
Mutation of smeRv Renders Stenotrophomonas maltophilia Resistant to First-Line Antibiotics Trimethoprim/Sulfamethoxazole and Levofloxacin

Abstract
1. Introduction
2. Results
2.1. Isolation of the Fluoroquinolone-Resistant Mutants
2.2. Mutation Analysis of S. maltophilia KE507
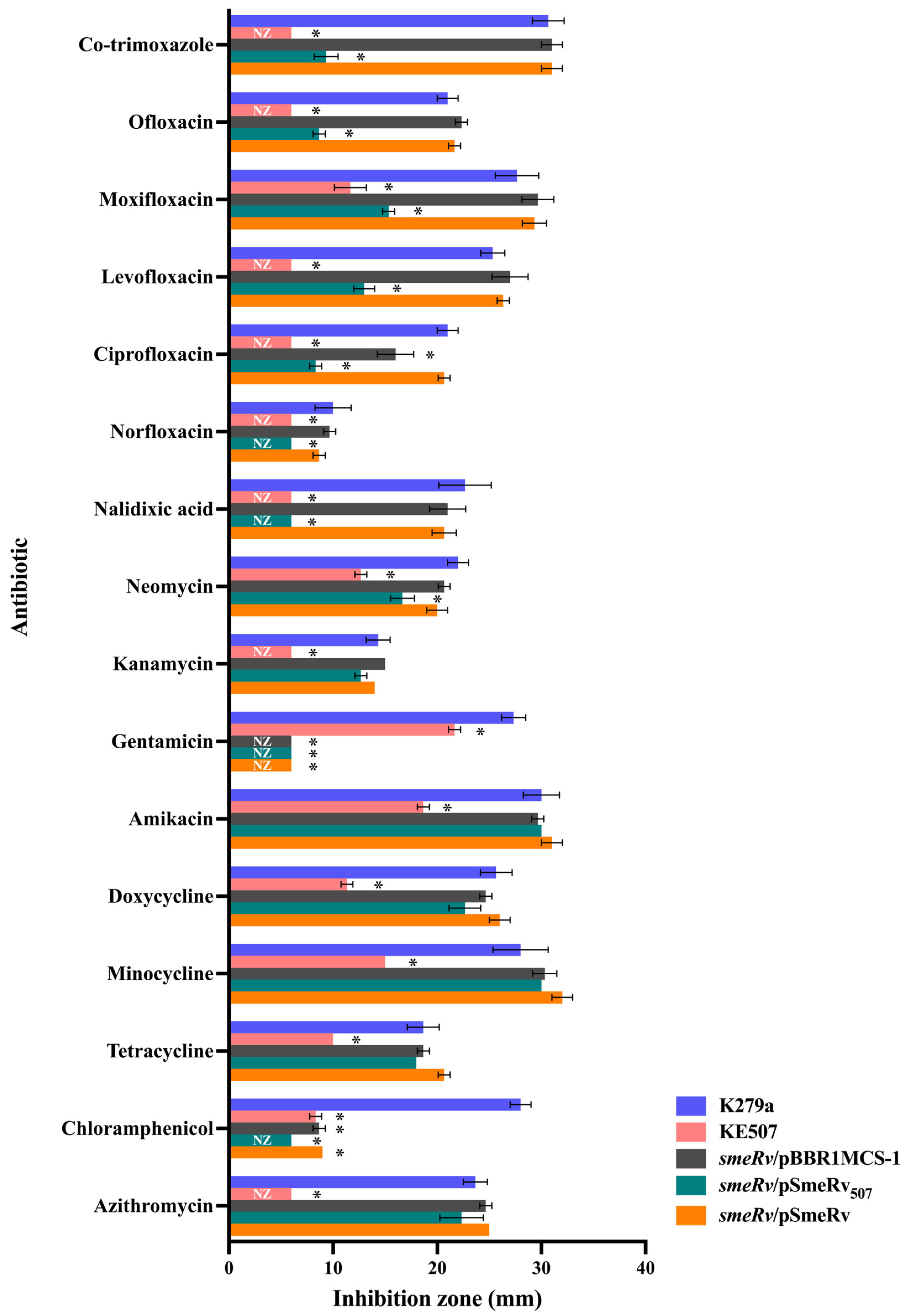
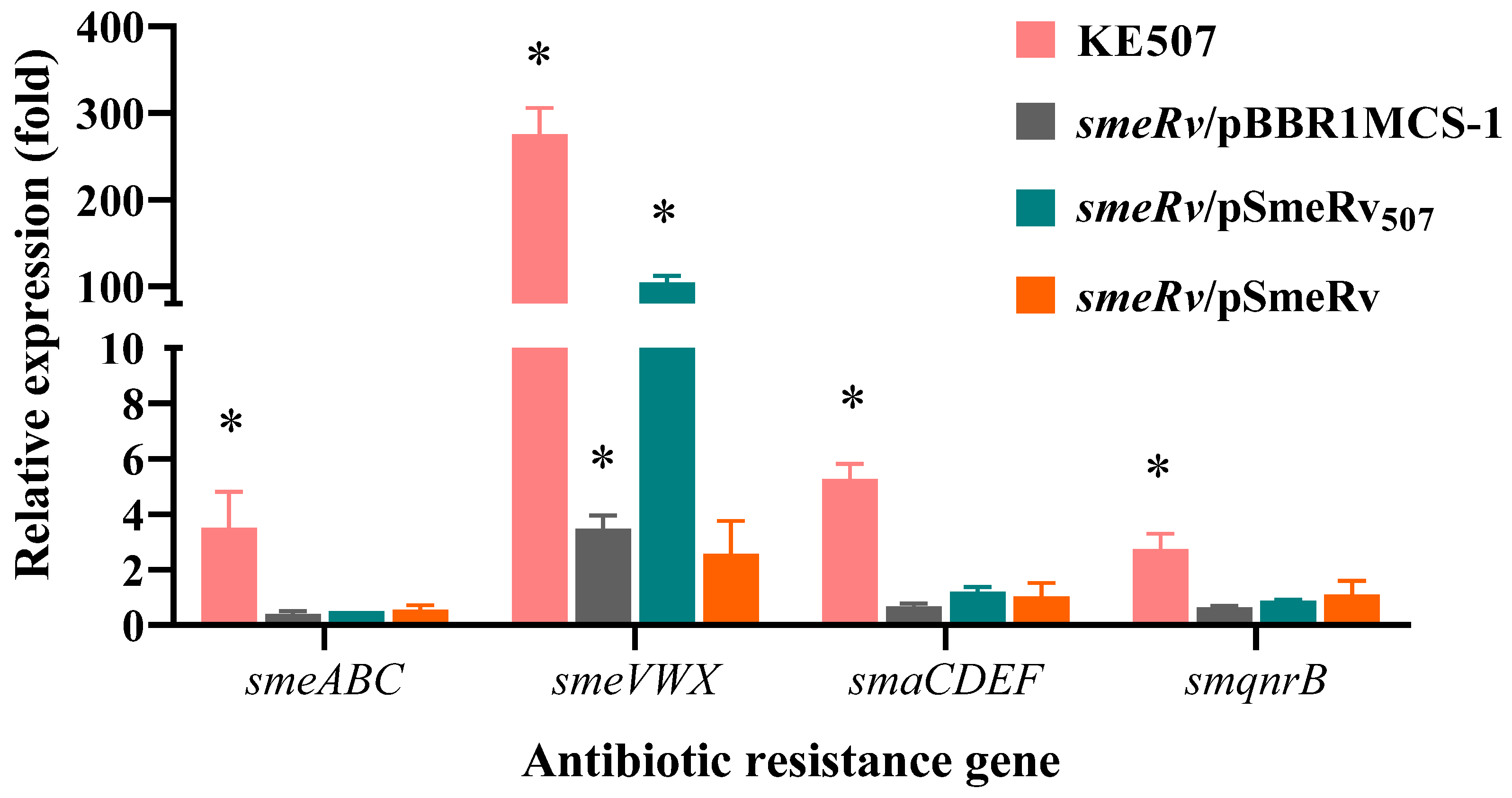
2.3. Characterization of the Mutations Responsible for the Antibiotic Resistance of KE507
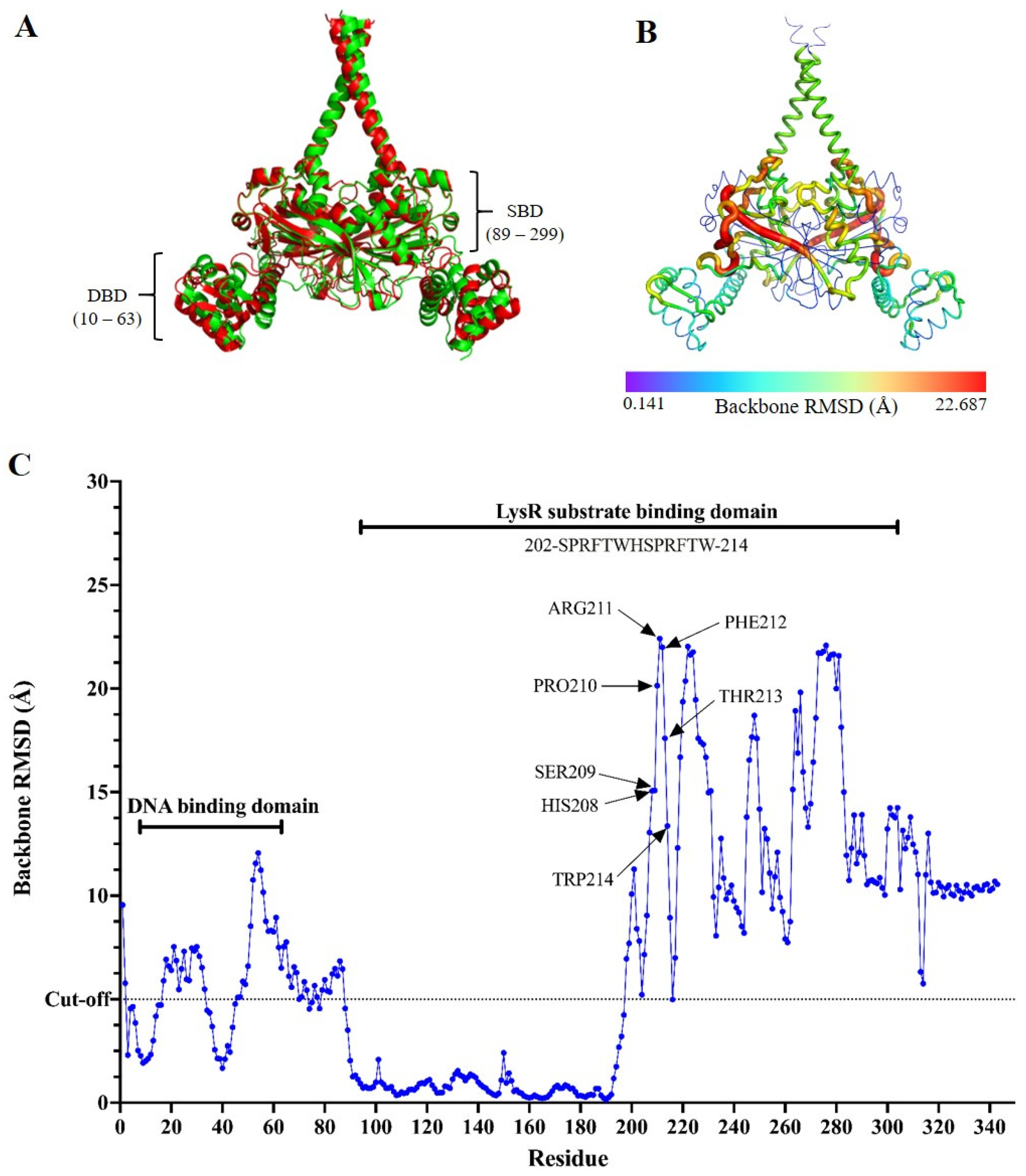
2.4. The Impact of the Insertion Mutation on the Protein Structure of SmeRv
3. Discussion
4. Materials and Methods
4.1. Bacterial Growth Conditions
4.2. Selection of KE507 Mutant
4.3. Molecular Biology Techniques
4.4. Antimicrobial Susceptibility Testing
4.5. Whole Genome Sequencing and Mutation Analysis
4.6. Real-Time Reverse Transcription PCR
4.7. Construction of the smeRv Mutant
4.8. Construction of pSmeRv and pSmeRv507
4.9. Prediction of SmeRv and SmeRv507 Protein Structures
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CLSI | Clinical and Laboratory Standards Institute |
| IMP | inner membrane |
| InDel | insertion/deletion |
| LTTRs | LysR-type transcriptional regulators |
| NZ | no inhibition zone |
| RMSD | root mean square deviation |
| RND | resistance-nodulation-cell division |
| RT-PCR | Reverse transcription polymerase chain reaction |
| SNP | Single nucleotide polymorphism |
References
- Brooke, J.S. Advances in the Microbiology of Stenotrophomonas maltophilia. Clin. Microbiol. Rev. 2021, 34, e0003019. [Google Scholar] [CrossRef] [PubMed]
- Al-Anazi, K.A.; Al-Jasser, A.M. Infections caused by Stenotrophomonas maltophilia in recipients of hematopoietic stem cell transplantation. Front. Oncol. 2014, 4, 232. [Google Scholar] [CrossRef] [PubMed]
- Adegoke, A.A.; Stenstrom, T.A.; Okoh, A.I. Stenotrophomonas maltophilia as an emerging ubiquitous pathogen: Looking beyond contemporary antibiotic therapy. Front. Microbiol. 2017, 8, 2276. [Google Scholar] [CrossRef]
- Hasbek, M.; Aldemir, O.; Cakir Kiymaz, Y.; Baysal, C.; Yildirim, D.; Buyuktuna, S.A. Mortality rates and risk factors associated with mortality in patients with Stenotrophomonas maltophilia primary bacteraemia and pneumonia. Diagn. Microbiol. Infect. Dis. 2024, 111, 116664. [Google Scholar] [CrossRef] [PubMed]
- Bhaumik, R.; Aungkur, N.Z.; Anderson, G.G. A guide to Stenotrophomonas maltophilia virulence capabilities, as we currently understand them. Front. Cell. Infect. Microbiol. 2023, 13, 1322853. [Google Scholar] [CrossRef]
