
The Co-Existence of mcr-1.1 and mcr-3.5 in Escherichia coli Isolated from Clinical Samples in Thailand

 , , , , and
, , , , and
Abstract
1. Introduction
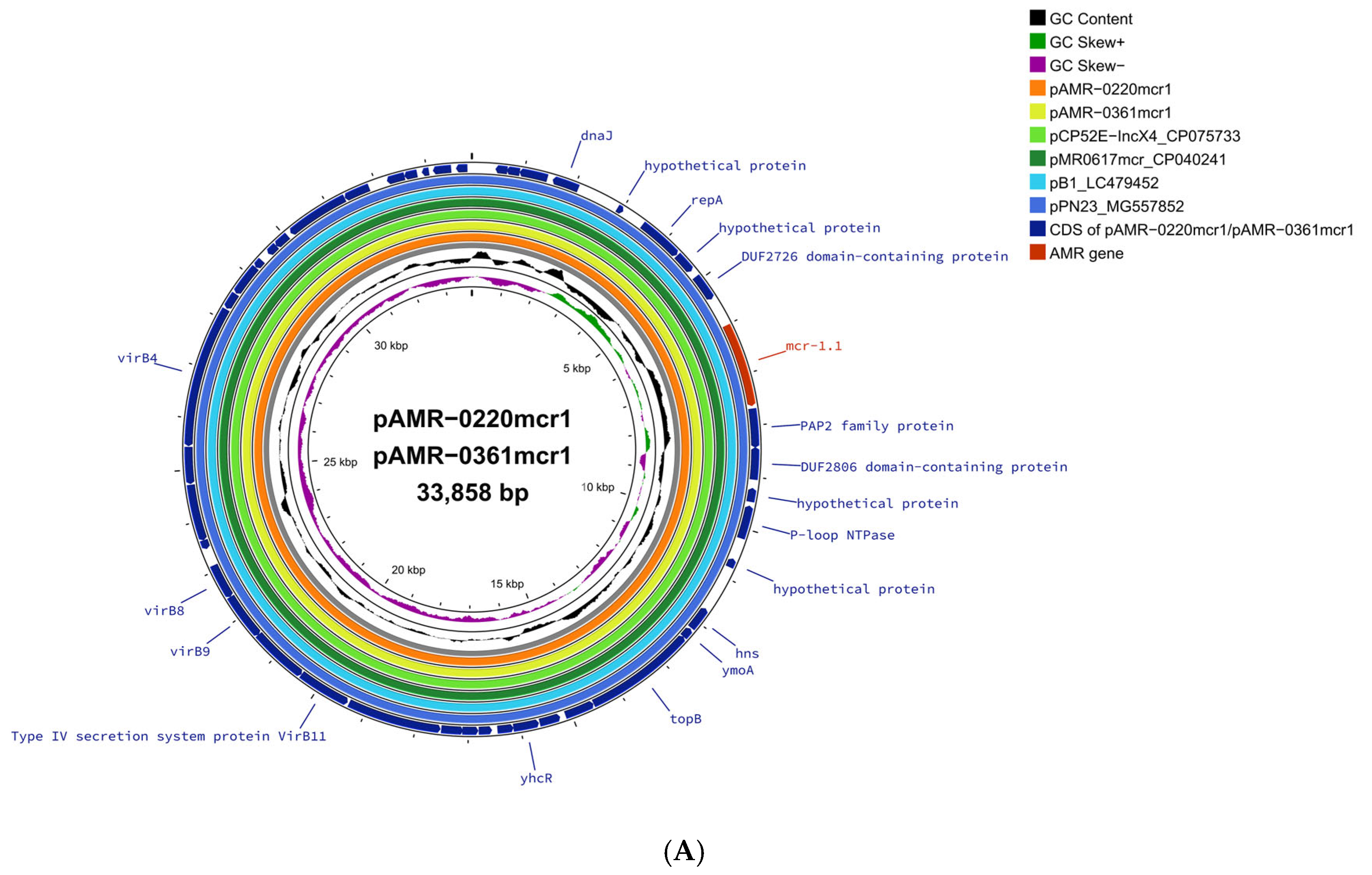
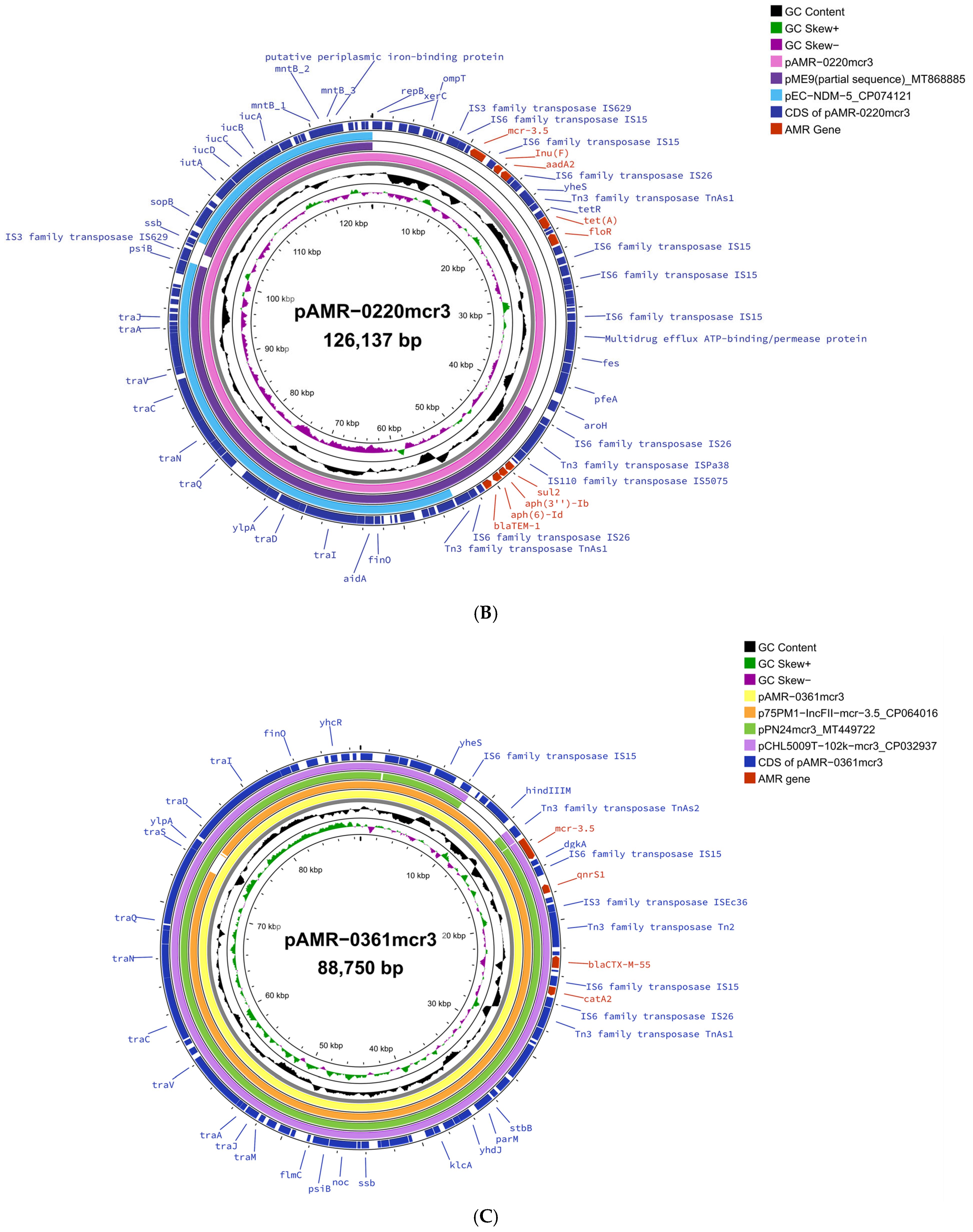
2. Results and Discussion
3. Material and Methods
3.1. Bacterial Strains and Antibiotic Susceptibility Testing
3.2. Detection of Extended Spectrum Beta-Lactamase (ESBL), Carbapenemase, and Colistin Resistance Genes
3.3. Short- and Long-Read Whole-Genome Sequencing
3.4. Analysis of Whole-Genome Sequencing Data
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- El-Sayed Ahmed, M.A.E.; Zhong, L.L.; Shen, C.; Yang, Y.; Doi, Y.; Tian, G.B. Colistin and its role in the Era of antibiotic resistance: An extended review (2000–2019). Emerg. Microbes Infect. 2020, 9, 868–885. [Google Scholar] [CrossRef]
- Zellweger, R.M.; Carrique-Mas, J.; Limmathurotsakul, D.; Day, N.P.J.; Thwaites, G.E.; Baker, S.; Southeast Asia Antimicrobial Resistance Network. A current perspective on antimicrobial resistance in Southeast Asia. J. Antimicrob. Chemother. 2017, 72, 2963–2972. [Google Scholar] [CrossRef] [PubMed]
- HPSR-AMR. Highlights Thailand One Health Report 2022: Antimicrobial Consumption and Antimicrobial Resistance. 2025. Available online: https://www.thaiamrwatch.net/materials/92 (accessed on 30 April 2025).
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Hussein, N.H.; AL-Kadmy, I.M.S.; Taha, B.M.; Hussein, J.D. Mobilized colistin resistance (mcr) genes from 1 to 10: A comprehensive review. Mol. Biol. Rep. 2021, 48, 2897–2907. [Google Scholar] [CrossRef]
- Olaitan, A.O.; Chabou, S.; Okdah, L.; Morand, S.; Rolain, J.M. Dissemination of the mcr-1 colistin resistance gene. Lancet Infect. Dis. 2016, 16, 147. [Google Scholar] [CrossRef]
- Phuadraksa, T.; Wichit, S.; Arikit, S.; Songtawee, N.; Yainoy, S. Co-occurrence of mcr-2 and mcr-3 genes on chromosome of multidrug-resistant Escherichia coli isolated from healthy individuals in Thailand. Int. J. Antimicrob. Agents 2022, 60, 106662. [Google Scholar] [CrossRef] [PubMed]
