

Postbiotic-Based Extracts from Native Probiotic Strains: A Promising Strategy for Food Preservation and Antimicrobial Defense
 , ,
, ,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
2.1. PBEs Co-Cultivated with L1PEag1 Inhibit the Cell Viability in a Dose-, Time-, and Producer Strain-Dependent Manner
2.2. Interaction of PBEs with E. coli L1PEag1 Led to Alterations in Cell Morphology and Ultrastructure
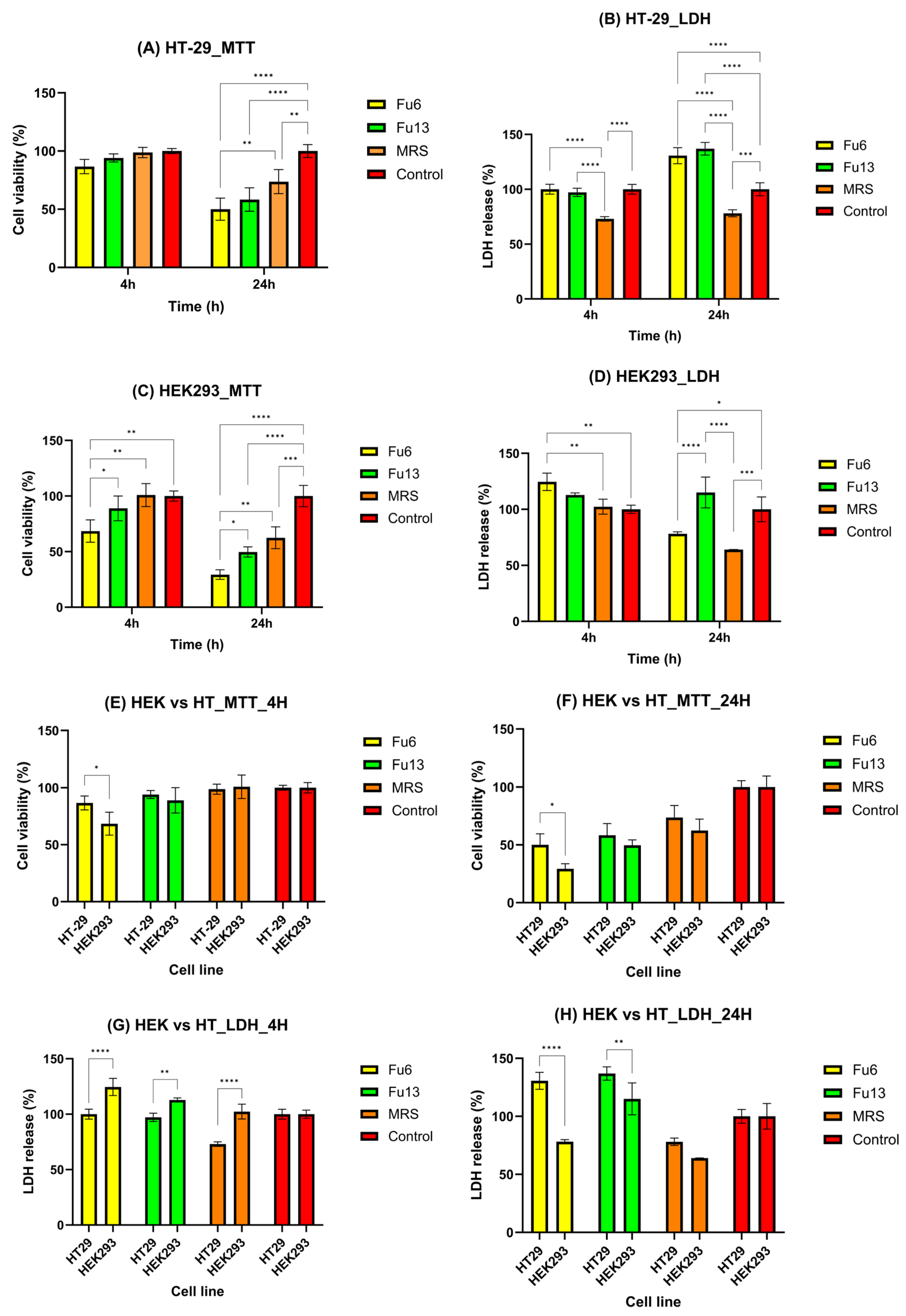
2.3. Cytotoxicity Evaluation of PBEs
2.4. Hemocompatibility
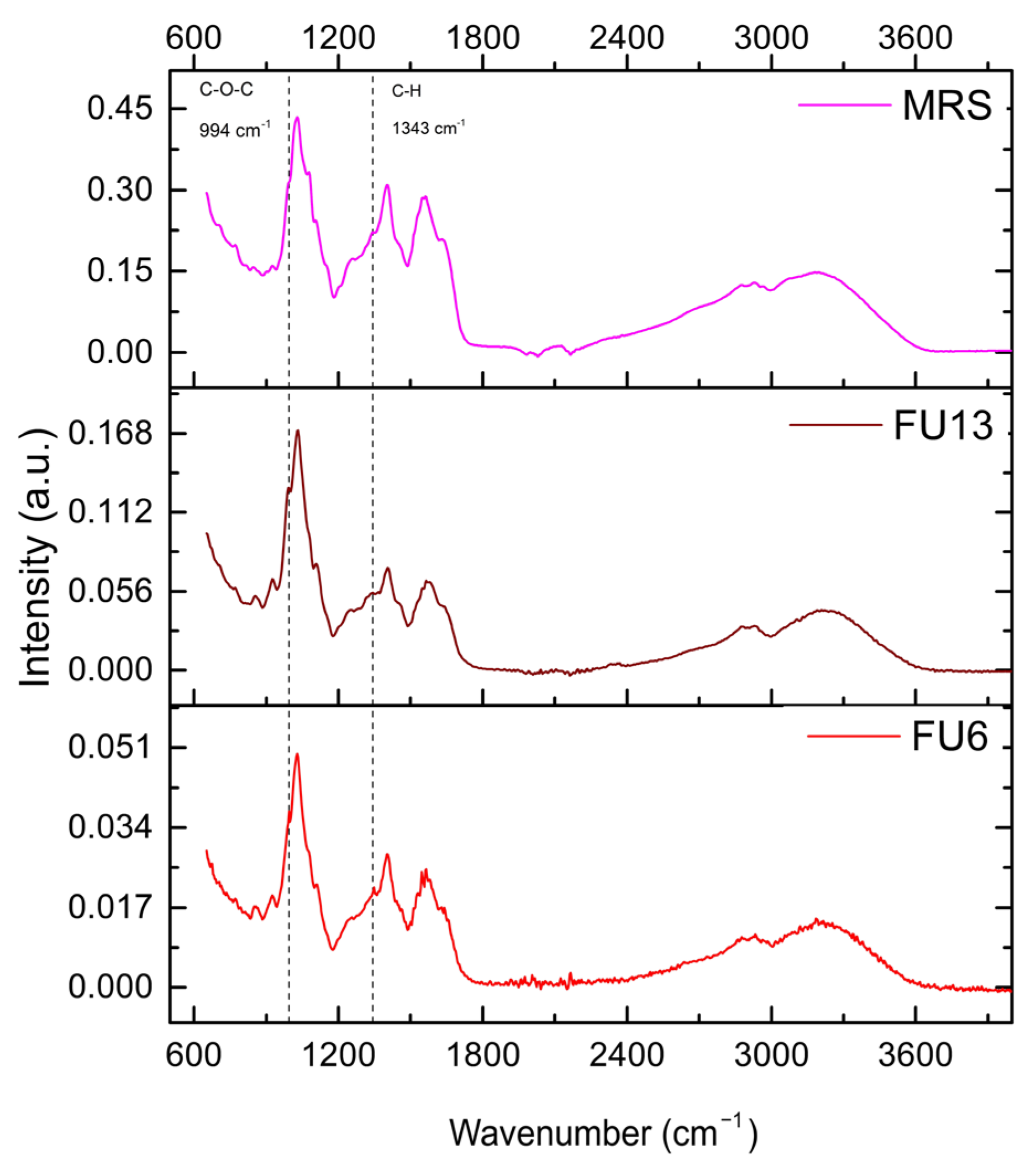
2.5. Bioactive Molecules Composition of PBEs Revealed by Infrared Spectrometry
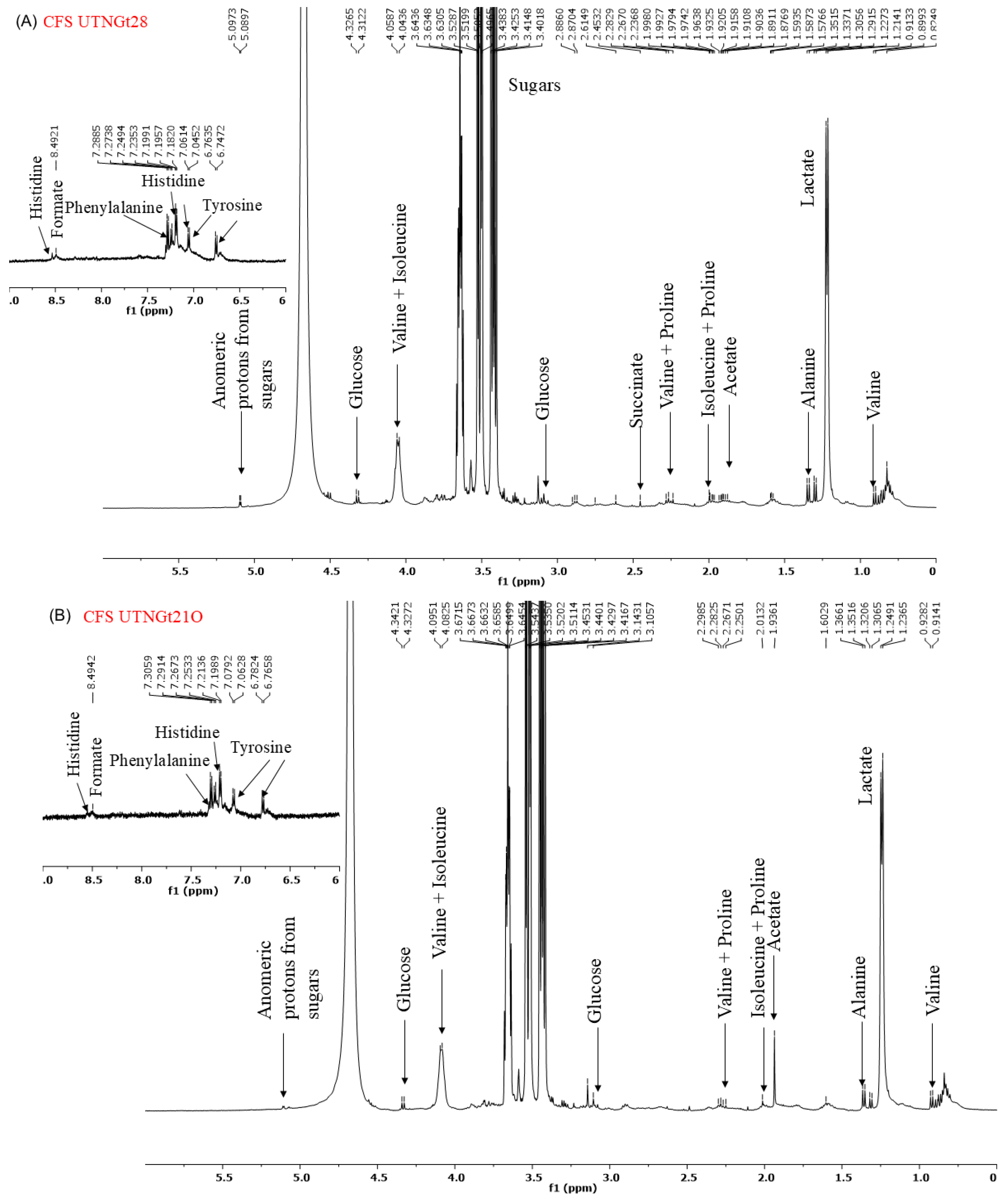
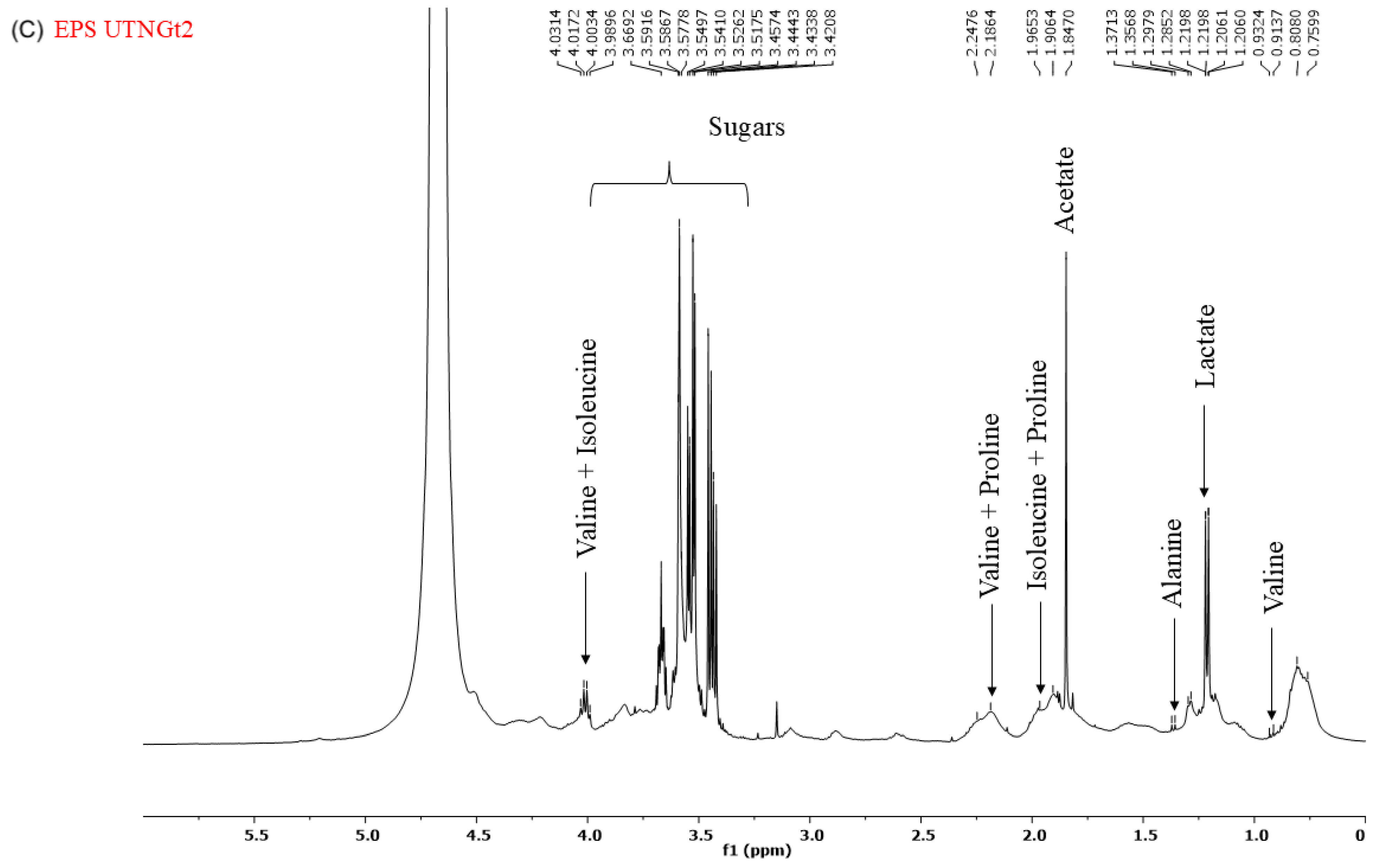
2.6. 1H-NMR Spectrum of CFS and EPS
2.7. Antioxidant Activity of PBEs
3. Materials and Methods
3.1. Bacterial Strains and Growth Conditions
3.2. Extraction of CFS and EPS, and Selection of PBEs with High Antimicrobial Activity Against E. coli L1PEag1
3.3. Characterization of Selected PBEs
3.3.1. Antimicrobial Activity
Determination of Minimum Inhibitory Concentration (MIC)
Time-Killing Assay
The Examination of Ultrastructural and Morphological Changes in E. coli L1PEag1 upon PBEs Treatment
3.3.2. Biocompatibility Assessment
Cells Viability Determination: MTT Assay
Lactate Dehydrogenase (LDH) Release Assay
Hemolytic Effect on Red Blood Cells
3.3.3. ATR-FTIR Assay
3.3.4. 1H-NMR Assay
3.3.5. Antioxidant Activity
DPPH (1,1-Diphenyl-2-picrylhydrazyl) Assay
CUPRAC (Cupric Reducing Antioxidant Capacity) Assay
FRAP (Ferric Reducing Antioxidant Power) Assay
TEAC (Trolox Equivalent Antioxidant Capacity) Assay
3.4. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Wu, J.; Lv, M.; Shao, Z.; Hungwe, M.; Wang, J.; Bai, X.; Xie, J.; Wang, Y.; Geng, W. Metabolism characteristics of lactic acid bacteria and the expanding applications in food industry. Front. Bioeng. Biotechnol. 2021, 9, 612285. [Google Scholar] [CrossRef]