- Mojica, M.F.; Humphries, R.; Lipuma, J.J.; Mathers, A.J.; Rao, G.G.; Shelburne, S.A.; Fouts, D.E.; Van Duin, D.; Bonomo, R.A. Clinical challenges treating Stenotrophomonas maltophilia infections: An update. JAC Antimicrob. Resist. 2022, 4, dlac040. [Google Scholar] [CrossRef]
- Crossman, L.C.; Gould, V.C.; Dow, J.M.; Vernikos, G.S.; Okazaki, A.; Sebaihia, M.; Saunders, D.; Arrowsmith, C.; Carver, T.; Peters, N.; et al. The complete genome, comparative and functional analysis of Stenotrophomonas maltophilia reveals an organism heavily shielded by drug resistance determinants. Genome Biol. 2008, 9, R74. [Google Scholar] [CrossRef]
- Chauviat, A.; Meyer, T.; Favre-Bonte, S. Versatility of Stenotrophomonas maltophilia: Ecological roles of RND efflux pumps. Heliyon 2023, 9, e14639. [Google Scholar] [CrossRef]
- Du, D.; Wang-Kan, X.; Neuberger, A.; van Veen, H.W.; Pos, K.M.; Piddock, L.J.V.; Luisi, B.F. Multidrug efflux pumps: Structure, function and regulation. Nat. Rev. Microbiol. 2018, 16, 523–539. [Google Scholar] [CrossRef]
- Chen, C.H.; Huang, C.C.; Chung, T.C.; Hu, R.M.; Huang, Y.W.; Yang, T.C. Contribution of resistance-nodulation-division efflux pump operon smeU1-V-W-U2-X to multidrug resistance of Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2011, 55, 5826–5833. [Google Scholar] [CrossRef]
- Sanchez, M.B.; Martinez, J.L. Overexpression of the efflux pumps SmeVWX and SmeDEF Is a major cause of resistance to co-trimoxazole in Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2018, 62, e00301-18. [Google Scholar] [CrossRef]
- Lu, H.F.; Tsai, Y.C.; Li, L.H.; Lin, Y.T.; Yang, T.C. Role of AzoR, a LysR-type transcriptional regulator, in SmeVWX pump-mediated antibiotic resistance in Stenotrophomonas maltophilia. J. Antimicrob. Chemother. 2021, 76, 2285–2293. [Google Scholar] [CrossRef] [PubMed]
- Blanco, P.; Corona, F.; Sanchez, M.B.; Martinez, J.L. Vitamin K(3) induces the expression of the Stenotrophomonas maltophilia SmeVWX multidrug efflux pump. Antimicrob. Agents Chemother. 2017, 61, e02453-16. [Google Scholar] [CrossRef]
- Wu, C.J.; Chiu, T.T.; Lin, Y.T.; Huang, Y.W.; Li, L.H.; Yang, T.C. Role of smeU1VWU2X operon in alleviation of oxidative stresses and occurrence of sulfamethoxazole-trimethoprim-resistant mutants in Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2018, 62, e02114-17. [Google Scholar] [CrossRef]
- Srijaruskul, K.; Charoenlap, N.; Namchaiw, P.; Chattrakarn, S.; Giengkam, S.; Mongkolsuk, S.; Vattanaviboon, P. Regulation by SoxR of mfsA, which encodes a major facilitator protein involved in paraquat resistance in Stenotrophomonas maltophilia. PLoS ONE 2015, 10, e0123699. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, M100, 34th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2024. [Google Scholar]
