

Levels of Different Microbial Groups on Inert Surfaces of Poultry Slaughterhouses: Identification Using Matrix-Assisted Laser Desorption Ionization Time-of-Flight and Detection of Extended-Spectrum Beta-Lactamase- and Carbapenemase-Producing Enterobacteria

 and
and
Abstract

1. Introduction
2. Results
2.1. Microbial Counts
2.2. Isolation and Identification of Microorganisms
2.3. Phenotypic Resistance to Antibiotics
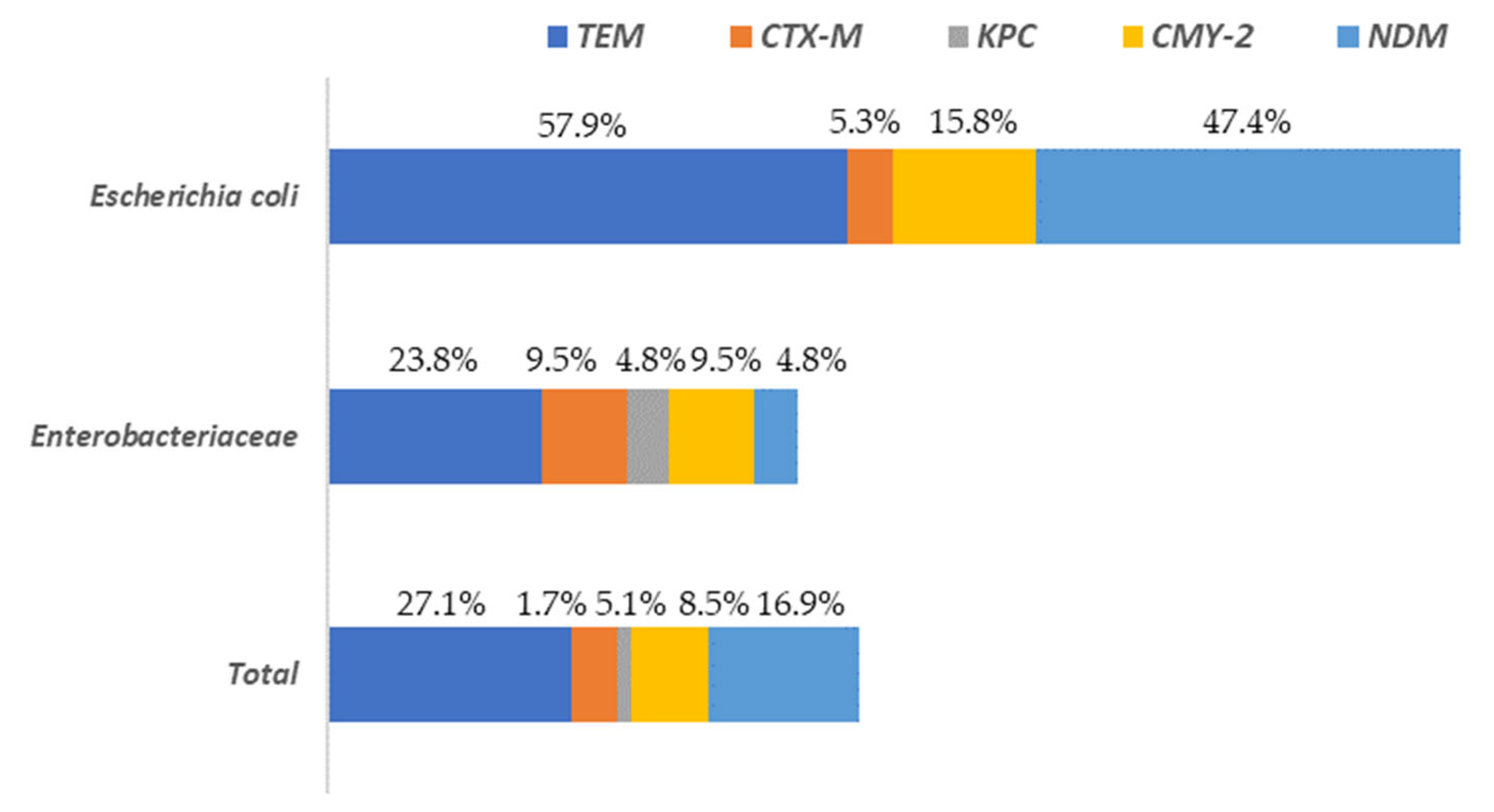
2.4. Detection of Resistance Genes to Beta-Lactam Antibiotics in Enterobacteriaceae
3. Discussion
3.1. Microbial Counts
3.2. Isolation and Identification of Microorganisms
3.3. Phenotypic Resistance to Antibiotics
3.4. Detection of Resistance Genes to Beta-Lactam Antibiotics in Enterobacteriaceae
4. Material and Methods
4.1. Sampling
4.2. Microbiological Analysis
4.3. Identification of Isolates Using MALDI-TOF
4.4. Confirmation of Isolates Using PCR
4.5. Phenotypic Resistance to Antibiotics
4.6. Detection of Resistance Genes to Beta-Lactam Antibiotics in Enterobacteriaceae
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- MAPA. Indicadores Económicos del Sector Avícola de Carne Ministerio de Agricultura, Pesca y Alimentación. 2022. Available online: https://www.mapa.gob.es/es/ganaderia/temas/produccion-y-mercados-ganaderos/indicadorescompleto_tcm30-623991.pdf (accessed on 14 May 2024).
- OECD-FAO. Agricultural Outlook 2023–2032. 2023. Available online: https://www.oecd-ilibrary.org/agriculture-and-food/oecd-fao-agricultural-outlook-2023-2032_08801ab7-en (accessed on 14 May 2024).