- Khanawapee, A.; Kerdsin, A.; Chopjitt, P.; Boueroy, P.; Hatrongjit, R.; Akeda, Y.; Tomono, K.; Nuanualsuwan, S.; Hamada, S. Distribution and Molecular Characterization of Escherichia coli Harboring mcr Genes Isolated from Slaughtered Pigs in Thailand. Microb. Drug Resist. 2021, 27, 971–979. [Google Scholar] [CrossRef]
- Paveenkittiporn, W.; Kamjumphol, W.; Ungcharoen, R.; Kerdsin, A. Whole-Genome Sequencing of Clinically Isolated Carbapenem-Resistant Enterobacterales Harboring mcr Genes in Thailand, 2016–2019. Front. Microbiol. 2020, 11, 586368. [Google Scholar] [CrossRef]
- Liu, L.; Feng, Y.; Zhang, X.; McNally, A.; Zong, Z. New Variant of mcr-3 in an Extensively Drug-Resistant Escherichia coli Clinical Isolate Carrying mcr-1 and blaNDM-5. Antimicrob. Agents Chemother. 2017, 61, e01757-17. [Google Scholar] [CrossRef]
- Creighton, J.; Anderson, T.; Howard, J.; Dyet, K.; Ren, X.; Freeman, J. Co-occurrence of mcr-1 and mcr-3 genes in a single Escherichia coli in New Zealand. J. Antimicrob. Chemother. 2019, 74, 3113–3116. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, P.; Yang, X.; Wang, Z.; Fanning, S.; Wang, J.; Du, P.; Bai, L. Identification of a novel hybrid plasmid coproducing MCR-1 and MCR-3 variant from an Escherichia coli strain. J. Antimicrob. Chemother. 2019, 74, 1517–1520. [Google Scholar] [CrossRef]
- Hernandez, M.; Iglesias, M.R.; Rodriguez-Lazaro, D.; Gallardo, A.; Quijada, N.; Miguela-Villoldo, P.; Campos, M.J.; Piriz, S.; Lopez-Orozco, G.; de Frutos, C.; et al. Co-occurrence of colistin-resistance genes mcr-1 and mcr-3 among multidrug-resistant Escherichia coli isolated from cattle, Spain, September 2015. Euro Surveill. 2017, 22, 30586. [Google Scholar] [CrossRef] [PubMed]