- Isaac-Bamgboye, F.J.; Mgbechidinma, C.L.; Onyeaka, H.; Isaac-Bamgboye, I.T.; Chukwugozie, D.C. Exploring the potential of postbiotics for food safety and human health improvement. J. Nutr. Metab. 2024, 2024, 1868161. [Google Scholar] [CrossRef] [PubMed]
- Vera-Santander, V.E.; Hernández-Figueroa, R.H.; Jiménez-Munguía, M.T.; Mani-López, E.; López-Malo, A. Health benefits of consuming foods with bacterial probiotics, postbiotics, and their metabolites: A review. Molecules 2023, 28, 1230. [Google Scholar] [CrossRef] [PubMed]
- Thorakkattu, P.; Khanashyam, A.C.; Shah, K.; Babu, K.S.; Mundanat, A.S.; Deliephan, A.; Deokar, G.S.; Santivarangkna, C.; Nirmal, N.P. Postbiotics: Current trends in food and pharmaceutical industry. Foods 2022, 11, 3094. [Google Scholar] [CrossRef] [PubMed]
- Scott, E.; De Paepe, K.; Van de Wiele, T. Postbiotics and Their Health Modulatory Biomolecules. Biomolecules 2022, 12, 1640. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Tumbarski, Y.; Nikolova, R.; Petkova, N.; Ivanov, I.; Lante, A. Biopreservation of fresh strawberries by carboxymethyl cellulose edible coatings enriched with a bacteriocin from Bacillus methylotrophicus BM47. Food Technol. Biotechnol. 2019, 57, 230–237. [Google Scholar] [CrossRef]
- Vieira, A.I.; Guerreiro, A.; Antunes, M.D.; Miguel MD, G.; Faleiro, M.L. Edible coatings enriched with essential oils on apples impair the survival of bacterial pathogens through a simulated gastrointestinal system. Foods 2019, 8, 57. [Google Scholar] [CrossRef]
- Tsilingiri, K.; Barbosa, T.; Penna, G.; Caprioli, F.; Sonzogni, A.; Viale, G.; Rescigno, M. Probiotic and postbiotic activity in health and disease: Comparison on a novel polarised ex-vivo organ culture model. Gut 2012, 61, 1007–1015. [Google Scholar] [CrossRef]
