
Persistence of Colistin Resistance and mcr-1.1-Positive E. coli in Poultry Despite Colistin Ban in Japan


Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Evaluation of Bacteria
2.2.1. Isolation and Identification of E. coli Strains
2.2.2. Antimicrobial Susceptibility
2.2.3. PCR Testing of ESBL Genes
2.3. Molecular Biology Techniques
2.3.1. DNA Extraction
2.3.2. NGS Analysis
2.3.3. Genome Informatics Analysis
3. Results
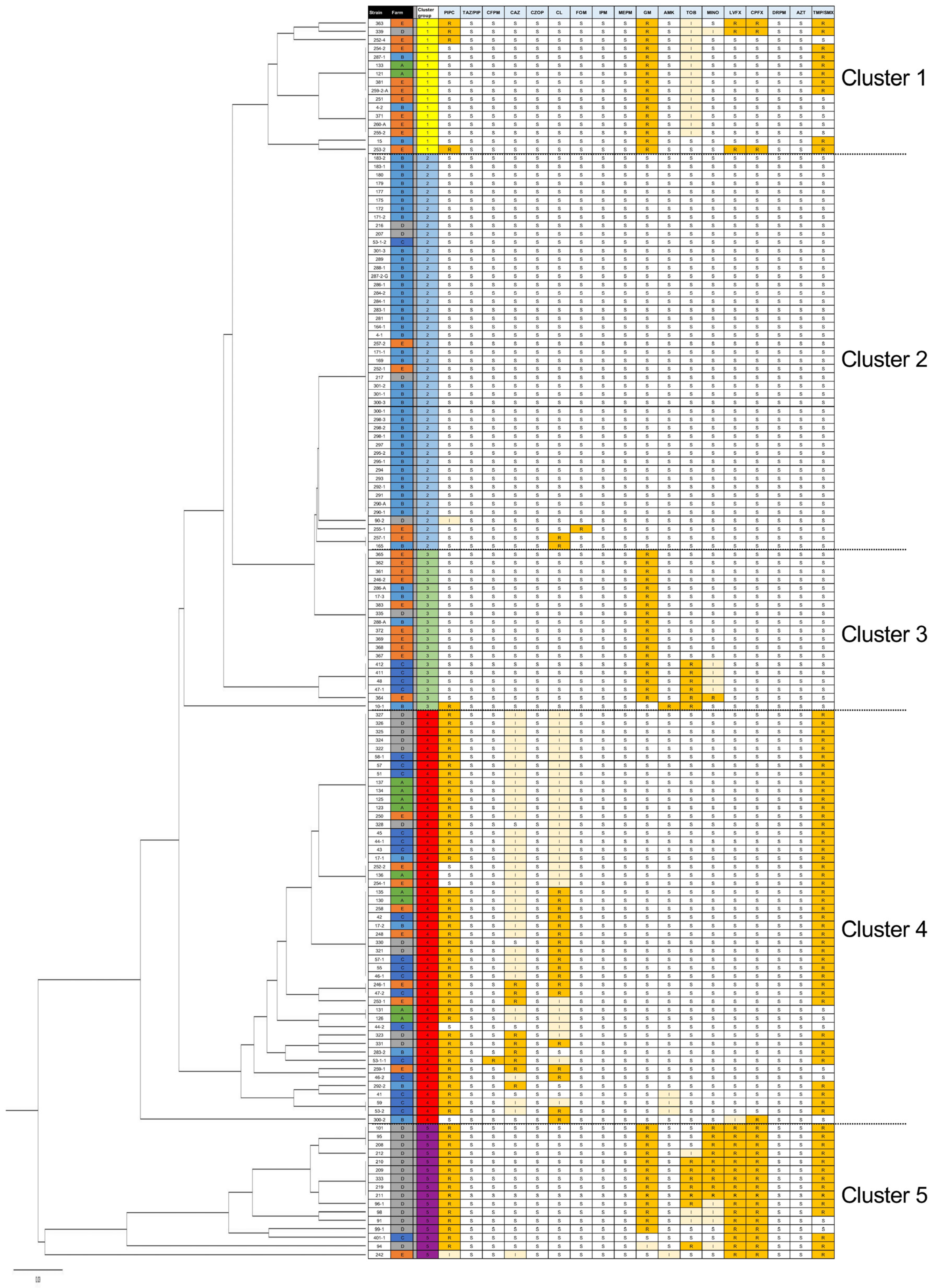
3.1. Antimicrobial Susceptibility of the E. coli Isolates Obtained from Broiler Feces
3.2. Whole-Genome Sequencing of Antimicrobial-Resistant E. coli Strains
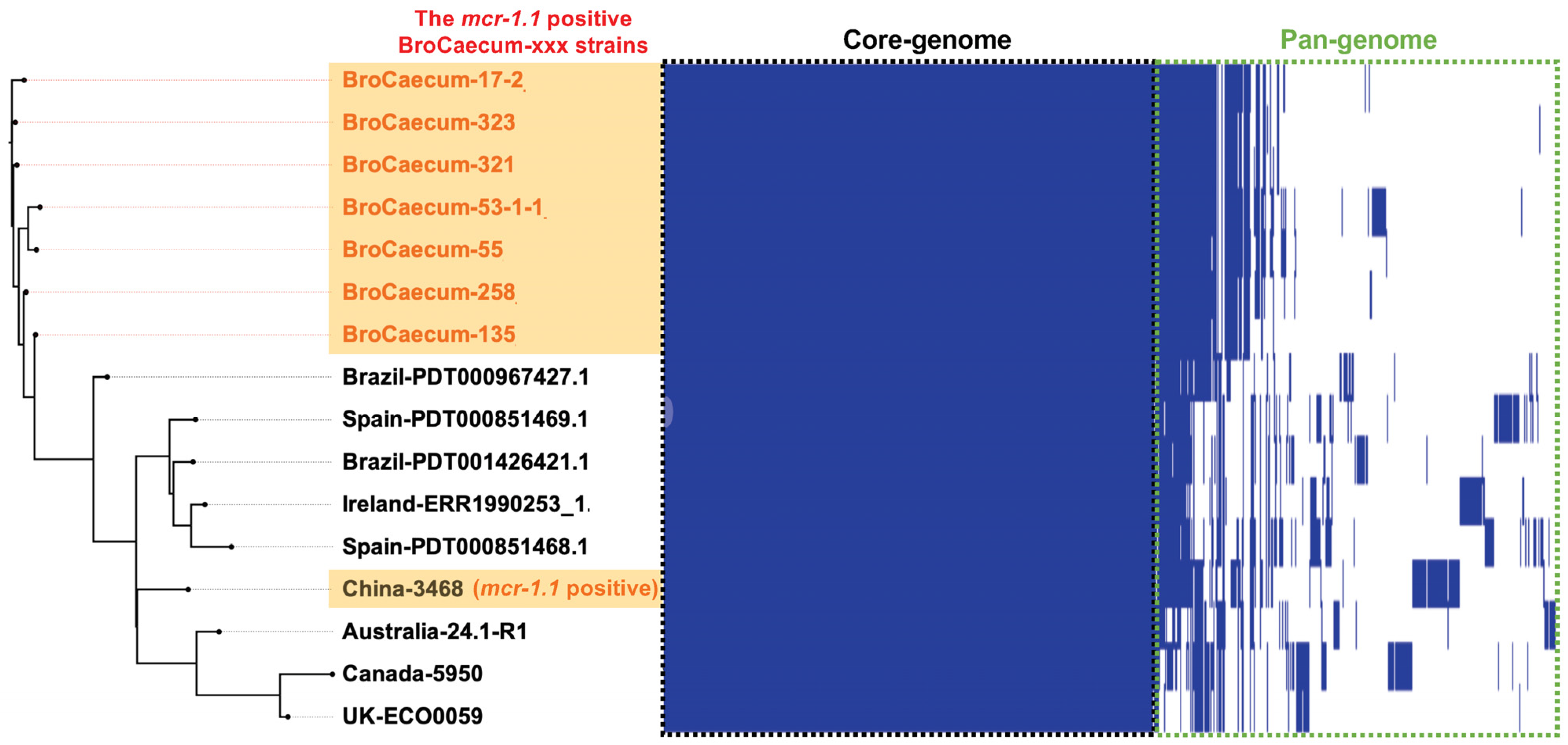
3.3. Comparative Genome Analysis of E. coli ST1485 Strains
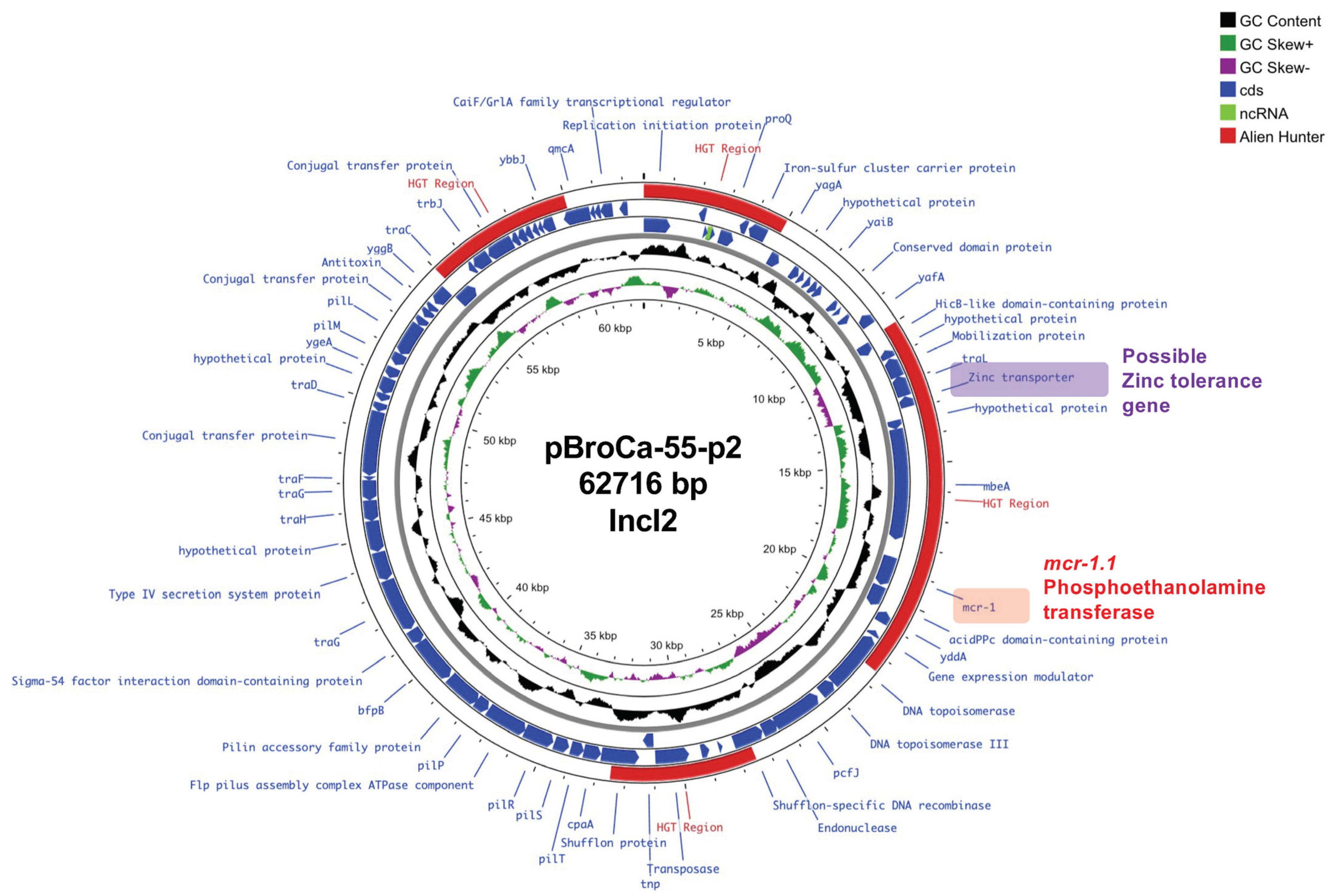
3.4. Complete Genome Sequence of the mcr-1.1-Positive E. coli BroCaecum-55 Strain
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MIC | Minimum inhibitory concentration |