- Del Río, E.; Panizo-Morán, M.; Prieto, M.; Alonso-Calleja, C.; Capita, R. Effect of various chemical decontamination treatments on natural microflora and sensory characteristics of poultry. Int. J. Food Microbiol. 2007, 115, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, H.; Saghaian, S.; Boccia, F. Antibiotic-free poultry meat consumption and its determinants. Foods 2023, 12, 1776. [Google Scholar] [CrossRef]
- Ricke, S.C. Impact of prebiotics on poultry production and food safety. Yale J. Biol. Med. 2018, 91, 151. [Google Scholar]
- Śmiałek, M.; Kowalczyk, J.; Koncicki, A. Influence of vaccination of broiler chickens against Escherichia coli with live attenuated vaccine on general properties of E. coli population, IBV vaccination efficiency, and production parameters—A field experiment. Poultry Sci. 2020, 99, 5452–5460. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Diarra, M.S.; Rehman, M.A.; Li, L.; Yu, H.; Yin, X.; Aslam, M.; Carrillo, C.D.; Yang, C.; Gong, J. Virulence potential of antimicrobial-resistant extraintestinal pathogenic Escherichia coli from retail poultry meat in a Caenorhabditis elegans model. J. Food Prot. 2023, 86, 100008. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Ding, X.; Zhao, Q.; Sun, H.; Li, T.; Li, Z.; Wang, H.; Zhang, L.; Zhang, C.; Xu, S. Development of an organic acid compound disinfectant to control food-borne pathogens and its application in chicken slaughterhouses. Poultry Sci. 2022, 101, 101842. [Google Scholar] [CrossRef]
- Rouger, A.; Tresse, O.; Zagorec, M. Bacterial contaminants of poultry meat: Sources, species, and dynamics. Microorganisms 2017, 5, 50. [Google Scholar] [CrossRef]
- Zeng, H.; De Reu, K.; Gabriël, S.; Mattheus, W.; De Zutter, L.; Rasschaert, G. Salmonella prevalence and persistence in industrialized poultry slaughterhouses. Poultry Sci. 2021, 100, 100991. [Google Scholar] [CrossRef] [PubMed]
- Ripolles-Avila, C.; Hascoët, A.S.; Martínez-Suárez, J.V.; Capita, R.; Rodríguez-Jerez, J.J. Evaluation of the microbiological contamination of food processing environments through implementing surface sensors in an Iberian pork processing plant: An approach towards the control of Listeria monocytogenes. Food Control 2019, 99, 40–47. [Google Scholar] [CrossRef]
- Botta, C.; Ferrocino, I.; Pessione, A.; Cocolin, L.; Rantsiou, K. Spatiotemporal distribution of the environmental microbiota in food processing plants as impacted by cleaning and sanitizing procedures: The case of slaughterhouses and gaseous ozone. Appl. Environ. Microbiol. 2020, 86, e01861-20. [Google Scholar] [CrossRef]
- Song, X.; Wang, H.; Xu, X. Investigation of microbial contamination in a chicken slaughterhouse environment. J. Food Sci. 2021, 86, 3598–3610. [Google Scholar] [CrossRef] [PubMed]
- Abreu, R.; Semedo-Lemsaddek, T.; Cunha, E.; Tavares, L.; Oliveira, M. Antimicrobial drug resistance in poultry production: Current status and innovative strategies for bacterial control. Microorganisms 2023, 11, 953. [Google Scholar] [CrossRef] [PubMed]
- Capita, R.; Alonso-Calleja, C. Antibiotic-resistant bacteria: A challenge for the food industry. Crit. Rev. Food Sci. Nutr. 2013, 53, 11–48. [Google Scholar] [CrossRef] [PubMed]
- Yengui, M.; Trabelsi, R.; Khannous, L.; Mathlouthi, N.E.; Adnan, M.; Siddiqui, A.J.; Noumi, E.; Snoussi, M.; Gdoura, R. Rapid detection of beta-lactamases genes among Enterobacterales in urine samples by using real-time PCR. BioMed. Res. Int. 2022, 2022, 8612933. [Google Scholar] [CrossRef] [PubMed]
- Maciuca, I.E.; Williams, N.J.; Tuchilus, C.; Dorneanu, O.; Guguianu, E.; Carp-Carare, C.; Rimbu, C.; Timofte, D. High prevalence of Escherichia coli-producing CTX-M-15 extended-spectrum beta-lactamases in poultry and human clinical isolates in Romania. Microb. Drug Resist. 2015, 21, 651–662. [Google Scholar] [CrossRef]
- Silva, N.; Carvalho, I.; Currie, C.; Sousa, M.; Igrejas, G.; Poeta, P. Extended-Spectrum-β-Lactamase and Carbapenemase-Producing Enterobacteriaceae in Food-Producing Animals in Europe: An Impact on Public Health? In Antibiotic Drug Resistance; Capelo-Martínez, J.L., Igrejas, G., Eds.; John Wiley & Sons Inc.: Hoboken, NY, USA, 2019; pp. 261–273. [Google Scholar]
- Pérez-Vázquez, M.; Sola Campoy, P.J.; Ortega, A.; Bautista, V.; Monzón, S.; Ruiz-Carrascoso, G.; Mingorance, J.; González-Barberá, E.M.; Gimeno, C.; Aracil, B.; et al. Emergence of NDM-producing Klebsiella pneumoniae and Escherichia coli in Spain: Phylogeny, resistome, virulence and plasmids encoding blaNDM-like genes as determined by WGS. J. Antimicrob. Chemother. 2019, 74, 3489–3496. [Google Scholar] [CrossRef] [PubMed]
- Galán-Sánchez, F.; Aznar-Marín, P.; Marín-Casanova, P.; Rodríguez-Iglesias, M. Diversity of bla genes and low incidence of CTX-M in plasmid-mediated AmpC-producing Escherichia coli clinical isolates. APMIS 2014, 122, 796–799. [Google Scholar] [CrossRef] [PubMed]
