
Microbiome and Resistome in Poultry Litter-Fertilized and Unfertilized Agricultural Soils
 , , and
, , and
Abstract
1. Introduction
2. Results
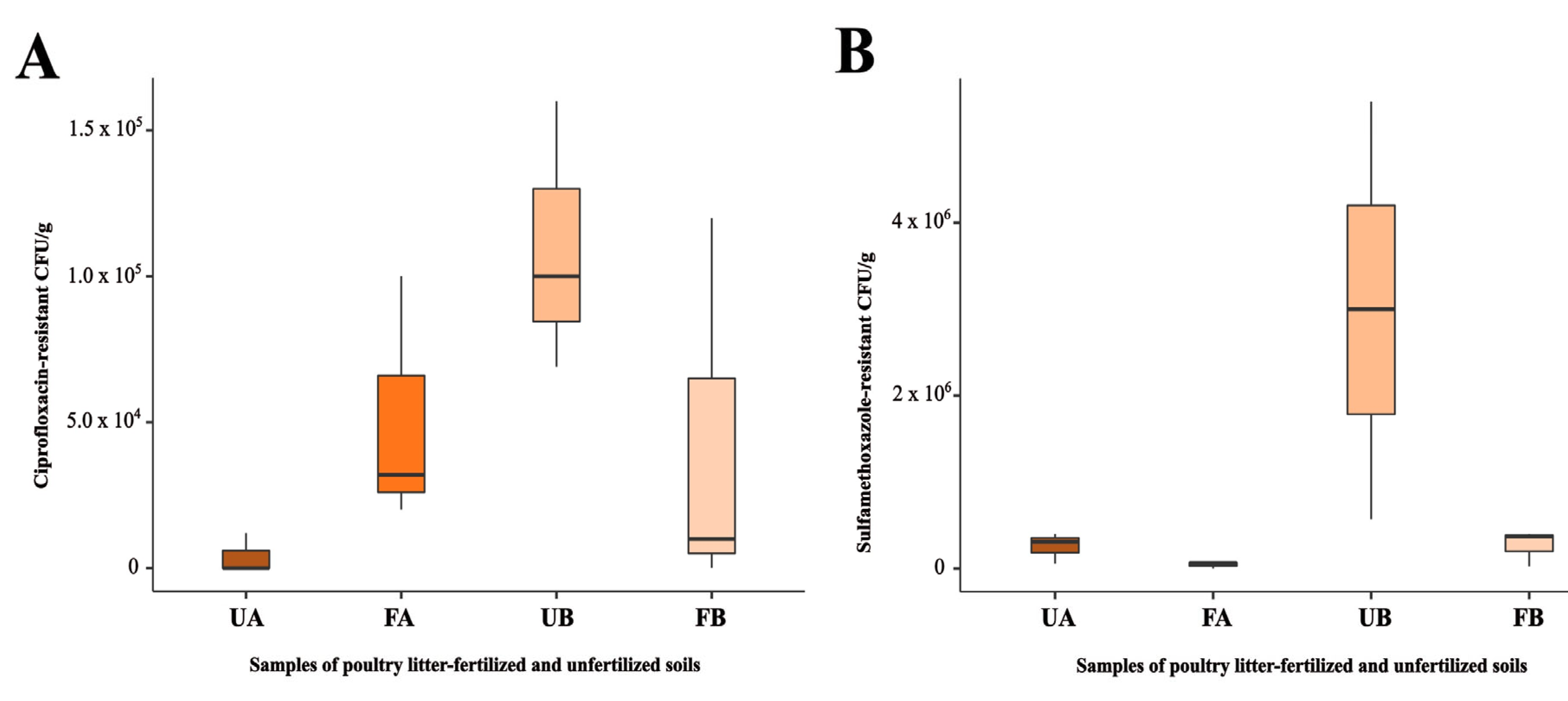
2.1. Antimicrobial-Resistant Bacteria Isolated from Agricultural Soils
2.2. Detection of Resistance Genes in Bacterial Isolates and Total DNA from the Same Agricultural Soils
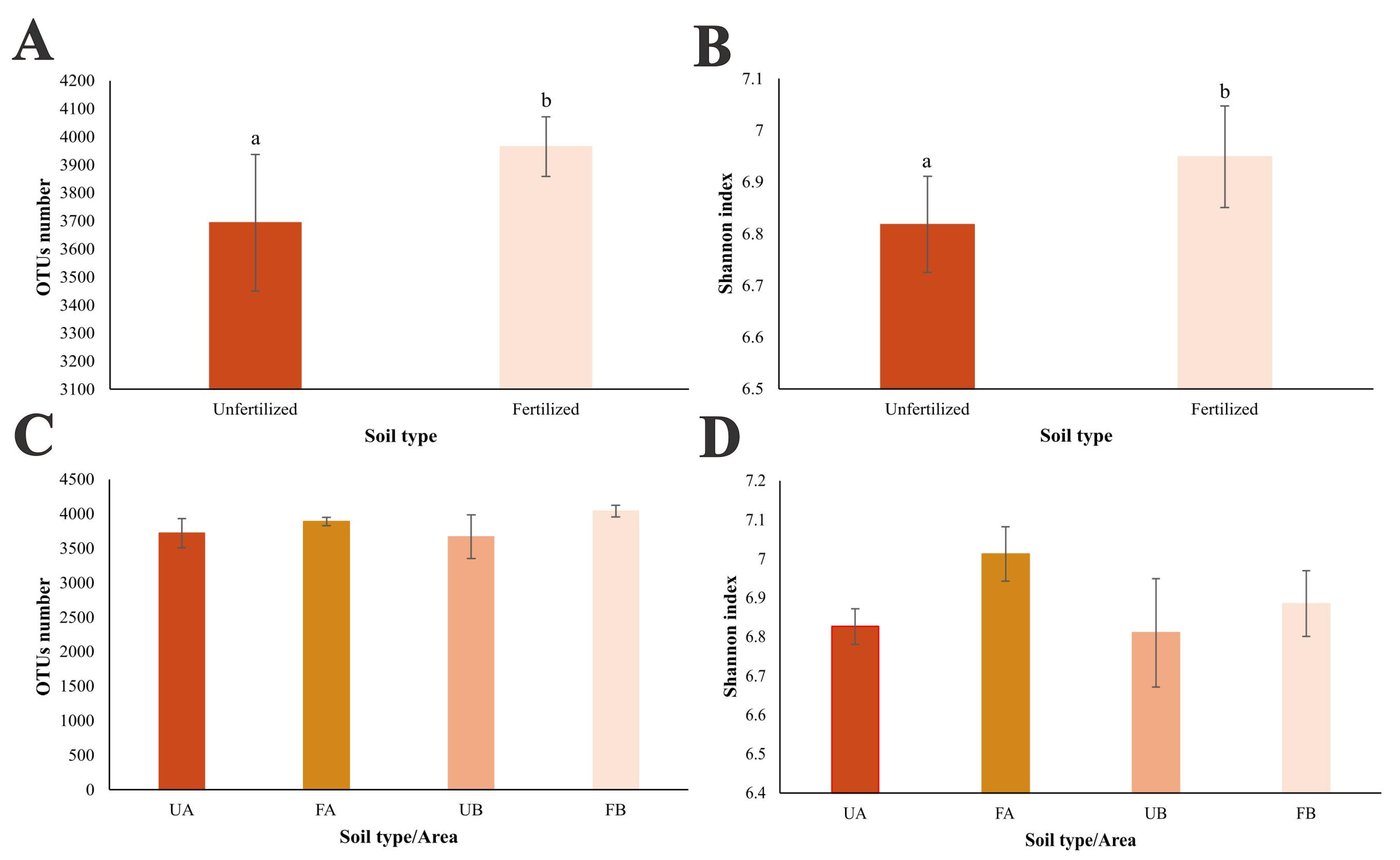
2.3. Structure of the Bacterial Community in Poultry Litter-Fertilized and Unfertilized Soils
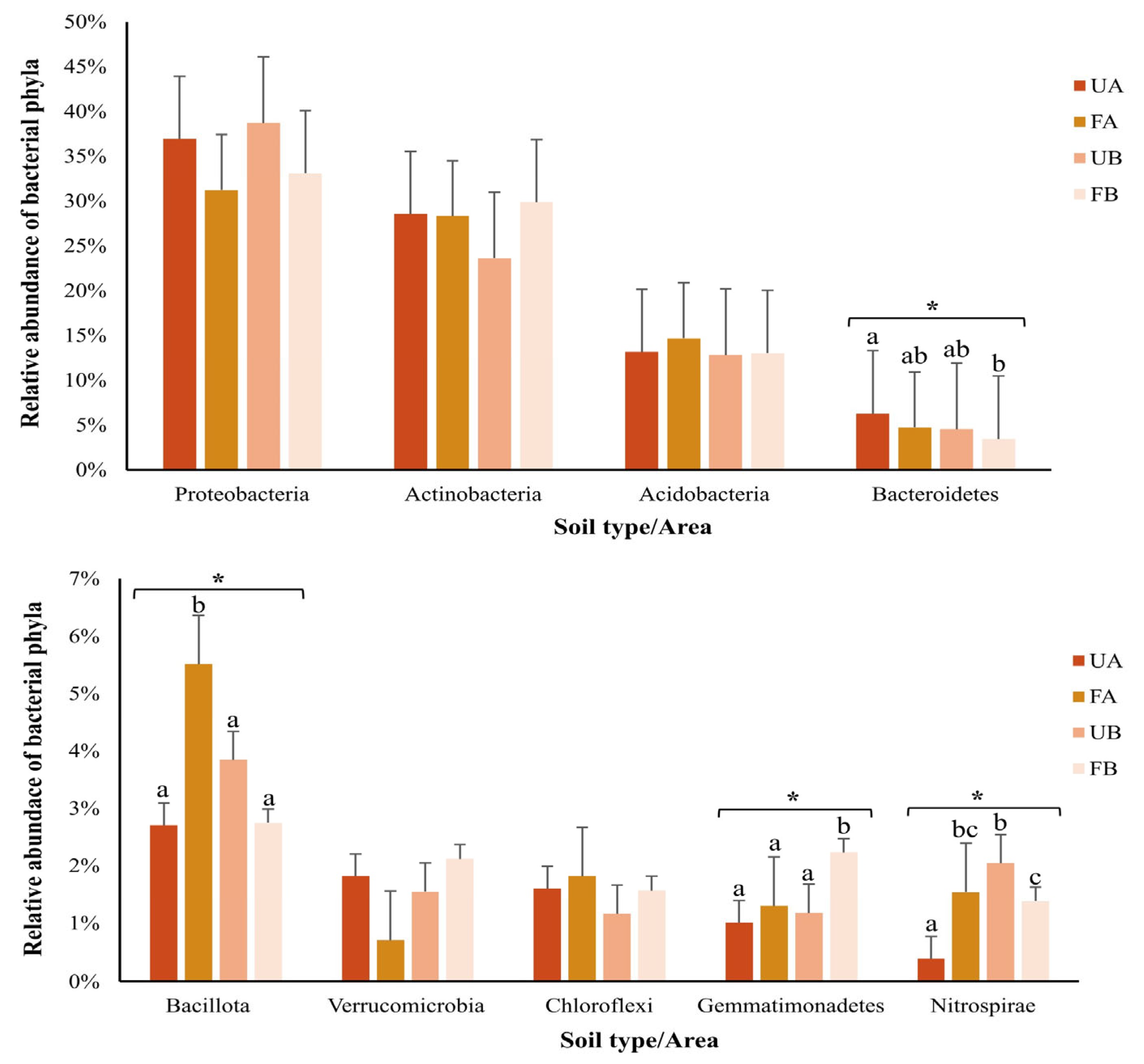
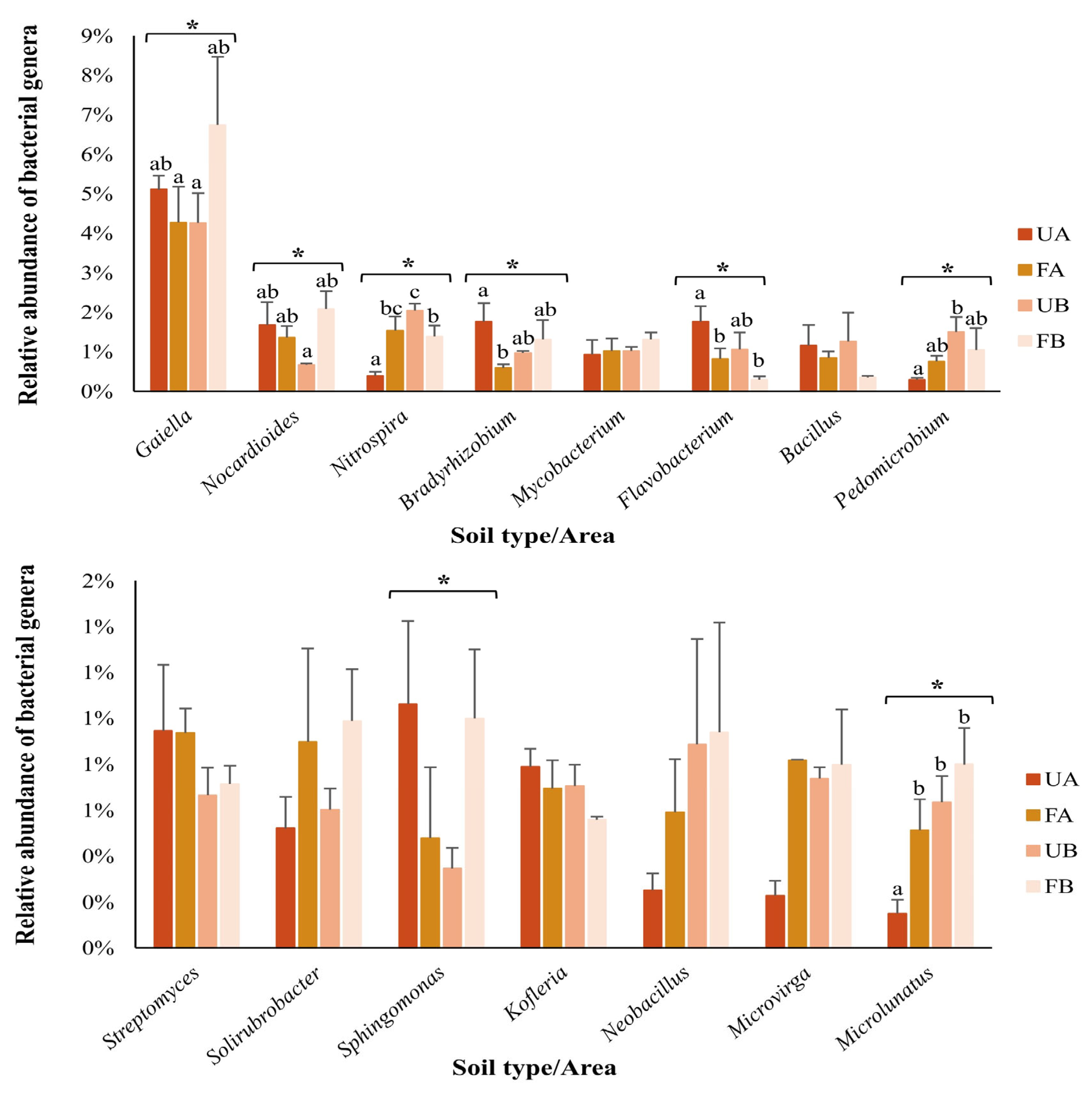
2.4. Composition of the Bacterial Community in Poultry Litter-Fertilized and Unfertilized Soils
3. Discussion
4. Materials and Methods
4.1. Study Area and Sampling
4.2. Isolation of Bacterial Strains Resistant to Different Antimicrobial Agents