- Almangour, T.A.; Alkherb, Z.; Alruwaite, S.; Alsahli, R.; Alali, H.; Almohaizeie, A.; Almuhisen, S.; Alowais, S.A.; Saleh, K.B.; Fetyani, L.; et al. Trimethoprim-sulfamethoxazole versus levofloxacin for the treatment of Stenotrophomonas maltophilia infections: A multicentre cohort study. J. Glob. Antimicrob. Resist. 2024, 38, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Chatree, Y.; Charoenlap, N.; Vanitshavit, V.; Ruangrassamee, P.; Mongkolsuk, S.; Vattanaviboon, P. Induction of antimicrobial resistance of Stenotrophomonas maltophilia by exposure to nonlethal levels of antibiotics. Microb. Drug Resist. 2023, 29, 115–126. [Google Scholar] [CrossRef]
- Ochoa-Sanchez, L.E.; Martinez, J.L.; Gil-Gil, T. Evolution of resistance against ciprofloxacin, tobramycin, and trimethoprim/sulfamethoxazole in the environmental opportunistic pathogen Stenotrophomonas maltophilia. Antibiotics 2024, 13, 330. [Google Scholar] [CrossRef]
- Sanchez, P.; Moreno, E.; Martinez, J.L. The biocide triclosan selects Stenotrophomonas maltophilia mutants that overproduce the SmeDEF multidrug efflux pump. Antimicrob. Agents Chemother. 2005, 49, 781–782. [Google Scholar] [CrossRef]
- Blanco, P.; Hjort, K.; Martinez, J.L.; Andersson, D.I. Antimicrobial peptide exposure selects for resistant and fit Stenotrophomonas maltophilia mutants that show cross-resistance to antibiotics. mSphere 2020, 5, e00717-20. [Google Scholar] [CrossRef]
- Vanitshavit, V.; Charoenlap, N.; Sallabhan, R.; Whangsuk, W.; Bhinija, K.; Dulyayangkul, P.; Mongkolsuk, S.; Vattanaviboon, P. Acquired resistance of Stenotrophomonas maltophilia to antimicrobials induced by herbicide paraquat dichloride. PLoS ONE 2024, 19, e0309525. [Google Scholar] [CrossRef]
- Xi, F. The enrofloxacin pollution control from fish to environment. Mar. Pollut. Bull. 2024, 199, 115923. [Google Scholar] [CrossRef]
- Sanchez, M.B.; Hernandez, A.; Rodriguez-Martinez, J.M.; Martinez-Martinez, L.; Martinez, J.L. Predictive analysis of transmissible quinolone resistance indicates Stenotrophomonas maltophilia as a potential source of a novel family of Qnr determinants. BMC Microbiol. 2008, 8, 148. [Google Scholar] [CrossRef]
- Shimizu, K.; Kikuchi, K.; Sasaki, T.; Takahashi, N.; Ohtsuka, M.; Ono, Y.; Hiramatsu, K. Smqnr, a new chromosome-carried quinolone resistance gene in Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2008, 52, 3823–3825. [Google Scholar] [CrossRef]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J.; et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024, 630, 493–500. [Google Scholar] [CrossRef]