- Hamame, A.; Davoust, B.; Rolain, J.M.; Diene, S.M. Genomic characterisation of an mcr-1 and mcr-3-producing Escherichia coli strain isolated from pigs in France. J. Glob. Antimicrob. Resist. 2022, 28, 174–179. [Google Scholar] [CrossRef]
- Yu, Y.; Andrey, D.O.; Yang, R.S.; Sands, K.; Tansawai, U.; Li, M.; Portal, E.; Gales, A.C.; Niumsup, P.R.; Sun, J.; et al. A Klebsiella pneumoniae strain co-harbouring mcr-1 and mcr-3 from a human in Thailand. J. Antimicrob. Chemother. 2020, 75, 2372–2374. [Google Scholar] [CrossRef] [PubMed]
- Leangapichart, T.; Stosic, M.S.; Hickman, R.A.; Lunha, K.; Jiwakanon, J.; Angkititrakul, S.; Magnusson, U.; Van Boeckel, T.P.; Jarhult, J.D.; Sunde, M. Exploring the epidemiology of mcr genes, genetic context and plasmids in Enterobacteriaceae originating from pigs and humans on farms in Thailand. J. Antimicrob. Chemother. 2023, 78, 1395–1405. [Google Scholar] [CrossRef]
- Roer, L.; Overballe-Petersen, S.; Hansen, F.; Schonning, K.; Wang, M.; Roder, B.L.; Hansen, D.S.; Justesen, U.S.; Andersen, L.P.; Fulgsang-Damgaard, D.; et al. Escherichia coli Sequence Type 410 Is Causing New International High-Risk Clones. mSphere 2018, 3, e00337-18. [Google Scholar] [CrossRef]
- Dadashi, M.; Sameni, F.; Bostanshirin, N.; Yaslianifard, S.; Khosravi-Dehaghi, N.; Nasiri, M.J.; Goudarzi, M.; Hashemi, A.; Hajikhani, B. Global prevalence and molecular epidemiology of mcr-mediated colistin resistance in Escherichia coli clinical isolates: A systematic review. J. Glob. Antimicrob. Resist. 2022, 29, 444–461. [Google Scholar] [CrossRef]
- Ruekit, S.; Srijan, A.; Serichantalergs, O.; Margulieux, K.R.; Mc Gann, P.; Mills, E.G.; Stribling, W.C.; Pimsawat, T.; Kormanee, R.; Nakornchai, S.; et al. Molecular characterization of multidrug-resistant ESKAPEE pathogens from clinical samples in Chonburi, Thailand (2017–2018). BMC Infect. Dis. 2022, 22, 695. [Google Scholar] [CrossRef]