4.3. Identification of Strains Isolated from Poultry Litter-Fertilized and Unfertilized Soils
4.4. Extraction of Total DNA from Soil Samples
4.5. Detection of Antimicrobial Resistance Genes
4.6. Analysis of the Total Bacterial Community in the Soil Samples Through Amplicon Sequencing of the Gene Encoding 16S rRNA
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Adekiya, A.O.; Ejue, W.S.; Olayanju, A.; Dunsin, O.; Aboyeji, C.M.; Aremu, C.; Adegbite, K.; Akinpelu, O. Different Organic Manure Sources and NPK Fertilizer on Soil Chemical Properties, Growth, Yield and Quality of Okra. Sci. Rep. 2020, 10, 16083. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.; Morra, M.J.; Stalder, T.; Jechalke, S.; Top, E.; Pollard, A.T.; Popova, I. Dairy Manure as a Potential Source of Crop Nutrients and Environmental Contaminants. J. Environ. Sci. 2021, 100, 117–130. [Google Scholar] [CrossRef]
- Ravindran, B.; Mupambwa, H.A.; Silwana, S.; Mnkeni, P.N.S. Assessment of Nutrient Quality, Heavy Metals and Phytotoxic Properties of Chicken Manure on Selected Commercial Vegetable Crops. Heliyon 2017, 3, e00493. [Google Scholar] [CrossRef] [PubMed]
- Vollú, R.E.; Cotta, S.R.; Jurelevicius, D.; de Assis Leite, D.C.; Parente, C.E.T.; Malm, O.; Martins, D.C.; Resende, Á.V.; Marriel, I.E.; Seldin, L. Response of the Bacterial Communities Associated With Maize Rhizosphere to Poultry Litter as an Organomineral Fertilizer. Front. Environ. Sci. 2018, 6, 118. [Google Scholar] [CrossRef]
- Parente, C.E.T.; Brusdzenski, G.S.; Zonta, E.; Lino, A.S.; Azevedo-Silva, C.E.; Dorneles, P.R.; Azeredo, A.; Torres, J.P.M.; Meire, R.O.; Malm, O. Fluoroquinolones and Trace Elements in Poultry Litter: Estimation of Environmental Load Based on Nitrogen Requirement for Crops. J. Environ. Sci. Health Part B 2020, 55, 1087–1098. [Google Scholar] [CrossRef]
- US Food and Drug Administration. US-FDA Guidance for Industry New Animal Drugs and New Animal Drug Combination Products Administered in or on Medicated Feed or Drinking Water of Food-Producing Animals: Recommendations for Drug Sponsors for Voluntarily Aligning Product Use Conditions with GFI #209; US Food and Drug Administration: Rockville, MD, USA, 2013.
- Sneeringer, S.; Macdonald, J.; Key, N.; Mcbride, W.; Mathews, K. Economic Research Service Economic Research Report Number 200 Economics of Antibiotic Use in U.S. Livestock Production; USDA: Washington, DC, USA, 2015.