- Lopes, E.; Saavedra, M.J.; Costa, E.; de Lencastre, H.; Poirel, L.; Aires-de-Sousa, M. Epidemiology of carbapenemase-producing Klebsiella pneumoniae in northern Portugal: Predominance of KPC-2 and OXA-J. Global Antimicrob. Resist. 2020, 22, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Delshadi, R.; Bahrami, A.; Assadpour, E.; Williams, L.; Jafari, S.M. Nano/microencapsulated natural antimicrobials to control the spoilage microorganisms and pathogens in different food products. Food Control 2021, 128, 108180. [Google Scholar] [CrossRef]
- Møretrø, T.; Langsrud, S. Residential bacteria on surfaces in the food industry and their implications for food safety and quality. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1022–1041. [Google Scholar] [CrossRef]
- Ferone, M.; Gowen, A.; Fanning, S.; Scannell, A.G. Microbial detection and identification methods: Bench top assays to omics approaches. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3106–3129. [Google Scholar] [CrossRef] [PubMed]
- Uzuriaga, M.; Leiva, J.; Guillén-Grima, F.; Rua, M.; Yuste, J.R. Clinical impact of rapid bacterial microbiological identification with the MALDI-TOF MS. Antibiotics 2023, 12, 1660. [Google Scholar] [CrossRef] [PubMed]
- Whyte, P.J.D.C.; Collins, J.D.; McGill, K.; Monahan, C.; O’mahony, H. Distribution and prevalence of airborne microorganisms in three commercial poultry processing plants. J. Food Prot. 2001, 64, 388–391. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Calleja, C.; Guerrero-Ramos, E.; Capita, R. Hygienic status assessment of two lamb slaughterhouses in Spain. J. Food Prot. 2017, 80, 1152–1158. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Takahashi, H.; Kondo, A.; Koike, F.; Kuda, T.; Kimura, B.; Kobayashi, M. Distribution of psychrophilic microorganisms in a beef slaughterhouse in Japan after cleaning. PLoS ONE 2022, 17, e0268411. [Google Scholar] [CrossRef] [PubMed]
- Nortje, G.L.; Nel, L.; Jordaan, E.; Badenhorst, K.; Goedhart, G.; Holzapfel, W.H.; Grimbeek, R.J. A quantitative survey of a meat production chain to determine the microbial profile of the final product. J. Food Prot. 1990, 53, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Gribble, A.; Mills, J.; Brightwell, G. The spoilage characteristics of Brochothrix thermosphacta and two psychrotolerant Enterobacteriacae in vacuum packed lamb and the comparison between high and low pH cuts. Meat Sci. 2014, 97, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Bruzaroski, S.R.; de Souza, R.P.; da Silva Pasquim, P.; Fagnani, R.; de Santana, E.H.W. Influence of storage temperature on the population of microorganisms in raw sheep milk and its physical-chemical profile. Res. Soc. Dev. 2020, 9, e27691210796. [Google Scholar] [CrossRef]
- Haleem, A.M.; Al-bakri, S.A.; Al-Hiyaly, S.A. Determination of microbial content in poultry meat in local Iraqi markets. J. Microbiol. Res. 2013, 3, 205–207. [Google Scholar]
- Hu, Z.; Peng, Z.; Zhang, X.; Li, Z.; Jia, C.; Li, X.; Lv, Y.; Tan, C.; Chen, H.; Wang, X. Prevalence and molecular characterization of antimicrobial-resistant Escherichia coli in pig farms, slaughterhouses, and terminal markets in Henan province of China. Foodborne Path. Dis. 2021, 18, 733–743. [Google Scholar] [CrossRef]
- Vázquez-Villanueva, J.; Vázquez, K.; Martínez-Vázquez, A.V.; Wong-González, A.; Hernández-Escareño, J.; Cabrero-Martínez, O.; Cruz-Pulido, W.L.; Guerrero, A.; Rivera, G.; Bocanegra-García, V. Molecular and antimicrobial susceptibility characterization of Escherichia coli isolates from bovine slaughterhouse process. Antibiotics 2023, 12, 291. [Google Scholar] [CrossRef]
- Lavilla Lerma, L.; Benomar, N.; Gálvez, A.; Abriouel, H. Prevalence of bacteria resistant to antibiotics and/or biocides on meat processing plant surfaces throughout meat chain production. Int. J. Food Microbiol. 2013, 161, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Sebsibe, M.A.; Asfaw, E.T. Occurrence of multi-drug resistant Escherichia coli and Escherichia coli O157: H7 in meat and swab samples of various contact surfaces at abattoir and butcher shops in Jimma town, Southwest district of Ethiopia. Inf. Drug Res. 2020, 13, 3853–3862. [Google Scholar] [CrossRef]
- Agostinho Davanzo, E.F.; Dos Santos, R.L.; Castro, V.H.L.; Palma, J.M.; Pribul, B.R.; Dallago, B.S.L.; Fuga, B.; Medeiros, M.; Titze de Almeida, S.S.; da Costa, H.M.B.; et al. Molecular characterization of Salmonella spp. and Listeria monocytogenes strains from biofilms in cattle and poultry slaughterhouses located in the federal District and State of Goiás, Brazil. PLoS ONE 2021, 16, e0259687. [Google Scholar] [CrossRef]