| CLSI | Clinical and Laboratory Standards Institute |
| S | Susceptible |
| I | Intermediate |
| R | Resistant |
| ESBL | Extended-spectrum β-lactamase |
| PIPC | Piperacillin |
| TAZ/PIPC | Tazobactam/piperacillin |
| CFPM | Cefepime |
| CAZ | Ceftazidime |
| CZOP | Cefozopran |
| CL | Colistin |
| FOM | Fosfomycin |
| IPM | Imipenem |
| MEPM | Meropenem |
| GM | Gentamicin |
| AMK | Amikacin |
| TOB | Tobramycin |
| MINO | Minocycline |
| LVFX | Levofloxacin |
| CPFX | Ciprofloxacin |
| DRPM | Doripenem |
| AZT | Aztreonam |
| TMP/SMX | Trimethoprim/sulfamethoxazole |
| MLST | Multilocus sequence typing |
| ST | Sequence type |
| AMR | Antimicrobial resistance |
| ARGs | Antimicrobial resistant genes |
| JVARM | Japanese Veterinary Antimicrobial Resistance Monitoring System |
| JANIS | Japan Nosocomial Infections Surveillance |
| UPGMA | Unweighted pair group method with arithmetic mean |
| SNPs | Single nucleotide polymorphisms |
| PLSDB | Plasmid Sequence Database |
| NGS | Next-generation sequencing |
| DDBJ | DNA Data Bank of Japan |
References
- Antimicrobial Resistance (AMR) One Health Platform System for Animals Antimicrobial. Available online: https://amr-onehealth-platform.ncgm.go.jp/resistantBacteria/202 (accessed on 20 January 2025).
- World Health Organization. Global Action Plan on Antimicrobial Resistance. 2015. Available online: https://www.who.int/publications/i/item/9789241509763 (accessed on 20 January 2025).
- National Action Plan on Antimicrobial Resistance (AMR) (2023–2027) Japan. Available online: https://www.kantei.go.jp/jp/singi/kokusai_kansen/pdf/action_plan.pdf (accessed on 20 January 2025).
- Dutil, L.; Irwin, R.; Finley, R.; Ng, L.K.; Avery, B.; Boerlin, P.; Bourgault, A.M.; Cole, L.; Daignault, D.; Desruisseau, A.; et al. Ceftiofur resistance in Salmonella enterica serovar Heidelberg from chicken meat and humans, Canada. Emerg. Infect. Dis. 2010, 16, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Food Health Effects Assessment of Drug-Resistant Bacteria Related to Colistin Sulfate for Use in Livestock. Food Safety Commission Working Group on Drug-Resistant Bacteria. 2016. Available online: https://www.fsc.go.jp/senmon/sonota/amr_wg/amr_info.data/161122_colistin_draft_report.pdf (accessed on 20 January 2025).
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Martiny, H.M.; Munk, P.; Brinch, C.; Szarvas, J.; Aarestrup, F.M.; Petersen, T.N. Global Distribution of mcr Gene Variants in 214K Metagenomic Samples. mSystems 2022, 7, e0010522. [Google Scholar] [CrossRef] [PubMed]
- WHO. Critically Important Antimicrobials for Human Medicine: 6th Revision. Available online: https://www.who.int/publications/i/item/9789241515528 (accessed on 20 January 2025).