- Palma, E.; Tilocca, B.; Roncada, P. Antimicrobial Resistance in Veterinary Medicine: An Overview. Int. J. Mol. Sci. 2020, 21, 1914. [Google Scholar] [CrossRef]
- Filippitzi, M.E.; Devreese, M.; Broekaert, K.; Rasschaert, G.; Daeseleire, E.; Meirlaen, J.; Dewulf, J. Quantitative Risk Model to Estimate the Level of Antimicrobial Residues That Can Be Transferred to Soil via Manure, Due to Oral Treatments of Pigs. Prev. Vet. Med. 2019, 167, 90–100. [Google Scholar] [CrossRef]
- Jechalke, S.; Heuer, H.; Siemens, J.; Amelung, W.; Smalla, K. Fate and Effects of Veterinary Antibiotics in Soil. Trends Microbiol. 2014, 22, 536–545. [Google Scholar] [CrossRef]
- Parente, C.E.T.; Azeredo, A.; Vollú, R.E.; Zonta, E.; Azevedo-Silva, C.E.; Brito, E.M.S.; Seldin, L.; Torres, J.P.M.; Meire, R.O.; Malm, O. Fluoroquinolones in Agricultural Soils: Multi-Temporal Variation and Risks in Rio de Janeiro Upland Region. Chemosphere 2019, 219, 409–417. [Google Scholar] [CrossRef]
- dos Santos Lopes, E.; Ferreira Santaren, K.C.; Araujo de Souza, L.C.; Parente, C.E.T.; Picão, R.C.; Jurelevicius, D.d.A.; Seldin, L. Cross-Environmental Cycling of Antimicrobial Resistance in Agricultural Areas Fertilized with Poultry Litter: A One Health Approach. Environ. Poll. 2024, 363, 125177. [Google Scholar] [CrossRef]
- Rahman, M.; Alam, M.-U.; Luies, S.K.; Kamal, A.; Ferdous, S.; Lin, A.; Sharior, F.; Khan, R.; Rahman, Z.; Parvez, S.M.; et al. Contamination of Fresh Produce with Antibiotic-Resistant Bacteria and Associated Risks to Human Health: A Scoping Review. Int. J. Environ. Res. Public Health 2021, 19, 360. [Google Scholar] [CrossRef]
- Marques, R.Z.; Wistuba, N.; Brito, J.C.M.; Bernardoni, V.; Rocha, D.C.; Gomes, M.P. Crop Irrigation (Soybean, Bean, and Corn) with Enrofloxacin-Contaminated Water Leads to Yield Reductions and Antibiotic Accumulation. Ecotoxicol. Environ. Saf. 2021, 216, 112193. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; Wellcome Trust: London, UK, 2016. [Google Scholar]
- WHO. Bacterial Priority List, 2024: Bacterial Pathogens of Public Health to Guide Research, Development and Strategies to Prevent and Control Resistance; WHO: Geneva, Switzerland, 2024. [Google Scholar]
- Huygens, J.; Daeseleire, E.; Mahillon, J.; Van Elst, D.; Decrop, J.; Meirlaen, J.; Dewulf, J.; Heyndrickx, M.; Rasschaert, G. Presence of Antibiotic Residues and Antibiotic Resistant Bacteria in Cattle Manure Intended for Fertilization of Agricultural Fields: A One Health Perspective. Antibiotics 2021, 10, 410. [Google Scholar] [CrossRef] [PubMed]
- Cytryn, E. The Soil Resistome: The Anthropogenic, the Native, and the Unknown. Soil Biol. Biochem. 2013, 63, 18–23. [Google Scholar] [CrossRef]
- Lopes, E.S.; Parente, C.E.T.; Picão, R.C.; Seldin, L. Irrigation Ponds as Sources of Antimicrobial-Resistant Bacteria in Agricultural Areas with Intensive Use of Poultry Litter. Antibiotics 2022, 11, 1650. [Google Scholar] [CrossRef]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A Web-Based Tool for the Analysis of Sets through Venn Diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Macedo, G.; van Veelen, H.P.J.; Hernandez-Leal, L.; van der Maas, P.; Heederik, D.; Mevius, D.; Bossers, A.; Schmitt, H. Targeted Metagenomics Reveals Inferior Resilience of Farm Soil Resistome Compared to Soil Microbiome after Manure Application. Sci. Total Environ. 2021, 770, 145399. [Google Scholar] [CrossRef]
- Cycoń, M.; Mrozik, A.; Piotrowska-Seget, Z. Antibiotics in the Soil Environment—Degradation and Their Impact on Microbial Activity and Diversity. Front. Microbiol. 2019, 10, 338. [Google Scholar] [CrossRef]
- Aldred, K.J.; Kerns, R.J.; Osheroff, N. Mechanism of Quinolone Action and Resistance. Biochemistry 2014, 53, 1565–1574. [Google Scholar] [CrossRef]
- Zhu, Y.-G.; Zhao, Y.; Zhu, D.; Gillings, M.; Penuelas, J.; Ok, Y.S.; Capon, A.; Banwart, S. Soil Biota, Antimicrobial Resistance and Planetary Health. Environ. Int. 2019, 131, 105059. [Google Scholar] [CrossRef]
- Furlan, J.P.R.; Stehling, E.G.; Pitondo-Silva, A. Importance of Sequencing to Determine Functional blaTEM Variants. Antimicrob. Agents Chemother. 2017, 61, 10–1128. [Google Scholar] [CrossRef]