- Stadtlober, G.A.W.; Fiorentini, Â.M.; Severo, J.; Bernardo, K.B.D.; Carvalho, I.R.; Loro, M.V. Contamination by aerobic mesophilal and Enterobacteriaceae bacteria in a pig refrigerator. Agron. Sci. Biotechnol. 2024, 10, 1–13. [Google Scholar] [CrossRef]
- Moura-Alves, M.; Carvalho, M.; Ribeiro, D.H.B.; Barbosa, J.; Silveira, L.; Pista, Â.; Pinto, H.P.; Saraiva, C.; Teixeira, P.; Esteves, A. Hygiene indicators and salmonellae on surfaces of swine carcasses from two slaughterhouses in northern Portugal. J. Food Prot. 2022, 85, 1566–1575. [Google Scholar] [CrossRef]
- Soares-Santos, V.; Barreto, A.S.; Semedo-Lemsaddek, T. Characterization of enterococci from food and food-related settings. J. Food Prot. 2015, 78, 1320–1326. [Google Scholar] [CrossRef] [PubMed]
- Wambui, J.; Tasara, T.; Njage, P.M.K.; Stephan, R. Species distribution and antimicrobial profiles of Enterococcus spp. isolates from Kenyan small and medium enterprise slaughterhouses. J. Food Prot. 2018, 81, 1445–1449. [Google Scholar] [CrossRef]
- Guerrero-Ramos, E.; Cordero, J.; Molina-González, D.; Poeta, P.; Igrejas, G.; Alonso-Calleja, C.; Capita, R. Antimicrobial resistance and virulence genes in enterococci from wild game meat in Spain. Food Microbiol. 2016, 53, 156–164. [Google Scholar] [CrossRef]
- Guerrero-Ramos, E.; Molina-Gonzalez, D.; Blanco-Moran, S.; Igrejas, G.; Poeta, P.; Alonso-Calleja, C.; Capita, R. Prevalence, antimicrobial resistance, and genotypic characterization of vancomycin-resistant enterococci in meat preparations. J. Food Prot. 2016, 79, 748–756. [Google Scholar] [CrossRef]
- Jang, Y.S.; Moon, J.S.; Kang, H.J.; Bae, D.; Seo, K.H. Prevalence, characterization, and antimicrobial susceptibility of Listeria monocytogenes from raw beef and slaughterhouse environments in Korea. Foodborne Path. Dis. 2021, 18, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Gowda, T.K.; Van Damme, I. Occurrence and antibiotic susceptibility of Listeria species and Staphylococcus aureus in cattle slaughterhouses of Kerala, South India. Foodborne Path. Dis. 2017, 14, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Tasci, F.; Sudagidan, M.; Yavuz, O.; Soyucok, A.; Aydin, A. Virulence properties of Listeria monocytogenes isolated from meat and meat contact surfaces in a slaughterhouse. Pol. J. Vet. Sci. 2024, 27, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Gregova, G.; Kmetova, M.; Kmet, V.; Venglovsky, J.; Feher, A. Antibiotic resistance of Escherichia coli isolated from a poultry slaughterhouse. Ann. Agric. Environ. Med. 2012, 19, 75–79. [Google Scholar] [PubMed]
- Elabbasy, M.T.; Hussein, M.A.; Algahtani, F.D.; Abd El-Rahman, G.I.; Morshdy, A.E.; Elkafrawy, I.A.; Adeboye, A.A. MALDI-TOF MS based typing for rapid screening of multiple antibiotic resistance E. coli and virulent non-O157 shiga toxin-producing E. coli isolated from the slaughterhouse settings and beef carcasses. Foods 2021, 10, 820. [Google Scholar] [CrossRef] [PubMed]
- Savin, M.; Bierbaum, G.; Mutters, N.T.; Schmithausen, R.M.; Kreyenschmidt, J.; García-Meniño, I.; Schmoger, S.; Käsbohrer, A.; Hammerl, J.A. Genetic characterization of carbapenem-resistant Klebsiella spp. from municipal and slaughterhouse wastewater. Antibiotics 2022, 11, 435. [Google Scholar] [CrossRef] [PubMed]
- Homeier-Bachmann, T.; Heiden, S.E.; Lübcke, P.K.; Bachmann, L.; Bohnert, J.A.; Zimmermann, D.; Schaufler, K. Antibiotic-resistant Enterobacteriaceae in wastewater of abattoirs. Antibiotics 2021, 10, 568. [Google Scholar] [CrossRef] [PubMed]
- WHO (World Health Organization). Critically Important Antimicrobials for Human Medicine, 6th ed.; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- OIE (World Organization for Animal Health). OIE List of Antimicrobial Agents of Veterinary Importance; World Organization for Animal Health: Paris, France, 2018. [Google Scholar]
- Nordmann, P.; Poirel, L.; Walsh, T.R.; Livermore, D.M. The emerging NDM carbapenemases. Trends Microbiol. 2011, 19, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.D.; Perez-Bonilla, D.; Hallowell, A.; Redding, L.E. Carbapenem prescribing at a veterinary teaching hospital before an outbreak of carbapenem-resistant Escherichia coli. J. Small Anim. Pract. 2022, 63, 442–446. [Google Scholar] [CrossRef]
- Jiménez-Belenguer, A.I.; Ferrús, M.A.; Hernández, M.; García-Hernández, J.; Moreno, Y.; Castillo, M.Á. Prevalence and characterization of Beta-lactam and Carbapenem-resistant bacteria isolated from organic fresh produce retailed in eastern Spain. Antibiotics 2023, 12, 387. [Google Scholar] [CrossRef]