- Srijan, A.; Margulieux, K.R.; Ruekit, S.; Snesrud, E.; Maybank, R.; Serichantalergs, O.; Kormanee, R.; Sukhchat, P.; Sriyabhaya, J.; Hinkle, M.; et al. Genomic Characterization of Nonclonal mcr-1-Positive Multidrug-Resistant Klebsiella pneumoniae from Clinical Samples in Thailand. Microb. Drug Resist. 2018, 24, 403–410. [Google Scholar] [CrossRef]
- Khine, N.O.; Wongsurawat, T.; Jenjaroenpun, P.; Hampson, D.J.; Prapasarakul, N. Comparative genomic analysis of Colistin resistant Escherichia coli isolated from pigs, a human and wastewater on colistin withdrawn pig farm. Sci. Rep. 2023, 13, 5124. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, W.; Tanaka, H.; Taniguchi, Y.; Iimura, M.; Soga, E.; Kubo, R.; Matsuo, N.; Kawamura, K.; Arakawa, Y.; Nagano, Y.; et al. Acquisition of mcr-1 and Cocarriage of Virulence Genes in Avian Pathogenic Escherichia coli Isolates from Municipal Wastewater Influents in Japan. Appl. Environ. Microbiol. 2019, 85, e01661-19. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Li, M.; Spiller, O.B.; Andrey, D.O.; Hinchliffe, P.; Li, H.; MacLean, C.; Niumsup, P.; Powell, L.; Pritchard, M.; et al. Balancing mcr-1 expression and bacterial survival is a delicate equilibrium between essential cellular defence mechanisms. Nat. Commun. 2017, 8, 2054. [Google Scholar] [CrossRef] [PubMed]
- Boueroy, P.; Wongsurawat, T.; Jenjaroenpun, P.; Chopjitt, P.; Hatrongjit, R.; Jittapalapong, S.; Kerdsin, A. Plasmidome in mcr-1 harboring carbapenem-resistant enterobacterales isolates from human in Thailand. Sci. Rep. 2022, 12, 19051. [Google Scholar] [CrossRef]
- Matamoros, S.; van Hattem, J.M.; Arcilla, M.S.; Willemse, N.; Melles, D.C.; Penders, J.; Vinh, T.N.; Thi Hoa, N.; Bootsma, M.C.J.; van Genderen, P.J.; et al. Global phylogenetic analysis of Escherichia coli and plasmids carrying the mcr-1 gene indicates bacterial diversity but plasmid restriction. Sci. Rep. 2017, 7, 15364. [Google Scholar] [CrossRef]
- Huang, J.; Zhu, J.; Gong, D.; Wu, L.; Zhu, Y.; Hu, L. Whole genome sequence of EC16, a bla(NDM-5)-, bla(CTX-M-55)-, and fosA3-coproducing Escherichia coli ST167 clinical isolate from China. J. Glob. Antimicrob. Resist. 2022, 29, 296–298. [Google Scholar] [CrossRef]