- Suzuki, S.; Ohnishi, M.; Kawanishi, M.; Akiba, M.; Kuroda, M. Investigation of a plasmid genome database for colistin-resistance gene mcr-1. Lancet Infect. Dis. 2016, 16, 284–285. [Google Scholar] [CrossRef]
- Kawanishi, M.; Abo, H.; Ozawa, M.; Uchiyama, M.; Shirakawa, T.; Suzuki, S.; Shima, A.; Yamashita, A.; Sekizuka, T.; Kato, K.; et al. Prevalence of Colistin Resistance Gene mcr-1 and Absence of mcr-2 in Escherichia coli Isolated from Healthy Food-Producing Animals in Japan. Antimicrob. Agents Chemother. 2017, 61, e02057-16. [Google Scholar] [CrossRef]
- Kusumoto, M.; Ogura, Y.; Gotoh, Y.; Iwata, T.; Hayashi, T.; Akiba, M. Colistin-Resistant mcr-1-Positive Pathogenic Escherichia coli in Swine, Japan, 2007–2014. Emerg. Infect. Dis. 2016, 22, 1315–1317. [Google Scholar] [CrossRef]
- Fukuda, A.; Sato, T.; Shinagawa, M.; Takahashi, S.; Asai, T.; Yokota, S.I.; Usui, M.; Tamura, Y. High prevalence of mcr-1, mcr-3 and mcr-5 in Escherichia coli derived from diseased pigs in Japan. Int. J. Antimicrob. Agents 2018, 51, 163–164. [Google Scholar] [CrossRef]
- Tada, T.; Uechi, K.; Nakasone, I.; Shimada, K.; Nakamatsu, M.; Kirikae, T.; Fujita, J. Emergence of a colistin-resistant Escherichia coli clinical isolate harboring mcr-1 in Japan. Int. J. Infect. Dis. 2017, 63, 21–22. [Google Scholar] [CrossRef]
- Tada, T.; Uechi, K.; Nakasone, I.; Nakamatsu, M.; Satou, K.; Hirano, T.; Kirikae, T.; Fujita, J. Emergence of IncX4 plasmids encoding mcr-1 in a clinical isolate of Klebsiella pneumoniae in Japan. Int. J. Infect. Dis. 2018, 75, 98–100. [Google Scholar] [CrossRef]
- Revocation of Designation of Feed Additive “Colistin Sulfate” in Ministry of Agriculture, Forestry and Fisheries Japan. Available online: https://www.maff.go.jp/j/syouan/tikusui/siryo/attach/pdf/additive-13.pdf (accessed on 20 January 2025).
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 34th Edition. Available online: https://clsi.org/standards/products/microbiology/documents/m100/ (accessed on 20 January 2025).
- Tanizawa, Y.; Fujisawa, T.; Kaminuma, E.; Nakamura, Y.; Arita, M. DFAST and DAGA: Web-based integrated genome annotation tools and resources. Biosci. Microbiota Food Health 2016, 35, 173–184. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Treangen, T.J.; Ondov, B.D.; Koren, S.; Phillippy, A.M. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014, 15, 524. [Google Scholar] [CrossRef]
- Grant, J.R.; Enns, E.; Marinier, E.; Mandal, A.; Herman, E.K.; Chen, C.Y.; Graham, M.; Van Domselaar, G.; Stothard, P. Proksee: In-depth characterization and visualization of bacterial genomes. Nucleic Acids Res. 2023, 51, W484–W492. [Google Scholar] [CrossRef]
- Molano, L.G.; Hirsch, P.; Hannig, M.; Muller, R.; Keller, A. The PLSDB 2025 update: Enhanced annotations and improved functionality for comprehensive plasmid research. Nucleic Acids Res 2025, 53, D189–D196. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D.; Nakagawa, S. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar]
- Antimicrobial Resistance (AMR) One Health Platform System for Animals Antimicrobial-Resistance. Available online: https://amr-onehealth-platform.ncgm.go.jp/resistantBacteria/201 (accessed on 20 January 2025).
- Yoshizawa, N.; Hikoda-Kogiku, Y.; Tamamura-Andoh, Y.; Kusumoto, M. mcr-1 remains detectable in various Escherichia coli lineages isolated from healthy swine after withdrawal of colistin use on the farm. J. Vet. Med. Sci. 2023, 85, 536–540. [Google Scholar] [CrossRef]
- Shen, C.; Zhong, L.L.; Zhong, Z.; Doi, Y.; Shen, J.; Wang, Y.; Ma, F.; Ahmed, M.; Zhang, G.; Xia, Y.; et al. Prevalence of mcr-1 in Colonized Inpatients, China, 2011–2019. Emerg. Infect. Dis. 2021, 27, 2502–2504. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, C.; Zhang, R.; Chen, Y.; Shen, Y.; Hu, F.; Liu, D.; Lu, J.; Guo, Y.; Xia, X.; et al. Changes in colistin resistance and mcr-1 abundance in Escherichia coli of animal and human origins following the ban of colistin-positive additives in China: An epidemiological comparative study. Lancet Infect. Dis. 2020, 20, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, T.; Wang, C.; Liang, G.; Lu, Q.; Wen, G.; Guo, Y.; Cheng, Y.; Wang, Z.; Shao, H.; et al. Prevalence of colistin resistance gene mcr-1 in Escherichia coli isolated from chickens in central China, 2014 to 2019. J. Glob. Antimicrob. Resist. 2022, 29, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Japanese Association of Swine Practicing Veterinarians (JASV). Collection of Case Studies on Efforts to De-antimicrobial Feed Additives. 2017. Available online: https://www.e-jasv.com/alic.pdf (accessed on 8 January 2025).