- Friese, A.; Schulz, J.; Laube, H.; von Salviati, C.; Hartung, J.; Roesler, U. Faecal occurrence and emissions of livestock-associated methicillin-resistant Staphylococcus aureus (laMRSA) and ESbl/AmpC-producing E. coli from animal farms in Germany. Berl. Munch. Tierarztl. Wochenschr. 2013, 126, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Li, Y.; Yang, Y.; Shen, Z.; Cai, C.; Wang, Y.; Wang, S. High prevalence and persistence of carbapenem and colistin resistance in livestock farm environments in China. J. Hazard. Mat. 2021, 406, 124298. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.S.; Choi, D.S.; Kim, Y.J.; Chon, J.W.; Kim, H.S.; Park, H.J.; Moon, J.S.; Wee, S.H.; Seo, K.H. Characterization of Escherichia coli–producing extended-spectrum β-lactamase (ESBL) isolated from chicken slaughterhouses in South Korea. Foodborne Pathog. Dis. 2015, 12, 741–748. [Google Scholar] [CrossRef]
- Wei, B.; Shang, K.; Cha, S.Y.; Zhang, J.F.; Jang, H.K.; Kang, M. Conjugative plasmid-mediated extended spectrum cephalosporin resistance in genetically diverse Escherichia coli from a chicken slaughterhouse. Animals 2021, 11, 2491. [Google Scholar] [CrossRef] [PubMed]
- Adel, W.A.; Ahmed, A.M.; Hegazy, Y.; Torky, H.A.; Shimamoto, T. High prevalence of ESBL and plasmid-mediated quinolone resistance genes in Salmonella enterica isolated from retail meats and slaughterhouses in Egypt. Antibiotics 2021, 10, 881. [Google Scholar] [CrossRef] [PubMed]
- Clemente, L.; Leão, C.; Moura, L.; Albuquerque, T.; Amaro, A. Prevalence and characterization of ESBL/AmpC producing Escherichia coli from fresh meat in Portugal. Antibiotics 2021, 10, 1333. [Google Scholar] [CrossRef] [PubMed]
- Economou, V.; Delis, G.; Stavrou, D.; Gousia, P.; Tsitsos, A.; Mantzios, T.; Choiliara, E.; Kolovos, N.; Soultos, N. Characterization of extended spectrum cephalosporin-resistant Escherichia coli strains isolated from raw poultry carcasses in catering services in Northern Greece. Vet. Sci. 2023, 10, 487. [Google Scholar] [CrossRef] [PubMed]
- Savin, M.; Bierbaum, G.; Schmithausen, R.M.; Heinemann, C.; Kreyenschmidt, J.; Schmoger, S.; Akbaba, I.; Käsbohrer, A.; Hammerl, J.A. Slaughterhouse wastewater as a reservoir for extended-spectrum β-lactamase (ESBL)-producing, and colistin-resistant Klebsiella spp. and their impact in a “One Health” perspective. Sci. Total Environ. 2022, 804, 150000. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Laorden, A.; Arraiz-Fernández, C.; González-Fandos, E. Microbiological quality and safety of fresh turkey meat at retail level, including the presence of ESBL-producing Enterobacteriaceae and methicillin-resistant S. aureus. Foods 2023, 12, 1274. [Google Scholar] [CrossRef]
- Gundran, R.S.; Cardenio, P.A.; Villanueva, M.A.; Sison, F.B.; Benigno, C.C.; Kreausukon, K.; Pichpol, D.; Punyapornwithaya, V. Prevalence and distribution of blaCTX-M, blaSHV, blaTEM genes in extended-spectrum β-lactamase-producing E. coli isolates from broiler farms in the Philippines. BMC Vet. Res. 2019, 15, 227. [Google Scholar] [CrossRef]
- Patil, S.; Chen, X.; Wen, F. Exploring the phenotype and genotype of multi-drug resistant Klebsiella pneumoniae harbouring blaCTX-M group extended-spectrum β-lactamases recovered from paediatric clinical cases in Shenzhen, China. Ann. Clin. Microbiol. Antimicrob. 2019, 18, 32. [Google Scholar] [CrossRef] [PubMed]
- Saliu, E.M.; Vahjen, W.; Zentek, J. Types and prevalence of extended–spectrum beta–lactamase producing Enterobacteriaceae in poultry. Animal Health Res. Rev. 2017, 18, 46–57. [Google Scholar] [CrossRef]
- Akpaka, P.E.; Vaillant, A.; Wilson, C.; Jayaratne, P. Extended spectrum beta-lactamase (ESBL) produced by gram-negative bacteria in Trinidad and Tobago. Int. J. Microbiol. 2021, 2021, 5582755. [Google Scholar] [CrossRef] [PubMed]
- Hasman, H.; Mevius, D.; Veldman, K.; Olesen, I.; Aarestrup, F.M. β-Lactamases among extended-spectrum β-lactamase (ESBL)-resistant Salmonella from poultry, poultry products and human patients in The Netherlands. J. Antimicrob. Chem. 2005, 56, 115–121. [Google Scholar] [CrossRef]
- Savin, M.; Alexander, J.; Bierbaum, G.; Hammerl, J.A.; Hembach, N.; Schwartz, T.; Schimthausen, R.M.; Sib, E.; Voigt, A.; Kreyenschmidt, J. Antibiotic-resistant bacteria, antibiotic resistance genes, and antibiotic residues in wastewater from a poultry slaughterhouse after conventional and advanced treatments. Sci. Rep. 2021, 11, 16622. [Google Scholar] [CrossRef] [PubMed]
- Microbiology of the Food Chain—Horizontal Method for the Detection and Enumeration of Listeria Monocytogenes and of Listeria spp.—Part 1: Detection Method. UNE-EN ISO 11290-1:2018. Available online: https://www.une.org/encuentra-tu-norma/busca-tu-norma/norma/?c=N0059546 (accessed on 12 May 2024).
- Microbiology of the Food Chain—Horizontal Method for the Detection, Enumeration and Serotyping of Salmonella—Part 1: Detection of Salmonella spp. UNE-EN ISO 6579-1:2017. Available online: https://www.une.org/encuentra-tu-norma/busca-tu-norma/norma/?c=N0058760 (accessed on 12 May 2024).