- Cicenia, A.; Scirocco, A.; Carabotti, M.; Pallotta, L.; Marignani, M.; Severi, C. Postbiotic activities of lactobacilli-derived factors. J. Clin. Gastroenterol. 2014, 48 (Suppl. S1), S18–S22. [Google Scholar] [CrossRef]
- Rajoka, M.S.R.; Zhao, H.; Lu, Y.; Lian, Z.; Li, N.; Hussain, N.; Shao, D.; Jin, M.; Li, Q.; Shi, J. Anticancer potential against cervix cancer (HeLa) cell line of probiotic Lactobacillus casei and Lactobacillus paracasei strains isolated from human breast milk. Food Funct. 2018, 9, 2705–2715. [Google Scholar] [CrossRef]
- Islam, S.; Biswas, S.; Jabin, T.; Moniruzzaman, M.; Biswas, J.; Uddin, M.S.; Akhtar-E-Ekram, M.; Elgorban, A.M.; Ghodake, G.; Syed, A.; et al. Probiotic potential of Lactobacillus plantarum DMR14 for preserving and extending shelf life of fruits and fruit juice. Heliyon 2023, 9, e17382. [Google Scholar] [CrossRef] [PubMed]
- Biswas, I.; Das Mohapatra, P.K. Recent advancement in metabiotics: A consortium with bioactive molecules after fermentation by probiotic bacteria with multidisciplinary application potential and future solution in health sector. Bioresour. Technol. Rep. 2023, 23, 101583. [Google Scholar] [CrossRef]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and inputs from lactic acid bacteria and their bacteriocins as alternatives to antibiotic growth promoters during food-animal production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef]
- Tenea, G.N. Postbiotics: A solution to protect tropical fruits towards postharvest adulteration. AgroLife Sci. J. 2021, 10, 189–196. [Google Scholar] [CrossRef]
- Tenea, G.N.; Angamarca, E.; Olmedo, D. Combinations of peptide-protein extracts from native probiotics suppress the growth of multidrug-resistant Staphylococcus aureus and Citrobacter freundii via membrane perturbation and ultrastructural changes. Antibiotics 2022, 11, 154. [Google Scholar] [CrossRef] [PubMed]