- Shi, J.P.; Zhu, H.; Liu, C.; Xie, H.; Li, C.C.; Cao, X.L.; Shen, H. Epidemiological and genomic characteristics of global mcr-positive Escherichia coli isolates. Front. Microbiol. 2023, 13, 1105401. [Google Scholar] [CrossRef]
- Tansawai, U.; Yu, Y.; Kiddee, A.; Assawatheptawee, K.; Sands, K.; Hassan, B.; Walsh, T.R.; Niumsup, P.R. Emergence of mcr-3-mediated IncP and IncFII plasmids in Thailand. J. Glob. Antimicrob. Resist. 2021, 24, 446–447. [Google Scholar] [CrossRef]
- Rozwandowicz, M.; Brouwer, M.S.M.; Fischer, J.; Wagenaar, J.A.; Gonzalez-Zorn, B.; Guerra, B.; Mevius, D.J.; Hordijk, J. Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 1121–1137. [Google Scholar] [CrossRef]
- Boonyasiri, A.; Brinkac, L.M.; Jauneikaite, E.; White, R.C.; Greco, C.; Seenama, C.; Tangkoskul, T.; Nguyen, K.; Fouts, D.E.; Thamlikitkul, V. Characteristics and genomic epidemiology of colistin-resistant Enterobacterales from farmers, swine, and hospitalized patients in Thailand, 2014–2017. BMC Infect. Dis. 2023, 23, 556. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 33rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2023. [Google Scholar]
- Olson, R.D.; Assaf, R.; Brettin, T.; Conrad, N.; Cucinell, C.; Davis, J.J.; Dempsey, D.M.; Dickerman, A.; Dietrich, E.M.; Kenyon, R.W.; et al. Introducing the Bacterial and Viral Bioinformatics Resource Center (BV-BRC): A resource combining PATRIC, IRD and ViPR. Nucleic Acids Res. 2023, 51, D678–D689. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| AMR-0201 | AMR-0220 * | AMR-0251 | AMR-0354 | AMR-0361 * | AMR-0429 | ||
|---|---|---|---|---|---|---|---|
| Antimicrobial Class | Antimicrobial Agent | Minimum Inhibitory Concentration (μg/mL) | |||||
| Carbapenem | Ertapenem | ≤0.25 (S) | ≤0.25 (S) | ≤0.25 (S) | ≤0.25 (S) | ≤0.25 (S) | ≤0.25 (S) |