- Jay, J.M. A review of aerobic and psychrotrophic plate count procedures for fresh meat and poultry products. J. Food Prot. 2002, 65, 1200–1206. [Google Scholar] [CrossRef] [PubMed]
- Cousin, M.A.; Jay, J.M.; Vasavada, P.C. Psychrotrophic microorganisms. In Compendium of Methods for the Microbiological Examination of Foods, 4th ed.; Downes, F.P., Ito, K., Eds.; American Public Health Association: Washington, DC, USA, 2001; pp. 159–166. [Google Scholar]
- Baird, R.M.; Corry, J.E.L.; Curtis, G.D.W. Pharmacopeia of culture media for food microbiology. Int. J. Food Microbiol. 1987, 5, 201–266. [Google Scholar]
- Sørum, M.; Holstad, G.; Lillehaug, A.; Kruse, H. Prevalence of vancomycin resistant enterococci on poultry farms established after the ban of avoparcin. Avian Dis. 2004, 48, 823–828. [Google Scholar] [CrossRef] [PubMed]
- Panera-Martínez, S.; Rodríguez-Melcón, C.; Serrano-Galán, V.; Alonso-Calleja, C.; Capita, R. Prevalence, quantification and antibiotic resistance of Listeria monocytogenes in poultry preparations. Food Control 2021, 135, 108608. [Google Scholar] [CrossRef]
- Fazzeli, H.; Arabestani, M.R.; Esfahani, B.N.; Khorvash, F.; Pourshafie, M.R.; Moghim, S.; Safaei, H.G.; Faghri, J.; Narimani, T. Development of PCR-based method for detection of Enterobacteriaceae in septicemia. J. Res. Med. Sci. 2012, 17, 671–675. [Google Scholar]
- El-Sayed, A.K.A.; Abou Dobara, M.; El-Shihy, E.S. Simultaneous detection of seven foodborne Enterobacteriaceae pathogens using multiplex PCR. J. Egypt. Acad. Soc. Environ. Dev. 2019, 20, 61–78. [Google Scholar] [CrossRef]
- Deasy, B.M.; Rea, M.C.; Fitzgerald, G.F.; Cogan, T.M.; Beresford, T.P. A rapid PCR based method to distinguish between Lactococcus and Enterococcus. Sys. Appl. Microbiol. 2000, 23, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.; Park, S.H.; Yeom, Y.S.; Shrivastav, A.; Lee, S.H.; Kim, Y.R.; Kim, H.Y. Simultaneous detection of Listeria species isolated from meat processed foods using multiplex PCR. Food Control 2013, 32, 659–664. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; Clinical and Laboratory Standars Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Weiß, D.; Engelmann, I.; Braun, S.D.; Monecke, S.; Ehricht, R. A multiplex real-time PCR for the direct, fast, economic and simultaneous detection of the carbapenemase genes blaKPC, blaNDM, blaVIM and blaOXA. J. Microbiol. Meth. 2017, 142, 20–26. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development Environment for R. 2019. Available online: http://www.rstudio.com/ (accessed on 15 March 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | TAC | Psychrotrophs | Enterobacteria | Coliforms | Enterococci | VRE |
|---|---|---|---|---|---|---|
| A1 | 4.25 × 101 | 3.33 × 101 | 1.22 × 100 | 5.47 × 10−1 | 1.50 × 10−1 | 1.13 × 10−1 |
| A2 | 4.18 × 102 | 3.36 × 102 | 1.14 × 10−1 | 9.37 × 10−1 | 3.12 × 10−1 | <5.20 × 10−2 |
| A3 | 1.41 × 102 | 8.49 × 101 | <3.92 × 10−3 | <3.92 × 10−3 | 7.84 × 10−3 | <1.96 × 10−2 |
| A4 | 9.55 × 102 | 2.51 × 102 | <1.25 × 10−2 | <1.25 × 10−2 | <1.25 × 10−2 | <6.25 × 10−2 |
| A5 | 1.74 × 100 | 1.61 × 10−1 | <1.46 × 10−3 | <1.46 × 10−3 | <1.46 × 10−3 | <7.31 × 10−3 |
| A6 | 2.38 × 103 | 1.34 × 103 | <5.70 × 10−2 | 1.14 × 10−1 | 1.88 × 100 | <2.85 × 10−1 |
| A7 | 2.12 × 102 | 1.90 × 102 | 7.47 × 10−1 | 1.68 × 10−2 | 7.47 × 10−2 | 4.67 × 10−2 |
| A8 | 3.12 × 101 | 1.63 × 101 | <2.21 × 10−3 | <2.21 × 10−3 | <2.21 × 10−3 | <1.11 × 10−2 |
| A9 | 9.07 × 101 | 5.76 × 101 | <5.71 × 10−3 | <5.71 × 10−3 | <5.71 × 10−3 | <2.86 × 10−2 |
| A10 | 4.80 × 101 | 3.66 × 101 | 1.57 × 10−3 | <1.57 × 10−3 | <1.57 × 10−3 | <7.86 × 10−3 |
| A11 | 4.94 × 100 | 1.70 × 100 | <7.72 × 10−3 | <7.72 × 10−3 | 7.72 × 10−3 | 2.16 × 100 |