- Mojica, M.F.; Bonomo, R.A.; van Duin, D. Treatment approaches for severe Stenotrophomonas maltophilia infections. Curr. Opin. Infect. Dis. 2023, 36, 572–584. [Google Scholar] [CrossRef]
- Tamma, P.D.; Aitken, S.L.; Bonomo, R.A.; Mathers, A.J.; van Duin, D.; Clancy, C.J. Infectious Diseases Society of America Guidance on the treatment of AmpC β-Lactamase-producing Enterobacterales, carbapenem-resistant Acinetobacter baumannii, and Stenotrophomonas maltophilia infections. Clin. Infect. Dis. 2022, 74, 2089–2114. [Google Scholar] [CrossRef]
- Canton, R.; Ruiz-Garbajosa, P. Treatment guidelines for multidrug-resistant Gram-negative microorganisms. Rev. Esp. Quimioter. 2023, 36 (Suppl. 1), 46–51. [Google Scholar] [CrossRef]
- Scoffone, V.C.; Trespidi, G.; Barbieri, G.; Irudal, S.; Perrin, E.; Buroni, S. Role of RND efflux pumps in drug resistance of cystic fibrosis pathogens. Antibiotics 2021, 10, 863. [Google Scholar] [CrossRef]
- Baugh, A.C.; Momany, C.; Neidle, E.L. Versatility and complexity: Common and uncommon facets of LysR-type transcriptional regulators. Annu. Rev. Microbiol. 2023, 77, 317–339. [Google Scholar] [CrossRef]
- Kim, Y.; Chhor, G.; Tsai, C.S.; Winans, J.B.; Jedrzejczak, R.; Joachimiak, A.; Winans, S.C. Crystal structure of the ligand-binding domain of a LysR-type transcriptional regulator: Transcriptional activation via a rotary switch. Mol. Microbiol. 2018, 110, 550–561. [Google Scholar] [CrossRef]
- Zaim, J.; Kierzek, A.M. The structure of full-length LysR-type transcriptional regulators. Modeling of the full-length OxyR transcription factor dimer. Nucleic Acids Res. 2003, 31, 1444–1454. [Google Scholar] [CrossRef]
- Maddocks, S.E.; Oyston, P.C.F. Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins. Microbiology 2008, 154, 3609–3623. [Google Scholar] [CrossRef]
- Lerche, M.; Dian, C.; Round, A.; Lonneborg, R.; Brzezinski, P.; Leonard, G.A. The solution configurations of inactive and activated DntR have implications for the sliding dimer mechanism of LysR transcription factors. Sci. Rep. 2016, 6, 19988. [Google Scholar] [CrossRef]
- Dulyayangkul, P.; Calvopina, K.; Heesom, K.J.; Avison, M.B. Novel mechanisms of efflux-mediated levofloxacin resistance and reduced amikacin susceptibility in Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2020, 65, e01284-20. [Google Scholar] [CrossRef]
- Li, X.Z.; Zhang, L.; Poole, K. SmeC, an outer membrane multidrug efflux protein of Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2002, 46, 333–343. [Google Scholar] [CrossRef]
- Chang, L.L.; Chen, H.F.; Chang, C.Y.; Lee, T.M.; Wu, W.J. Contribution of integrons, and SmeABC and SmeDEF efflux pumps to multidrug resistance in clinical isolates of Stenotrophomonas maltophilia. J Antimicrob. Chemother. 2004, 53, 518–521. [Google Scholar] [CrossRef]