- Tenea, G.N.; Angamarca, E.; Cifuentes, V.; Hidalgo, J. Preventing microbe colonization on avocado (Persea nubigena var. guatemalensis) through metabiotic treatment, a promising postharvest safety improvement. Front. Microbiol. 2024, 15, 1344735. [Google Scholar] [CrossRef]
- Molina, D.; Carrión-Olmedo, J.C.; Jarrín-V, P.; Tenea, G.N. Genome characterization of a multi-drug resistant Escherichia coli strain, L1PEag1, isolated from commercial cape gooseberry fruits (Physalis peruviana L.). Front. Microbiol. 2024, 15, 1392333. [Google Scholar] [CrossRef]
- Rocchetti, M.T.; Russo, P.; Capozzi, V.; Drider, D.; Spano, G.; Fiocco, D. Bioprospecting antimicrobials from Lactiplantibacillus plantarum: Key factors underlying its probiotic action. Int. J. Mol. Sci. 2021, 22, 12076. [Google Scholar] [CrossRef]
- Kang, C.E.; Park, Y.J.; Kim, J.H.; Lee, N.K.; Paik, H.D. Probiotic Weissella cibaria displays antibacterial and anti-biofilm effect against cavity-causing Streptococcus mutans. Microb. Pathog. 2023, 180, 106151. [Google Scholar] [CrossRef]
- Tenea, G.N.; Hurtado, P. Next-generation sequencing for whole-genome characterization of Weissella cibaria UTNGt21O strain originated from wild Solanum quitoense Lam. fruits: An atlas of metabolites with biotechnological significance. Front. Microbiol. 2021, 12, 675002. [Google Scholar] [CrossRef]
- Remus, D.M.; van Kranenburg, R.; van Swam, I.I.; Taverne, N.; Bongers, R.S.; Wels, M.; Wells, J.M.; Bron, P.A.; Kleerebezem, M. Impact of 4 Lactobacillus plantarum capsular polysaccharide clusters on surface glycan composition and host cell signaling. Microb. Cell Fact. 2012, 11, 149. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, A.K.; Ayyash, M.M.; Olaimat, A.N.; Osaili, T.M.; Al-Nabulsi, A.A.; Shah, N.P.; Holley, R. Exopolysaccharides as antimicrobial agents: Mechanism and spectrum of activity. Front. Microbiol. 2021, 12, 664395. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.P.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, X.; Teng, D.; Mao, R.; Hao, Y.; Yang, N.; Wang, X.; Li, Z.; Wang, X.; Wang, J. Development of chimeric peptides to facilitate the neutralization of lipopolysaccharides during bactericidal targeting of multidrug-resistant Escherichia coli. Commun. Biol. 2020, 3, 41. [Google Scholar] [CrossRef]