- Furlan, J.P.R.; Stehling, E.G. Detection of β-Lactamase Encoding Genes in Feces, Soil and Water from a Brazilian Pig Farm. Environ. Monit. Assess. 2018, 190, 76. [Google Scholar] [CrossRef]
- Haghighi, S.; Reza Goli, H. High Prevalence of blaVEB, blaGES and blaPER Genes in Beta-Lactam Resistant Clinical Isolates of Pseudomonas aeruginosa. AIMS Microbiol. 2022, 8, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Röderova, M.; Halova, D.; Papousek, I.; Dolejska, M.; Masarikova, M.; Hanulik, V.; Pudova, V.; Broz, P.; Htoutou-Sedlakova, M.; Sauer, P.; et al. Characteristics of Quinolone Resistance in Escherichia coli Isolates from Humans, Animals, and the Environment in the Czech Republic. Front. Microbiol. 2017, 7, 2147. [Google Scholar] [CrossRef]
- Yang, H.Y.; Nam, Y.S.; Lee, H.J. Prevalence of Plasmid-Mediated Quinolone Resistance Genes among Ciprofloxacin-Nonsusceptible Escherichia coli and Klebsiella Pneumoniae Isolated from Blood Cultures in Korea. Can. J. Inf. Dis. Med. Micro 2014, 25, 163–169. [Google Scholar] [CrossRef]
- Taha, S.A.; Omar, H.H.; Hassan, w.H. Characterization of Plasmid-Mediated qnrA and qnrB Genes among Enterobacteriaceae Strains: Quinolone Resistance and ESBL Production in Ismailia, Egypt. Egypt. J. Med. Hum. Gen. 2019, 20, 26. [Google Scholar] [CrossRef]
- Liu, C.; Yan, H.; Sun, Y.; Chen, B. Contribution of Enrofloxacin and Cu2+ to the Antibiotic Resistance of Bacterial Community in a River Biofilm. Environ. Poll. 2021, 291, 118156. [Google Scholar] [CrossRef]
- Zhao, J.R.; Fan, X.Y.; Li, X.; Gao, Y.X.; Zhang, Z.X. Impact of Ciprofloxacin and Copper Combined Pollution on Activated Sludge: Abundant-Rare Taxa and Antibiotic Resistance Genes. Bioresour. Technol. 2022, 349, 126882. [Google Scholar] [CrossRef]
- Alvarez-Arevalo, M.; Sterndorff, E.B.; Faurdal, D.; Mourched, A.-S.; Charusanti, P.; Jørgensen, T.S.; Weber, T. Complete, Circular Genome Sequence of a Bosea sp. Isolate from Soil. Microbiol. Resour. Announc. 2023, 12. [Google Scholar] [CrossRef]
- Mazel, D. Integrons: Agents of bacterial evolution. Nat. Rev. Microbiol. 2006, 4, 608–620. [Google Scholar] [CrossRef]
- Gillings, M.R. Integrons: Past, Present, and Future. Microbiol. Mol. Biol. Rev. 2014, 78, 257–277. [Google Scholar] [CrossRef] [PubMed]
- Gillings, M.R.; Gaze, W.H.; Pruden, A.; Smalla, K.; Tiedje, J.M.; Zhu, Y.G. Using the class 1 integron-integrase gene as a proxy for anthropogenic pollution. ISME J. 2015, 9, 1269–1279. [Google Scholar] [CrossRef] [PubMed]
- Freitag, C.; Michael, G.B.; Kadlec, K.; Hassel, M.; Schwarz, S. Detection of plasmid-borne extended-spectrum β-lactamase (ESBL) genes in Escherichia coli isolates from bovine mastitis. Vet. Microbiol. 2017, 200, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Parente, C.E.T.; Brito, E.M.S.; Caretta, C.A.; Cervantes-Rodríguez, E.A.; Fábila-Canto, A.P.; Vollú, R.E.; Seldin, L.; Malm, O. Bacterial Diversity Changes in Agricultural Soils Influenced by Poultry Litter Fertilization. Braz. J. Micro 2021, 52, 675–686. [Google Scholar] [CrossRef]
- Minkina, T.; Sushkova, S.; Delegan, Y.; Bren, A.; Mazanko, M.; Kocharovskaya, Y.; Filonov, A.; Rajput, V.D.; Mandzhieva, S.; Rudoy, D.; et al. Effect of Chicken Manure on Soil Microbial Community Diversity in Poultry Keeping Areas. Environ. Geochem. Health 2023, 45, 9303–9319. [Google Scholar] [CrossRef]
- Xiao, Y.; Xiang, Y.; Zhou, W.; Chen, J.; Li, K.; Yang, H. Microbial Community Mapping in Intestinal Tract of Broiler Chicken. Poult. Sci. 2017, 96, 1387–1393. [Google Scholar] [CrossRef]
- Han, X.-M.; Hu, H.-W.; Chen, Q.-L.; Yang, L.-Y.; Li, H.-L.; Zhu, Y.-G.; Li, X.-Z.; Ma, Y.-B. Antibiotic Resistance Genes and Associated Bacterial Communities in Agricultural Soils Amended with Different Sources of Animal Manures. Soil Biol. Biochem. 2018, 126, 91–102. [Google Scholar] [CrossRef]
- Sood, U.; Gupta, V.; Kumar, R.; Lal, S.; Fawcett, D.; Rattan, S.; Poinern, G.E.J.; Lal, R. Chicken Gut Microbiome and Human Health: Past Scenarios, Current Perspectives, and Futuristic Applications. Indian J. Microbiol. 2020, 60, 2–11. [Google Scholar] [CrossRef]