- Cho, H.H.; Sung, J.Y.; Kwon, K.C.; Koo, S.H. Expression of Sme efflux pumps and multilocus sequence typing in clinical isolates of Stenotrophomonas maltophilia. Ann. Lab. Med. 2012, 32, 38–43. [Google Scholar] [CrossRef]
- Bhagirath, A.Y.; Li, Y.; Patidar, R.; Yerex, K.; Ma, X.; Kumar, A.; Duan, K. Two component regulatory systems and antibiotic resistance in Gram-negative pathogens. Int. J. Mol. Sci. 2019, 20, 1781. [Google Scholar] [CrossRef]
- Dingemans, J.; Poudyal, B.; Sondermann, H.; Sauer, K. The yin and yang of SagS: Distinct residues in the HmsP domain of SagS independently regulate biofilm formation and biofilm drug tolerance. mSphere 2018, 3, e00192-18. [Google Scholar] [CrossRef]
- Chen, S.K.; Guan, H.H.; Wu, P.H.; Lin, L.T.; Wu, M.C.; Chang, H.Y.; Chen, N.C.; Lin, C.C.; Chuankhayan, P.; Huang, Y.C.; et al. Structural insights into the histidine-containing phospho-transfer protein and receiver domain of sensor histidine kinase suggest a complex model in the two-component regulatory system in Pseudomonas aeruginosa. IUCrJ 2020, 7, 934–948. [Google Scholar] [CrossRef]
- Gupta, K.; Marques, C.N.; Petrova, O.E.; Sauer, K. Antimicrobial tolerance of Pseudomonas aeruginosa biofilms is activated during an early developmental stage and requires the two-component hybrid SagS. J. Bacteriol. 2013, 195, 4975–4987. [Google Scholar] [CrossRef]
- Tang, L.; Dong, S.; Rasheed, N.; Wu, H.W.; Zhou, N.; Li, H.; Wang, M.; Zheng, J.; He, J.; Chao, W.C.H. Vibrio parahaemolyticus prey targeting requires autoproteolysis-triggered dimerization of the type VI secretion system effector RhsP. Cell Rep. 2022, 41, 111732. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Wood, G.L.; Washington, J.A. Antibacterial Susceptibility Tests: Dilution and Disk Diffusion Methods. In Manual of Clinical Microbiology; Murray, P.R., Ed.; ASM Press: Washington DC, USA, 1995; pp. 1327–1341. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2012. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 8 November 2024).
- Wang, K.; Li, M.; Hakonarson, H. ANNOVAR: Functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010, 38, e164. [Google Scholar] [CrossRef]
- Alexeyev, M.F. The pKNOCK series of broad-host-range mobilizable suicide vectors for gene knockout and targeted DNA insertion into the chromosome of gram-negative bacteria. Biotechniques 1999, 26, 824–826, 828. [Google Scholar] [CrossRef]
- Kovach, M.E.; Phillips, R.W.; Elzer, P.H.; Roop, R.M., 2nd; Peterson, K.M. pBBR1MCS: A broad-host-range cloning vector. Biotechniques 1994, 16, 800–802. [Google Scholar]