- Arai, N.; Shibahara, T.; Nishiura, R.; Tamamura-Andoh, Y.; Nishiura, H.; Muneta, Y.; Sawada, H.; Watanabe-Yanai, A.; Iwata, T.; Akiba, M.; et al. ICEmST contributes to colonization of Salmonella in the intestine of piglets. Sci. Rep. 2024, 14, 31407. [Google Scholar] [CrossRef]
- Shigemura, H.; Matsui, M.; Sekizuka, T.; Onozuka, D.; Noda, T.; Yamashita, A.; Kuroda, M.; Suzuki, S.; Kimura, H.; Fujimoto, S.; et al. Decrease in the prevalence of extended-spectrum cephalosporin-resistant Salmonella following cessation of ceftiofur use by the Japanese poultry industry. Int. J. Food Microbiol. 2018, 274, 45–51. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Farm | aph(6)-Id | aph(3″)-Ib | blaTEM | blaCMY-2 | mcr-1 | qnrS1 | tet(A) | sul2 | dfrA14 |
|---|---|---|---|---|---|---|---|---|---|---|
| BroCaecum-17-2 | B | + | + | + | + | + | + | + | + | + |
| BroCaecum-53-1-1 | C | + | + | + | + | + | + | – | + | + |
| BroCaecum-55 | C | + | + | + | + | + | + | – | + | + |
| BroCaecum-135 | A | + | + | + | + | + | + | – | + | + |
| BroCaecum-258 | E | + | + | + | + | + | + | – | + | + |
| BroCaecum-321 | D | + | + | + | + | + | + | + | + | + |
| BroCaecum-323 | D | + | + | + | + | + | + | – | + | + |
| Chromosome | pBroCa-55-p1 | pBroCa-55-p2 | pBroCa-55-p3 | pBroCa-55-p4 | |
|---|---|---|---|---|---|
| Total length (bp) | 5,098,687 | 176,133 | 62,716 | 5875 | 3373 |
| Status | circular | circular | circular | circular | circular |
| Inc replicon | – | IncFIB | IncI2 | rep_cluster_2401 | – |
| Copy number | 1.0 | 1.3 | 2.2 | 4.4 | 2.5 |
| GC content (%) | 50.6% | 50.2% | 42.8% | 47.5% | 55.2% |
| No. of CDSs | 4648 | 181 | 74 | 7 | 4 |
| No. of rRNA | 22 | – | – | – | – |
| No. of tRNA | 86 | – | – | – | – |
| MLST | ST1485 | – | – | – | – |
| GenBank ID | AP039418 | AP039419 | AP039420 | AP039421 | AP039422 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawano, K.; Masaki, T.; Kawaguchi, T.; Kuroda, M. Persistence of Colistin Resistance and mcr-1.1-Positive E. coli in Poultry Despite Colistin Ban in Japan. Antibiotics 2025, 14, 360. https://doi.org/10.3390/antibiotics14040360
Kawano K, Masaki T, Kawaguchi T, Kuroda M. Persistence of Colistin Resistance and mcr-1.1-Positive E. coli in Poultry Despite Colistin Ban in Japan. Antibiotics. 2025; 14(4):360. https://doi.org/10.3390/antibiotics14040360
Chicago/Turabian StyleKawano, Komei, Takayuki Masaki, Tatsuya Kawaguchi, and Makoto Kuroda. 2025. "Persistence of Colistin Resistance and mcr-1.1-Positive E. coli in Poultry Despite Colistin Ban in Japan" Antibiotics 14, no. 4: 360. https://doi.org/10.3390/antibiotics14040360
APA StyleKawano, K., Masaki, T., Kawaguchi, T., & Kuroda, M. (2025). Persistence of Colistin Resistance and mcr-1.1-Positive E. coli in Poultry Despite Colistin Ban in Japan. Antibiotics, 14(4), 360. https://doi.org/10.3390/antibiotics14040360