- Pin Viso, N.D.; Rizzo, P.F.; Young, B.J.; Gabioud, E.; Bres, P.; Riera, N.I.; Merino, L.; Farber, M.D.; Crespo, D.C. The Use of Raw Poultry Waste as Soil Amendment Under Field Conditions Caused a Loss of Bacterial Genetic Diversity Together with an Increment of Eutrophic Risk and Phytotoxic Effects. Microb. Ecol. 2023, 86, 1082–1095. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, M.; Liu, M.; Cheng, C.; Qiao, L.; Li, Y.; Li, Z. Responses of Soil Available Nutrients and Microbial Performance in a Newly Established Apple Orchard after Five-Year Fertilization with Different Sources of Livestock Manure. J. Agric. Food Res. 2025, 19, 101635. [Google Scholar] [CrossRef]
- Sun, Y.; Guo, Y.; Shi, M.; Qiu, T.; Gao, M.; Tian, S.; Wang, X. Effect of Antibiotic Type and Vegetable Species on Antibiotic Accumulation in Soil-Vegetable System, Soil Microbiota, and Resistance Genes. Chemosphere 2021, 263, 128099. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. Antibiotic Resistance in the Environment: A Link to the Clinic? Curr. Opin. Microbiol. 2010, 13, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Gaze, W.H.; Krone, S.M.; Larsson, D.G.J.; Li, X.-Z.; Robinson, J.A.; Simonet, P.; Smalla, K.; Timinouni, M.; Topp, E.; Wellington, E.M.; et al. Influence of Humans on Evolution and Mobilization of Environmental Antibiotic Resistome. Emerg. Infect. Dis. 2013, 19, e120871. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.-Y.; Shen, Q.; Zhao, F.J. Antibiotics and Antibiotic Resistance from Animal Manures to Soil: A Review. Eur. J. Soil Sci. 2018, 69, 181–195. [Google Scholar] [CrossRef]
- Bai, H.; He, L.-Y.; Wu, D.-L.; Gao, F.-Z.; Zhang, M.; Zou, H.-Y.; Yao, M.-S.; Ying, G.-G. Spread of Airborne Antibiotic Resistance from Animal Farms to the Environment: Dispersal Pattern and Exposure Risk. Environ. Int. 2022, 158, 106927. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Hu, H.W.; Chen, Q.L.; Singh, B.K.; Yan, H.; Chen, D.; He, J.Z. Transfer of antibiotic resistance from manure-amended soils to vegetable microbiomes. Environ. Int. 2019, 130, 104912. [Google Scholar]
- Sun, Y.; Snow, D.; Walia, H.; Li, X. Transmission Routes of the Microbiome and Resistome from Manure to Soil and Lettuce. Environ. Sci. Technol. 2021, 55, 11102–11112. [Google Scholar] [CrossRef]
- Rosi, A.; Canavesi, V.; Segoni, S.; Dias Nery, T.; Catani, F.; Casagli, N. Landslides in the Mountain Region of Rio de Janeiro: A Proposal for the Semi-Automated Definition of Multiple Rainfall Thresholds. Geosciences 2019, 9, 203. [Google Scholar] [CrossRef]
- Santos, H.G.; Jacomine, P.K.T.; Anjos, L.H.C.; Oliveira, V.A.; Lumbreras, J.F.; Coelho, M.R.; Almeida, J.A.; Araújo Filho, J.C.; Oliveira, J.B.; Cunha, T.J.F. Sistema Brasileiro de Classificação de Solos, 5th ed.; Embrapa: Brasília, DF, Brazil, 2018; ISBN 9788570358004. [Google Scholar]
- USDA—United States Department of Agriculture. Natural Resources Conservation Service Soil Taxonomy: A Basic System of Soil Classification for Making and Interpreting Soil Surveys, 2nd ed.; USDA: Washington, DC, USA, 1999.
- Massol-Deya, A.A.; Odelson, D.A.; Hickey, R.F. Bacterial Community Fingerprinting of Amplified 16S and 16–23S Ribosomal DNA Gene Sequences and Restriction Endonuclease Analysis (ARDRA). In Molecular Microbial Ecology Manual; Akkermans, A.D.L., Van Elsas, J.D., De Bruijn, F.J., Eds.; Springer: Dordrecht, The Netherlands, 1995; pp. 289–296. ISBN 978-94-010-4156-0. [Google Scholar]
- Xu, X.; Kong, F.; Cheng, X.; Yan, B.; Du, X.; Gai, J.; Ai, H.; Shi, L.; Iredell, J. Integron Gene Cassettes in Acinetobacter spp. Strains from South China. Int. J. Antimicrob. Agents 2008, 32, 441–445. [Google Scholar] [CrossRef]
- Campana, E.H.; Xavier, D.E.; Petrolini, F.V.-B.; Cordeiro-Moura, J.R.; de Araujo, M.R.E.; Gales, A.C. Carbapenem-Resistant and Cephalosporin-Susceptible: A Worrisome Phenotype among Pseudomonas aeruginosa Clinical Isolates in Brazil. Braz. J. Inf. Dis. 2017, 21, 57–62. [Google Scholar] [CrossRef]
- Picão, R.C.; Poirel, L.; Gales, A.C.; Nordmann, P. Diversity of β-Lactamases Produced by Ceftazidime-Resistant Pseudomonas aeruginosa Isolates Causing Bloodstream Infections in Brazil. Antimicrob. Agents Chemother. 2009, 53, 3908–3913. [Google Scholar] [CrossRef]
