattle. Anim. Microbiome 2023, 5, 56. [Google Scholar] [CrossRef]
- Elsayed, M.G.; Elkomy, A.A.; Gaballah, M.S.; Elbadawy, M. Nephrotoxicity of cefepime: A new cephalosporin antibiotic in rats. J. Pharmacol. Pharmacother. 2014, 5, 33–38. [Google Scholar] [CrossRef]
- Zapun, A.; Contreras-Martel, C.; Vernet, T. Penicillin-binding proteins and beta-lactam resistance. FEMS Microbiol. Rev. 2008, 32, 361–385. [Google Scholar] [CrossRef] [PubMed]
- Helander, I.M.; Mattila-Sandholm, T. Fluorometric assessment of gram-negative bacterial permeabilization. J. Appl. Microbiol. 2000, 88, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Timms, A.R.; Steingrimsdottir, H.; Lehmann, A.R.; Bridges, B.A. Mutant sequences in the rpsL gene of Escherichia coli B/r: Mechanistic implications for spontaneous and ultraviolet light mutagenesis. Mol. Gen. Genet. MGG 1992, 232, 89–96. [Google Scholar] [CrossRef]
- Chan, L.C.; Basuino, L.; Diep, B.; Hamilton, S.; Chatterjee, S.S.; Chambers, H.F. Ceftobiprole- and ceftaroline-resistant methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2015, 59, 2960–2963. [Google Scholar] [CrossRef] [PubMed]
- Rimbara, E.; Noguchi, N.; Kawai, T.; Sasatsu, M. Mutations in penicillin-binding proteins 1, 2 and 3 are responsible for amoxicillin resistance in Helicobacter pylori. J. Antimicrob. Chemother. 2008, 61, 995–998. [Google Scholar] [CrossRef]
- Yin, Y.; Chen, H.; Li, S.; Gao, H.; Sun, S.; Li, H.; Wang, R.; Jin, L.; Liu, Y.; Wang, H. Daptomycin resistance in methicillin-resistant Staphylococcus aureus is conferred by IS256 insertion in the promoter of mprF along with mutations in mprF and walK. Int. J. Antimicrob. Agents 2019, 54, 673–680. [Google Scholar] [CrossRef]
- Ho, C.M.; Li, C.Y.; Ho, M.W.; Lin, C.Y.; Liu, S.H.; Lu, J.J. High rate of qacA- and qacB-positive methicillin-resistant Staphylococcus aureus isolates from chlorhexidine-impregnated catheter-related bloodstream infections. Antimicrob. Agents Chemother. 2012, 56, 5693–5697. [Google Scholar] [CrossRef]
- Nong, Y.; Steinig, E.; Pollock, G.L.; Taiaroa, G.; Carter, G.P.; Monk, I.R.; Pang, S.; Daley, D.A.; Coombs, G.W.; Forde, B.M.; et al. Emergence and clonal expansion of a qacA-harbouring sequence type 45 lineage of methicillin-resistant Staphylococcus aureus. Commun. Biol. 2024, 7, 349. [Google Scholar] [CrossRef]
- Rajesh, T.; Song, E.; Kim, J.N.; Lee, B.R.; Kim, E.J.; Park, S.H.; Kim, Y.G.; Yoo, D.; Park, H.Y.; Choi, Y.H.; et al. Inactivation of phosphomannose isomerase gene abolishes sporulation and antibiotic production in Streptomyces coelicolor. Appl. Microbiol. Biotechnol. 2012, 93, 1685–1693. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Zhang, Y.; Wang, C.; Hu, D.; Liu, J.; Chen, L.; Shi, F.; Tang, X.; Hao, Z.; Suo, J.; et al. EtcPRS (Mut) as a molecular marker of halofuginone resistance in Eimeria tenella and Toxoplasma gondii. iScience 2023, 26, 106334. [Google Scholar] [CrossRef]
- Pankey, G.A.; Sabath, L.D. Clinical relevance of bacteriostatic versus bactericidal mechanisms of action in the treatment of Gram-positive bacterial infections. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2004, 38, 864–870. [Google Scholar] [CrossRef]
- Blondeau, J.M.; Zhao, X.; Hansen, G.; Drlica, K. Mutant prevention concentrations of fluoroquinolones for clinical isolates of Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2001, 45, 433–438. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Liu, D.; Ashokkumar, M.; Ye, X.; Jin, T.Z.; Guo, M. Antibacterial mechanism of ultrasound against Escherichia coli: Alterations in membrane microstructures and properties. Ultrason. Sonochem. 2021, 73, 105509. [Google Scholar] [CrossRef]