- Wee, J.; Wei, G.W. Evaluation of AlphaFold 3’s protein-protein complexes for predicting binding free energy changes upon mutation. J. Chem. Inf. Model. 2024, 64, 6676–6683. [Google Scholar] [CrossRef]
- Mulder, N.J.; Kersey, P.; Pruess, M.; Apweiler, R. In silico characterization of proteins: UniProt, InterPro and Integr8. Mol. Biotechnol. 2008, 38, 165–177. [Google Scholar] [CrossRef]
- Reva, B.A.; Finkelstein, A.V.; Skolnick, J. What is the probability of a chance prediction of a protein structure with an rmsd of 6 Å. Fold Des. 1998, 3, 141–147. [Google Scholar] [CrossRef]
- Pathak, Y.; Singh, P.K.; Saraswat, M. Protein structure prediction RMSD ≤ 5 Å using machine learning models. Int. J. Data Min. Bioinform. 2016, 14, 71–85. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus Tag/Position | Gene Product | Type | Effect | Amino Acid Change |
|---|---|---|---|---|
| smlt0042 | Putative rearrangement hotspot (RHS)-repeat protein | SNV | G>C | R631P |
| smlt0570 | Putative sensor histidine kinase/response regulator fusion protein | INS | Frameshift insertion >CGCCGCCATGGCGGCCTCGGCCTGGGCCTGGCGATCGTGCAGCAGCTGGTCGAACTGCACGGCGGCACCGTCGC | Q459fs |
| INS | In-frame insertion >GGCATGCCG | H565insAMH | ||
| smlt1827 | SmeRv (LysR-type transcriptional regulator) | INS | In-frame insertion >TGCCAGGTGAAGCGCGGCGAA | Q208ins- HSPRFTW |
| smlt3968 | Conserved hypothetical protein | SNV | C>G | P56P |
| Position 2935035 | SNV | C>A Intergenic region between smlt2891 (putative two-component system response regulator transcriptional regulatory protein) and smlt2892 (putative Major Facilitator Superfamily transporter | - |
| Gene | Locus Tag | Product | Antibiotic Resistance | Relative Expression (Fold) |
|---|---|---|---|---|
| smeABC | smlt4474-6 | RND-type tripartite efflux protein | β-lactams, Aminoglycosides, Quinolones | 3.5 ± 1.4 * |
| smeDEF | smlt4070-2 | RND-type tripartite efflux protein | Fluoroquinolones, Macrolides, Tetracycline, Chloramphenicol, Cotrimoxazole | 0.7 ± 0.1 |
| smeGH | smlt3170-1 | RND-type tripartite efflux protein | Quinolones, Macrolides, Tetracycline, Chloramphenicol | 0.8 ± 0.2 |
| smeIJK | smlt4279-81 | RND-type tripartite efflux proteins | Aminoglycosides, Fluoroquinolones, Tetracycline | 1 ± 0.1 |
| smeOP | smlt3925-4 | RND-type tripartite efflux transporter | Quinolones, Aminoglycosides, Macrolides, β-lactams, Doxycycline | 1 ± 0.1 |
| smeVWX | smlt1830-3 | RND-type tripartite efflux transporter | Quinolones, Cotrimoxazole, Tetracycline, Chloramphenicol | 275.7 ± 30.2 * |
| smeYZ | smlt2201-2 | RND-type efflux protein and MFP | Aminoglycosides, Cotrimoxazole | 0.8 ± 0.4 |