| A12 | 1.16 × 102 | 9.31 × 101 | 1.29 × 101 | 1.73 × 100 | 2.67 × 10−2 | <3.33 × 10−2 |
| A13 | 1.58 × 101 | 9.19 × 100 | 3.23 × 10−2 | <4.04 × 10−3 | <4.04 × 10−3 | <2.02 × 10−2 |
| A14 | 4.29 × 102 | 2.91 × 102 | 2.10 × 10−1 | <3.62 × 10−3 | <3.62 × 10−3 | <1.81 × 10−2 |
| A15 | 1.60 × 101 | 1.28 × 101 | <3.43 × 10−3 | <3.43 × 10−3 | <3.43 × 10−3 | <1.72 × 10−2 |
| A16 | 1.00 × 101 | 6.00 × 100 | <2.58 × 10−3 | <2.58 × 10−3 | <2.58 × 10−3 | <1.29 × 10−2 |
| A17 | 9.79 × 101 | 1.93 × 102 | 5.84 × 100 | <4.87 × 10−3 | <4.87 × 10−3 | <2.43 × 10−2 |
| A18 | 1.72 × 101 | 2.96 × 100 | <3.69 × 10−3 | <3.69 × 10−3 | <3.69 × 10−3 | <1.85 × 10−2 |
| A19 | 4.17 × 102 | 4.59 × 102 | <3.06 × 10−2 | <3.06 × 10−2 | <3.06 × 10−2 | <1.53 × 10−1 |
| A20 | 3.87 × 101 | 3.24 × 101 | 2.01 × 10−1 | 3.92 × 10−2 | <4.90 × 10−3 | <2.45 × 10−2 |
| A21 | 3.45 × 102 | 1.05 × 102 | 3.30 × 100 | <2.82 × 10−3 | 2.82 × 10−3 | <1.41 × 10−2 |
| A22 | 5.48 × 102 | 1.16 × 103 | 1.95 × 101 | <5.17 × 10−3 | 1.55 × 10−2 | <2.58 × 10−2 |
| B1 | 7.49 × 101 | 1.33 × 101 | <1.02 × 10−2 | <1.02 × 10−2 | <1.02 × 10−2 | <5.12 × 10−2 |
| B2 | 2.44 × 102 | 8.95 × 101 | 1.06 × 10−2 | 3.17 × 10−2 | <4.23 × 10−2 | <5.29 × 10−2 |
| B3 | 5.23 × 101 | 1.94 × 101 | <6.76 × 10−3 | <6.76 × 10−3 | <6.76 × 10−3 | <3.38 × 10−2 |
| B4 | 4.70 × 102 | 1.95 × 102 | <1.29 × 10−2 | <1.29 × 10−2 | <3.87 × 10−2 | <6.45 × 10−2 |
| B5 | 6.14 × 100 | 5.39 × 10−1 | 1.46 × 10−3 | <1.46 × 10−3 | <1.46 × 10−3 | <7.28 × 10−3 |
| B6 | 5.05 × 102 | 2.48 × 101 | 7.07 × 10−1 | 1.06 × 100 | <2.65 × 10−1 | <4.42 × 10−1 |
| B7 | 6.21 × 100 | 6.28 × 10−1 | <4.57 × 10−3 | <4.57 × 10−3 | <4.57 × 10−3 | <2.28 × 10−2 |
| B8 | 4.11 × 100 | 4.28 × 100 | <4.57 × 10−3 | <4.57 × 10−3 | <4.57 × 10−3 | <2.28 × 10−2 |
| B9 | 8.70 × 101 | 1.71 × 10−1 | <5.71 × 10−3 | <5.71 × 10−3 | <5.71 × 10−3 | <2.85 × 10−2 |
| B10 | 1.18 × 102 | 6.18 × 101 | 4.01 × 10−1 | 2.13 × 10−2 | <3.04 × 10−3 | <1.52 × 10−2 |
| B11 | 1.27 × 103 | 2.90 × 103 | 8.91 × 101 | 1.69 × 102 | 4.88 × 100 | 5.71 × 10−1 |
| B12 | 1.43 × 101 | 8.84 × 100 | <3.24 × 10−2 | <3.24 × 10−2 | <3.24 × 10−2 | <1.62 × 10−1 |
| B13 | 8.97 × 102 | 5.72 × 102 | <1.20 × 10−2 | <1.20 × 10−2 | <1.20 × 10−2 | <5.98 × 10−2 |
| B14 | 1.80 × 102 | 9.91 × 101 | 1.63 × 100 | 4.18 × 10−1 | <9.09 × 10−3 | <4.55 × 10−2 |
| B15 | 1.91 × 101 | 1.75 × 101 | 4.56 × 10−2 | 9.12 × 10−3 | <3.04 × 10−3 | <1.52 × 10−2 |
| B16 | 8.76 × 101 | 3.45 × 10−2 | <6.90 × 10−3 | <6.90 × 10−3 | <6.90 × 10−3 | <3.45 × 10−2 |
| B17 | 7.59 × 101 | 1.93 × 10−1 | <5.14 × 10−3 | <5.14 × 10−3 | <5.14 × 10−3 | <2.57 × 10−2 |
| B18 | 3.60 × 101 | 1.81 × 101 | 2.27 × 10−2 | 5.67 × 10−3 | <5.67 × 10−3 | <2.83 × 10−2 |
| B19 | 1.46 × 101 | 7.31 × 10−2 | <2.66 × 10−3 | <2.66 × 10−3 | <2.66 × 10−3 | <1.33 × 10−2 |
| B20 | 2.43 × 100 | 1.91 × 10−1 | 6.93 × 10−3 | 2.77 × 10−2 | 6.93 × 10−3 | 3.47 × 10−2 |
| B21 | 1.16 × 100 | 2.00 × 10−1 | <7.26 × 10−3 | <7.26 × 10−3 | <7.26 × 10−3 | <3.63 × 10−2 |
| B22 | 3.63 × 100 | 2.79 × 101 | <5.01 × 10−3 | <5.01 × 10−3 | <5.01 × 10−3 | <2.51 × 10−2 |
| Sample | Gram-Negative Bacteria | Gram-Positive Bacteria | ||
|---|---|---|---|---|
| Enterobacteriaceae | E. coli | Enterococcus spp. | Listeria spp. | |
| A1 | VRBGA/VRBA/SCA | KAE/SBV | ||
| A2 | VRBGA/VRBA/SCA | KAE | ||
| A3 | KAE | |||
| A4 | VRBGA | |||
| A6 | VRBGA/VRBA | KAE | ||
| A7 | VRBGA/VRBA | KAE/SBV | ||
| A10 | VRBGA | |||
| A11 | SBV | |||
| A12 | VRBGA/VRBA | KAE | ||
| A13 | VRBGA | OCLA | ||
| A14 | VRBGA | |||
| A17 | VRBGA | |||
| A20 | VRBGA/VRBA | |||
| A21 | VRBGA | KAE | ||
| A22 | VRBGA/SCA | KAE | OCLA | |
| B2 | VRBGA/VRBA | KAE | ||
| B3 | KAE | |||
| B4 | SCA | KAE | ||
| B5 | VRBGA | |||
| B6 | VRBGA/VRBA | KAE | ||