| Imipenem | ≤1 (S) | ≤1 (S) | ≤1 (S) | ≤1 (S) | ≤1 (S) | ≤1 (S) | |
| Meropenem | ≤1 (S) | ≤1 (S) | ≤1 (S) | ≤1 (S) | ≤1 (S) | ≤1 (S) | |
| Cephem | Cefepime 4 | >16 (R) | >16 (R) | >16 (R) | >16 (R) | 8 (R) | 16 (R) |
| Ceftazidime 3 | >16 (R) | >16 (R) | >16 (R) | >16 (R) | 8 (R) | 16 (R) | |
| Cephalothin 1 | >16 (R) | >16 (R) | >16 (R) | >16 (R) | >16 (R) | >16 (R) | |
| Ceftriaxone 3 | >16 (R) | >16 (R) | >16 (R) | >16 (R) | >16 (R) | >16 (R) | |
| Cefuroxime 2 | >16 (R) | >16 (R) | >16 (R) | >16 (R) | >16 (R) | >16 (R) | |
| Cefoxitin 2 | >16 (R) | 8 (S) | 8 (S) | 8 (S) | ≤4 (S) | ≤4 (S) | |
| β-lactam/β-lactam combination agent | Ampicillin | >16 (R) | >16 (R) | >16 (R) | >16 (R) | >16 (R) | >16 (R) |
| Amoxicillin-Clavulanate | >16/8 (R) | 8/4 (R) | ≤4/2 (R) | 8/4 (R) | >16/8 (R) | >16/8 (R) | |
| Piperacillin-Tazobactam | ≤4/4 (S) | ≤4/4 (S) | ≤4/4 (S) | ≤4/4 (S) | ≤4/4 (S) | ≤4/4 (S) | |
| 5-Fluoroquinolone | Ciprofloxacin | >2 (R) | >2 (R) | >2 (R) | ≤0.5 (S) | >2 (R) | 1 (S) |
| Norfloxacin | >8 (R) | >8 (R) | >8 (R) | ≤2 (S) | >8 (R) | ≤2 (S) | |
| Folate Antagonist | Trimethoprim | >8 (R) | ≤1 (S) | >8 (R) | ≤1 (S) | >8 (R) | >8 (R) |
| Trimethoprim-Sulfamethoxazole | >2/38 (R) | ≤0.5/9.5 (S) | >2/38 (R) | ≤0.5/9.5 (S) | >2/38 (R) | >2/38 (R) | |
| Aminoglycoside | Gentamicin | >8 (R) | >8 (R) | >8 (R) | ≤2 (S) | >8 (R) | >8 (R) |
| Tobramycin | 8 (I) | 8 (I) | 8 (I) | ≤2 (S) | 8 (I) | 8 (I) | |
| Nitrofurantoin Polymyxin | Nitrofurantoin | ≤16 (S) | ≤16 (S) | 32 (S) | ≤16 (S) | 64 (I) | 32 (S) |
| Colistin a | 4 (R) | 4 (R) | 4 (R) | 4 (R) | 4 (R) | 4 (R) | |
| Aminoglycoside | Beta-Lactam | Polymyxin | Phenicol | Quinolone | Lincosamide | Macrolide | Tetracycline | Trimethoprim | Sulfonamide | Rifampin | ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample ID | MLST | Replicon Type | aac(3)-IIa | aac(3)-IId | aadA1 | aadA2 | aadA22 | aph(3)-Ia | aph(3)-Ib | aph(6)-Id | blaCMY-2 | blaCTX-M-15 | blaCTX-M-55 | blaTEM-1 | mcr-1.1 | mcr-3.4 | mcr-3.5 | catA2 | cmlA1 | floR | qnrS1 | qnrS13 | lnu(F) | mdf(A) | mph(A) | erm(B) | tet(A) | tet(B) | tet(M) | dfrA12 | dfrA14 | dfrA17 | sul1 | sul2 | sul3 | arr-2 |