- Wang, G.; Zeng, H. Antibacterial effect of cell-free supernatant from Lactobacillus pentosus L-36 against Staphylococcus aureus from bovine mastitis. Molecules 2022, 27, 7627. [Google Scholar] [CrossRef]
- Ozma, M.A.; Ghotaslou, R.; Asgharzadeh, M.; Abbasi, A.; Rezaee, M.A.; Kafil, H.S. Cytotoxicity assessment and antimicrobial effects of cell-free supernatants from probiotic lactic acid bacteria and yeast against multidrug-resistant Escherichia coli. Lett. Appl. Microbiol. 2024, 77, ovae084. [Google Scholar] [CrossRef]
- Westmoreland, C.; Holmes, A.M. Assuring consumer safety without animals: Applications for tissue engineering. Organogenesis 2009, 5, 67–72. [Google Scholar] [CrossRef]
- Chuah, L.O.; Foo, H.L.; Loh, T.C.; Mohammed Alitheen, N.B.; Yeap, S.K.; Abdul Mutalib, N.E.; Abdul Rahim, R.; Yusoff, K. Postbiotic metabolites produced by Lactobacillus plantarum strains exert selective cytotoxicity effects on cancer cells. BMC Complement. Altern. Med. 2019, 19, 114. [Google Scholar] [CrossRef]
- Nandhra, G.K.; Mark, E.B.; Di Tanna, G.L.; Haase, A.; Poulsen, J.; Christodoulides, S.; Kung, V.; Klinge, M.W.; Knudsen, K.; Borghammer, P.; et al. Normative values for region-specific colonic and gastrointestinal transit times in 111 healthy volunteers using the 3D-Transit electromagnet tracking system: Influence of age, gender, and body mass index. Neurogastroenterol. Motil. 2020, 32, e13734. [Google Scholar] [CrossRef]