- Toleman, M.A.; Bennett, P.M.; Bennett, D.M.C.; Jones, R.N.; Walsh, T.R. Global Emergence of Trimethoprim/Sulfamethoxazole Resistance in Stenotrophomonas maltophilia Mediated by Acquisition of sul Genes. Emerg. Inf. Dis. 2007, 13, 559–565. [Google Scholar] [CrossRef]
- Cattoir, V.; Poirel, L.; Rotimi, V.; Soussy, C.-J.; Nordmann, P. Multiplex PCR for Detection of Plasmid-Mediated Quinolone Resistance qnr Genes in ESBL-Producing Enterobacterial Isolates. J. Antimicrob. Chemother. 2007, 60, 394–397. [Google Scholar] [CrossRef]
- Sundberg, C.; Al-Soud, W.A.; Larsson, M.; Alm, E.; Yekta, S.S.; Svensson, B.H.; Sørensen, S.J.; Karlsson, A. 454 Pyrosequencing Analyses of Bacterial and Archaeal Richness in 21 Full-Scale Biogas Digesters. FEMS Microbiol. Ecol. 2013, 85, 612–626. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Hammer, R.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Paleontol. Electron. 2001, 4, 1. [Google Scholar]
- The Jamovi Project. Jamovi, version 2.3. Computer Software. The Jamovi Project: Sydney, Australia, 2022. Available online: https://www.jamovi.org (accessed on 14 February 2025).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Identification | Origen | sul1 | sul2 | int1 | int2 | qnrA | qnrB | qnrS | blaTEM | blaGES | blaSHV |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SULFAMETHOXAZOLE | ||||||||||||
| SSFA5.2 | Bacillus sp. | FA | − | − | − | − | − | − | − | + | − | − |
| SSUA3.1 | Klebsiella sp. | UA | − | − | − | − | − | − | − | − | − | + |
| SSUA3.2 | Klebsiella variicola | UA | − | − | − | − | − | − | − | + | − | + |
| SSUA4.1 | Pseudomonas sp. | UA | − | − | + | + | − | − | − | − | − | − |
| SSUA12 | Klebsiella sp. | UA | − | − | − | − | − | − | − | − | − | + |
| SSFB3.1 | Bacillus sp. | FB | − | − | − | − | − | − | − | + | − | − |
| SSFB5.2 | Bacillus sp. | FB | − | − | + | − | − | − | − | − | − | − |
| SSFB9.1 | Citrobacter sp. | FB | − | − | − | − | + | − | − | + | − | − |
| SSFB12 | Citrobacter sp. | FB | − | − | − | − | + | − | − | − | − | − |
| SSUB4.1 | Bacillus sp. | UB | − | − | − | − | − | − | − | + | − | − |
| SSUB4.2 | Bacillus sp. | UB | − | − | − | − | − | − | − | + | + | − |
| SSUB5.1 | Lynsinibacillus sp. | UB | − | − | − | − | − | − | − | + | − | − |
| SSUB6.1 | Bacillus sp. | UB | − | − | − | − | − | − | − | + | + | − |
| SSUB11 | Citrobacter sp. | UB | − | − | − | − | − | + | − | + | − | − |
| SSUB13 | Proteobacteria | UB | − | − | − | − | − | − | − | + | − | − |
| CIPROFLOXACIN | ||||||||||||
| CSUA1.2 | Bosea sp. | UA | − | − | − | − | − | − | − | + | − | + |
| CSUA4.1 | Bosea sp. | UA | − | − | − | − | + | − | − | − | − | + |
| CSUB5 | Stenotrophomonas sp. | UB | − | − | − | − | + | − | − | − | − | − |
| CSUB8 | Stenotrophomonas sp. | UB | − | − | − | − | + | − | − | − | − | − |
| CSFB1.2 | Microbacterium sp. | FB | − | − | − | − | − | − | − | + | − | − |
| CSFB5.1 | Bosea sp. | FB | − | − | − | − | − | − | − | − | − | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopes, E.d.S.; de Souza, L.C.A.; Santaren, K.C.F.; Parente, C.E.T.; Seldin, L. Microbiome and Resistome in Poultry Litter-Fertilized and Unfertilized Agricultural Soils. Antibiotics 2025, 14, 355. https://doi.org/10.3390/antibiotics14040355
Lopes EdS, de Souza LCA, Santaren KCF, Parente CET, Seldin L. Microbiome and Resistome in Poultry Litter-Fertilized and Unfertilized Agricultural Soils. Antibiotics. 2025; 14(4):355. https://doi.org/10.3390/antibiotics14040355
Chicago/Turabian StyleLopes, Eliene dos Santos, Larissa Coutinho Araujo de Souza, Karen Caroline Ferreira Santaren, Cláudio Ernesto Taveira Parente, and Lucy Seldin. 2025. "Microbiome and Resistome in Poultry Litter-Fertilized and Unfertilized Agricultural Soils" Antibiotics 14, no. 4: 355. https://doi.org/10.3390/antibiotics14040355
APA StyleLopes, E. d. S., de Souza, L. C. A., Santaren, K. C. F., Parente, C. E. T., & Seldin, L. (2025). Microbiome and Resistome in Poultry Litter-Fertilized and Unfertilized Agricultural Soils. Antibiotics, 14(4), 355. https://doi.org/10.3390/antibiotics14040355