| AMR-0201 | 410 | IncFIB, IncI1, IncX4 | ||||||||||||||||||||||||||||||||||
| AMR-0220 * | 410 | IncFIB, IncI1, IncX4, IncY | ||||||||||||||||||||||||||||||||||
| AMR-0251 | 10 | IncHI2, IncY | ||||||||||||||||||||||||||||||||||
| AMR-0354 | 38 | IncFII, IncI1,IncI2 | ||||||||||||||||||||||||||||||||||
| AMR-0361 * | 46 | IncFIB, IncFII, IncN, IncX4 | ||||||||||||||||||||||||||||||||||
| AMR-0429 | 617 | incFII, IncI1, IncQ1, IncX1, IncFI, IncY | ||||||||||||||||||||||||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nobthai, P.; Ruekit, S.; Peerapongpaisarn, D.; Sukhchat, P.; Swierczewski, B.E.; Ruamsap, N.; Lertsethtakarn, P. The Co-Existence of mcr-1.1 and mcr-3.5 in Escherichia coli Isolated from Clinical Samples in Thailand. Antibiotics 2025, 14, 596. https://doi.org/10.3390/antibiotics14060596
Nobthai P, Ruekit S, Peerapongpaisarn D, Sukhchat P, Swierczewski BE, Ruamsap N, Lertsethtakarn P. The Co-Existence of mcr-1.1 and mcr-3.5 in Escherichia coli Isolated from Clinical Samples in Thailand. Antibiotics. 2025; 14(6):596. https://doi.org/10.3390/antibiotics14060596
Chicago/Turabian StyleNobthai, Panida, Sirigade Ruekit, Dutsadee Peerapongpaisarn, Prawet Sukhchat, Brett E. Swierczewski, Nattaya Ruamsap, and Paphavee Lertsethtakarn. 2025. "The Co-Existence of mcr-1.1 and mcr-3.5 in Escherichia coli Isolated from Clinical Samples in Thailand" Antibiotics 14, no. 6: 596. https://doi.org/10.3390/antibiotics14060596
APA StyleNobthai, P., Ruekit, S., Peerapongpaisarn, D., Sukhchat, P., Swierczewski, B. E., Ruamsap, N., & Lertsethtakarn, P. (2025). The Co-Existence of mcr-1.1 and mcr-3.5 in Escherichia coli Isolated from Clinical Samples in Thailand. Antibiotics, 14(6), 596. https://doi.org/10.3390/antibiotics14060596