- Procházková, N.; Falony, G.; Dragsted, L.O.; Licht, T.R.; Raes, J.; Roager, H.M. Advancing human gut microbiota research by considering gut transit time. Gut 2023, 72, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Roma, D.; Cecchini, M.E.; Tonini, M.P.; Capella, V.; Aiassa, D.; Rodriguez, N.; Mañas, F. Toxicity assessment and DNA repair kinetics in HEK293 cells exposed to environmentally relevant concentrations of Glyphosate (Roundup® Control Max). Toxicol. Res. 2023, 12, 970–978. [Google Scholar] [CrossRef]
- Tan, H.K.; Foo, H.L.; Loh, T.C.; Banu, N.; Alitheen, M.; Rahim, A.R. Cytotoxic effect of proteinaceous postbiotic metabolites produced by Lactobacillus plantarum I-UL4 cultivated in different media composition on MCF-7 breast cancer cell. Malays. J. Microbiol. 2015, 11, 207–214. [Google Scholar]
- Thirabunyanon, M.; Boonprasom, P.; Niamsup, P. Probiotic potential of lactic acid bacteria isolated from fermented dairy milks on antiproliferation of colon cancer cells. Biotechnol. Lett. 2009, 31, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas-Escudero, J.; Galán-Madruga, D.; Cáceres, J.O. Rapid, reliable and easy-to-perform chemometric-less method for rice syrup adulterated honey detection using FTIR-ATR. Talanta 2023, 253, 123961. [Google Scholar] [CrossRef]
- Arefi, A.; Sturm, B.; Babor, M.; Horf, M.; Hoffmann, T.; Höhne, M.; Friedrich, K.; Schroedter, L.; Venus, J.; Olszewska-Widdrat, A. Digital model of biochemical reactions in lactic acid bacterial fermentation of simple glucose and biowaste substrates. Heliyon 2024, 10, e38791. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Wang, G.; Zhang, Q.; Liu, X.; Gu, Z.; Zhang, H.; Chen, Y.Q.; Chen, W. Determining antioxidant activities of lactobacilli cell-free supernatants by cellular antioxidant assay: A comparison with traditional methods. PLoS ONE 2015, 10, e0119058. [Google Scholar] [CrossRef]
- Lee, J.; Hwang, K.T.; Heo, M.S.; Lee, J.H.; Park, K.Y. Resistance of Lactobacillus plantarum KCTC 3099 from Kimchi to oxidative stress. J. Med. Food 2005, 8, 299–304. [Google Scholar] [CrossRef]