| mdtD | smlt3623 | MFS-type transporter protein | Macrolides, Tetracycline | 2.2 ± 0.4 * |
| mfsA | smlt1083 | MFS-type transporter protein | Fluoroquinolones; Aminoglycosides, Cephalosporins | 1.2 ± 0.4 |
| smrA | smlt1471 | ABC-type efflux pump | Quinolones, Tetracycline | 1.2 ± 0.3 |
| tcrA | smlt1069 | MFS-type transporter protein | Tetracycline | 1 ± 0 |
| macABC | smlt1539 | ABC-type efflux pump | Macrolides, Aminoglycosides | 1.2 ± 0.4 |
| smaAB | smlt2642-3 | ABC-type efflux pump | Aminoglycosides | 2 ± 0.8 |
| smaCDEF | smlt1651-4 | ABC-type efflux pump | Levofloxacin | 5.3 ± 0.6 * |
| pmpM | smlt1381 | MATE-type efflux protein | Fluoroquinolones | 1.3 ± 1.2 |
| blaL1 | smlt2667 | β-lactamase-L1 | β-lactams | 1.1 ± 0.2 |
| blaL2 | smlt3722 | β-lactamase-L2 | β-lactams | 0.8 ± 0.2 |
| aph(3′)IIC | smlt2120 | Aminoglycoside phosphotransferase | Aminoglycosides | 2.6 ± 0.8 |
| aac(6′)-Iz | smlt3615 | Aminoglycoside 6′-N-acetyltransferase | Aminoglycosides | 0.4 ± 0 |
| smqnrB | smlt1071 | Quinolone resistance protein | Quinolones | 2.7 ± 0.6 * |
| Primer | Sequence (5′→3′) | Gene | Product 1 |
|---|---|---|---|
| BT2781 | GCCCGCACAAGCGGTGGAG | 16S rRNA forward primer | rRNA |
| BT2782 | ACGTCATCCCCACCTTCC | 16S rRNA reverse primer | rRNA |
| BT8127 | CCCGCATCAACCTCGACTAC | smeABC forward primer | RND |
| BT8128 | CAGCACCTTTACCTGTGCCT | smeABC reverse primer | RND |
| BT8129 | CAACGTCACCCTCGGCTATG | smeDEF forward primer | RND |
| BT8130 | CGACGCTCACTTCAGAGAACT | smeDEF reverse primer | RND |
| BT8133 | CCGATCCACGTCCTGTTCAA | smeIJK forward primer | RND |
| BT8134 | GTAGACGTACTCGCCATCCG | smeIJK reverse primer | RND |
| BT8137 | GAACTGGACGTGGCTGACTTC | smeOP forward primer | RND |
| BT8138 | CAGGTCGATCAGCACTTCGC | smeOP reverse primer | RND |
| BT8139 | GGAGTTCACCAAGGTGCGT | smeVWX forward primer | RND |
| BT8140 | GAAGTCGACCTTGCCCGAAT | smeVWX reverse primer | RND |
| BT8141 | CCAGTGCCGAGTACGAACAG | smeYZ forward primer | RND |
| BT8142 | CGCGCATCGACATTGATACC | smeYZ reverse primer | RND |
| BT8149 | ATCGGCGTGATCGGATTCAT | mdtD forward primer | MFS |
| BT8150 | CCGATGCGCGAGAAAAGATT | mdtD reverse primer | MFS |
| BT7111 | TCTGGTACGGATTGGCCTGC | mfsA forward primer | MFS |
| BT7112 | CCGACCATCGAAGGCACCAC | mfsA reverse primer | MFS |
| BT8255 | CGGGTGATGATGTCTGGCTT | tcrA forward primer | MFS |
| BT8256 | TGAAGGTCACATAGCCGACG | tcrA reverse primer | MFS |
| BT8189 | GATCCAGGAACTGGCGGTC | smrA forward primer | ABC |
| BT8190 | CTGGGCGATGGTGGTGATG | smrA reverse primer | ABC |
| BT8193 | CCGAAGCACAGCTGAAAACTG | macABC forward primer | ABC |
| BT8194 | GGTACTTGCGGTCGGGGTC | macABC reverse primer | ABC |