- Singh, V.; Nandanwar, H. IMT-P8 potentiates Gram-positive specific antibiotics in intrinsically resistant Gram-negative bacteria. Antimicrob. Agents Chemother. 2024, 68, e0075324. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jia, Y.; Yang, K.; Tong, Z.; Shi, J.; Li, R.; Xiao, X.; Ren, W.; Hardeland, R.; Reiter, R.J.; et al. Melatonin overcomes MCR-mediated colistin resistance in Gram-negative pathogens. Theranostics 2020, 10, 10697–10711. [Google Scholar] [CrossRef]
- Song, M.; Liu, Y.; Huang, X.; Ding, S.; Wang, Y.; Shen, J.; Zhu, K. A broad-spectrum antibiotic adjuvant reverses multidrug-resistant Gram-negative pathogens. Nat. Microbiol. 2020, 5, 1040–1050. [Google Scholar] [CrossRef]
- Paixão, L.; Rodrigues, L.; Couto, I.; Martins, M.; Fernandes, P.; de Carvalho, C.C.; Monteiro, G.A.; Sansonetty, F.; Amaral, L.; Viveiros, M. Fluorometric determination of ethidium bromide efflux kinetics in Escherichia coli. J. Biol. Eng. 2009, 3, 18. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Sample | Clean Data | Q20% 1 | Q30% 2 | Mapped Reads | Mapped Rate (%) | Coverage (%) | Aver-Dep 3 |
|---|---|---|---|---|---|---|---|
| 1/2 × MIC | 7,590,116 | 98.56 | 96.6 | 7,570,381 | 99.74 | 99.97 | 408.0 |
| 2 × MIC | 7,721,716 | 98.57 | 96.67 | 7,690,056 | 99.59 | 99.97 | 414.0 |
| 6 × MIC | 8,540,206 | 98.52 | 96.45 | 8,516,293 | 99.72 | 99.97 | 458.9 |
| 100 × MIC | 7,654,960 | 98.54 | 96.52 | 7,636,588 | 99.76 | 99.97 | 411.6 |
| Sample | Type | Annotation of Coding Gene |
|---|---|---|
| 1/2 × MIC | stoploss | |
| 2 × MIC | nonsynonymous | Multimodular transpeptidase-transglycosylase/Penicillin-binding protein 1A/1B (PBP1) |
| nonsynonymous | Multimodular transpeptidase-transglycosylase/Penicillin-binding protein 1A/1B (PBP1) | |
| nonsynonymous | Signal transduction histidine kinase | |
| synonymous | RGD-containing lipoprotein | |
| nonsynonymous | Drug resistance transporter, EmrB/QacA subfamily | |
| 6 × MIC | nonsynonymous | Hypothetical protein |
| nonsynonymous | Mannose-6-phosphate isomerase | |
| 100 × MIC | nonsynonymous | Prolyl-tRNA synthetase, bacterial type |
| stoploss | Phosphoesterase, DHH family protein | |
| stoploss | Phosphoesterase, DHH family protein | |
| synonymous | LSU m5C1962 methyltransferase RlmI | |
| synonymous | Membrane component of multidrug resistance system | |
| nonsynonymous | Drug resistance transporter, EmrB/QacA subfamily |
| Sample | Inter-Chromosomal Translocation | Deletion | Insertion | Inversion | Intra-Chromosomal Translocation | Total |
|---|---|---|---|---|---|---|
| 1/2 × MIC | 42 | 0 | 6 | 0 | 2 | 50 |
| 2 × MIC | 43 | 1 | 1 | 0 | 4 | 49 |
| 6 × MIC | 44 | 1 | 3 | 0 | 5 | 53 |
| 100 × MIC | 43 | 1 | 1 | 0 | 3 | 48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Li, C.; Guo, F.; Yu, Z. Comparative Effects of Intermittent vs. Constant Ceftiofur Hydrochloride Exposure on Staphylococcus aureus In Vitro. Antibiotics 2025, 14, 686. https://doi.org/10.3390/antibiotics14070686
Wang J, Li C, Guo F, Yu Z. Comparative Effects of Intermittent vs. Constant Ceftiofur Hydrochloride Exposure on Staphylococcus aureus In Vitro. Antibiotics. 2025; 14(7):686. https://doi.org/10.3390/antibiotics14070686
Chicago/Turabian StyleWang, Junli, Chongyang Li, Fanxi Guo, and Zugong Yu. 2025. "Comparative Effects of Intermittent vs. Constant Ceftiofur Hydrochloride Exposure on Staphylococcus aureus In Vitro" Antibiotics 14, no. 7: 686. https://doi.org/10.3390/antibiotics14070686
APA StyleWang, J., Li, C., Guo, F., & Yu, Z. (2025). Comparative Effects of Intermittent vs. Constant Ceftiofur Hydrochloride Exposure on Staphylococcus aureus In Vitro. Antibiotics, 14(7), 686. https://doi.org/10.3390/antibiotics14070686