- Gu, X.; Wang, H.; Wang, L.; Zhang, K.; Tian, Y.; Wang, X.; Xu, G.; Guo, Z.; Ahmad, S.; Egide, H.; et al. The antioxidant activity and metabolomic analysis of the supernatant of Streptococcus alactolyticus strain FGM. Sci. Rep. 2024, 14, 8413. [Google Scholar] [CrossRef]
- Xiang, Y.; Li, X.; Zheng, H.; Chen, J.Y.; Lin, L.B.; Zhang, Q.L. Purification and antibacterial properties of a novel bacteriocin against Escherichia coli from Bacillus subtilis isolated from blueberry ferments. LWT-Food Sci. Technol. 2021, 146, 111456. [Google Scholar] [CrossRef]
- Yasir, M.; Dutta, D.; Willcox, M.D.P. Activity of antimicrobial peptides and ciprofloxacin against Pseudomonas aeruginosa biofilms. Molecules 2020, 25, 3843. [Google Scholar] [CrossRef]
- Ge, J.; Sun, Y.; Xin, X.; Wang, Y.; Ping, W. Purification and partial characterization of a novel bacteriocin synthesized by Lactobacillus paracasei HD1-7 isolated from Chinese sauerkraut juice. Sci. Rep. 2016, 6, 19366. [Google Scholar] [CrossRef] [PubMed]
- Alemka, A.; Clyne, M.; Shanahan, F.; Tompkins, T.; Corcionivoschi, N.; Bourke, B. Probiotic colonization of the adherent mucus layer of HT29MTXE12 cells attenuates Campylobacter jejuni virulence properties. Infect. Immun. 2010, 78, 2812–2822. [Google Scholar] [CrossRef]
- van Meerloo, J.; Kaspers, G.J.; Cloos, J. Cell sensitivity assays: The MTT assay. Methods Mol. Biol. 2011, 731, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Chan, F.K.; Moriwaki, K.; De Rosa, M.J. Detection of necrosis by release of lactate dehydrogenase activity. Methods Mol. Biol. 2013, 979, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Marinas, I.C.; Ignat, L.; Maurușa, I.E.; Gaboreanu, M.D.; Adina, C.; Popa, M.; Chifiriuc, M.C.; Angheloiu, M.; Georgescu, M.; Iacobescu, A.; et al. Insights into the physico-chemical and biological characterization of sodium lignosulfonate—Silver nanosystems designed for wound management. Heliyon 2024, 10, e26047. [Google Scholar] [CrossRef]