| BT8197 | TTGCCGAAGTGGATTCGCAG | smaAB forward primer | ABC |
| BT8198 | GTGAGACGATGCGGGTGTAG | smaAB reverse primer | ABC |
| BT9036 | TCCGATTCCAGTCCCTCGAT | smaCDEF forward primer | ABC |
| BT9037 | CGTATCCAGCCCATCGAACT | smaCDEF reverse primer | ABC |
| BT8199 | TATGCGTTCGCCTTCCTCAC | pmpM forward primer | MATE |
| BT8200 | GCACCAGCGCTTTCAGGATG | pmpM reverse primer | MATE |
| BT8213 | GGTCACCTGCTGGACAACAT | blaL1 forward primer | ENZ |
| BT8214 | CACTTCGCCGTCCATGATGA | blaL1 reverse primer | ENZ |
| BT8215 | GGCATTGCTGGACAGGCG | blaL2 forward primer | ENZ |
| BT8216 | GCCCTTGGCAAAGCTGTTCA | blaL2 reverse primer | ENZ |
| BT8225 | TAATTGCCACCGCCGAAGAA | aph(3′)IIc forward primer | ENZ |
| BT8226 | AGTCATCGGCATCCACCAACC | aph(3′)IIc reverse primer | ENZ |
| BT8231 | GACGGTTGGTTTCGCTGAAG | aac(6′)Iz forward primer | ENZ |
| BT8232 | GCGGAAATAGACGACCCGTT | aac(6′)Iz reverse primer | ENZ |
| BT6866 | TCAATGGCGCCACGCTGAAG | smqnrB forward primer | ARP |
| BT6867 | TCCAGCGTTACCCGCGAGAA | smqnrB reverse primer | ARP |
| BT8566 | ATGCACATGGGCAACGAGAA | smeRv fragment forward primer | smeRv fragment |
| BT8567 | GCACCATTCAACGCAGACAT | smeRv fragment reverse primer | smeRv fragment |
| BT9444 | CCAGGATCCCCGACCAT | smeRv full-length forward primer | SmeRv |
| BT9445 | CGCTCTTGTTGCAGGCTA | smeRv full-length reverse primer | SmeRv |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boonyong, N.; Charoenlap, N.; Tipanyo, P.; Grittanaanun, P.; Mongkolsuk, S.; Vattanaviboon, P. Mutation of smeRv Renders Stenotrophomonas maltophilia Resistant to First-Line Antibiotics Trimethoprim/Sulfamethoxazole and Levofloxacin. Antibiotics 2025, 14, 550. https://doi.org/10.3390/antibiotics14060550
Boonyong N, Charoenlap N, Tipanyo P, Grittanaanun P, Mongkolsuk S, Vattanaviboon P. Mutation of smeRv Renders Stenotrophomonas maltophilia Resistant to First-Line Antibiotics Trimethoprim/Sulfamethoxazole and Levofloxacin. Antibiotics. 2025; 14(6):550. https://doi.org/10.3390/antibiotics14060550
Chicago/Turabian StyleBoonyong, Nuchjaree, Nisanart Charoenlap, Parinya Tipanyo, Pitthawat Grittanaanun, Skorn Mongkolsuk, and Paiboon Vattanaviboon. 2025. "Mutation of smeRv Renders Stenotrophomonas maltophilia Resistant to First-Line Antibiotics Trimethoprim/Sulfamethoxazole and Levofloxacin" Antibiotics 14, no. 6: 550. https://doi.org/10.3390/antibiotics14060550
APA StyleBoonyong, N., Charoenlap, N., Tipanyo, P., Grittanaanun, P., Mongkolsuk, S., & Vattanaviboon, P. (2025). Mutation of smeRv Renders Stenotrophomonas maltophilia Resistant to First-Line Antibiotics Trimethoprim/Sulfamethoxazole and Levofloxacin. Antibiotics, 14(6), 550. https://doi.org/10.3390/antibiotics14060550