- Madhu, G.; Bose, V.C.; Aiswaryaraj, A.S.; Maniammal, K.; Biju, V. Defect dependent antioxidant activity of nanostructured nickel oxide synthesized through a novel chemical method. Colloids Surf. Physicochem. Eng. Asp. 2013, 429, 44–50. [Google Scholar] [CrossRef]
- Celik, S.E.; Ozyürek, M.; Güçlü, K.; Apak, R. Determination of antioxidants by a novel on-line HPLC-cupric reducing antioxidant capacity (CUPRAC) assay with post-column detection. Anal. Chim. Acta 2010, 674, 79–88. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Methods Enzymol. 1999, 299, 15–27. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molina, D.; Marinas, I.C.; Angamarca, E.; Hanganu, A.; Stan, M.; Chifiriuc, M.C.; Tenea, G.N. Postbiotic-Based Extracts from Native Probiotic Strains: A Promising Strategy for Food Preservation and Antimicrobial Defense. Antibiotics 2025, 14, 318. https://doi.org/10.3390/antibiotics14030318
Molina D, Marinas IC, Angamarca E, Hanganu A, Stan M, Chifiriuc MC, Tenea GN. Postbiotic-Based Extracts from Native Probiotic Strains: A Promising Strategy for Food Preservation and Antimicrobial Defense. Antibiotics. 2025; 14(3):318. https://doi.org/10.3390/antibiotics14030318
Chicago/Turabian StyleMolina, Diana, Ioana C. Marinas, Evelyn Angamarca, Anamaria Hanganu, Miruna Stan, Mariana C. Chifiriuc, and Gabriela N. Tenea. 2025. "Postbiotic-Based Extracts from Native Probiotic Strains: A Promising Strategy for Food Preservation and Antimicrobial Defense" Antibiotics 14, no. 3: 318. https://doi.org/10.3390/antibiotics14030318
APA StyleMolina, D., Marinas, I. C., Angamarca, E., Hanganu, A., Stan, M., Chifiriuc, M. C., & Tenea, G. N. (2025). Postbiotic-Based Extracts from Native Probiotic Strains: A Promising Strategy for Food Preservation and Antimicrobial Defense. Antibiotics, 14(3), 318. https://doi.org/10.3390/antibiotics14030318