| B10 | VRBGA | VRBA | ||
| B11 | VRBGA/VRBA | SBV | ||
| B14 | VRBGA/VRBA | |||
| B15 | VRBGA | VRBA/SCA | ||
| B18 | VRBGA/VRBA | |||
| B20 | VRBA | SBV | ||
| Slaughterhouse A | Slaughterhouse B | ||||
|---|---|---|---|---|---|
| Identification | Surface | Area (cm2) | Identification | Surface | Area (cm2) |
| A1 | Stunning tank | 3544.5 | B1 | Stunning tank | 976.0 |
| A2 | Slaughter area | 960.8 | B2 | Slaughter area | 945.0 |
| A3 | Bleeding tank | 2550.0 | B3 | Bleeding tank | 1480.0 |
| A4 | Sink (outer surface) | 800.0 | B4 | Sink (external surface) | 775.0 |
| A5 | Scalding tank door | 6842.5 | B5 | Scalding tank door | 6870.0 |
| A6 | Plucker (rubber fingers) | 175.3 | B6 | Plucker (rubber fingers) | 113.1 |
| A7 | Cloacal sphincter splitter | 5351.5 | B7 | Cloacal sphincter splitter | 2190.0 |
| A8 | Abdominal opening equipment | 4515.8 | B8 | Abdominal opening equipment | 2190.0 |
| A9 | Eviscerator | 1750.8 | B9 | Eviscerator | 1752.0 |
| A10 | Abdominal cavity aspirator equipment | 6360.3 | B10 | Leg cutter | 3294.0 |
| A11 | Trachea removal equipment | 1295.8 | B11 | Head cutter | 1050.0 |
| A12 | Neck cutter | 1500.0 | B12 | Gizzard conveyor belt | 308.2 |
| A13 | Spray washing equipment | 2475.0 | B13 | Crop extractor | 836.0 |
| A14 | Leg and neck carrying belt | 2760.0 | B14 | Bone neck cutter | 1100.0 |
| A15 | Sorter | 2915.0 | B15 | Abdominal aspiration equipment | 3290.0 |
| A16 | Box conveyor belt | 3876.7 | B16 | Leg conveyor belt | 1449.0 |
| A17 | Cutting board | 2054.4 | B17 | Liver conveyor belt | 1944.0 |
| A18 | Chopper | 2707.3 | B18 | Pre-cooling rollers | 1764.0 |
| A19 | Kneader | 326.7 | B19 | Container for collecting chicken wings | 3760.0 |
| A20 | Stuffer | 2040.0 | B20 | Breast conveyor belt | 1443.0 |
| A21 | Meat conveyor belt | 3550.0 | B21 | Thigh conveyor belt | 1378.0 |
| A22 | Packaging area cutting board | 1935.0 | B22 | Packaging area cutting board | 1995.0 |
| Microorganism | Culture Media | Incubation (T/t) | Reference |
|---|---|---|---|
| TAC 1 | PCA | 30 °C/72 h | [73] |
| Psychrotrophs 1 | PCA | 7 °C/10 days | [74] |
| Enterobacteria 2,3 | VRBGA | 37 °C/24 h | [75] |
| Coliforms 2,3 | VRBA | 42 °C/24 h | [75] |
| Enterococci 2 | KAE | 42 °C/24 h | [75] |
| Vancomycin-resistant enterococci 1 | SBV | 42 °C/48 h | [76] |
| Listeria spp. | Semi-Fraser broth | 30 °C/24 h | |
| Fraser broth | 37 °C/24 h | UNE-EN ISO 11290-1:2018 [71] | |
| OCLA | 37 °C/48 h | ||
| Salmonella spp. | Buffered peptone water | 37 °C/24 h | |
| Rappaport-Vassiliadis broth | 42 °C/24 h | UNE-EN ISO 6579-1:2017 [72] | |
| SCA | 42 °C/24 h |
| Microorganism | Primer | Sequence (5′ to 3′) | Size | Program |
|---|---|---|---|---|
| Enterobacteriaceae | rpoB-F | CAGGTCGTCACGGTAACAAG | 512 pb | (94 °C—30 s/60 °C—30 s/72 °C—60 s) × 35 |
| rpoB-R | GTGGTTCAGTTTCAGCATGTAC | |||
| Escherichia coli | uidA-F | GTCACGCCGTATGTTATTG | 530 pb | (94 °C—30 s/58 °C—60 s/72 °C—90 s) × 35 |
| uidA-R | CCAAAGCCAGTAAAGTAGAAC | |||
| Enterococcus spp. | 16S-F | TCAACCGGGGAGGGT | 738 pb | (94 °C—30 s/60 °C—60 s/72 °C—45 s) × 30 |
| 16S-R | ATTACTAGCGATTCCGG | |||
| Listeria spp. | prs-F | GCTGAAGAGATTGCGAAAGAAG | 370 pb | (94 °C—30 s/58 °C—30 s/72 °C—45 s) × 35 |
| prs-R | CAAAGAAACCTTGGATTTGCGG |
| Gene | Primer | Sequence(5′ to 3′) | Size | Reference |
|---|---|---|---|---|
| blaTEM | F | GATAAATCTGGAGCCGGTGA | 78 pb | [16] |
| R | GATACGGGAGGGCTTACCAT | |||
| blaCTX-M | F | CACCAATGATATTGCGGTGA | 77 pb | [16] |
| R | GTTGCGGCTGGGTAAAATAG | |||
| blaKPC | F | CTGTATCGCCGTCTAGTTCTG | 101 pb | [83] |
| R | AGTTTAGCGAATGGTTCCG | |||
| blaCMY-2 | F | CCAGAACTGACAGGCAAACA | 65 pb | [16] |
| R | CCTGCCGTATAGGTGGCTAA | |||
| blaNDM | F | ATGGAGACTGGCGACCAAC | 87 pb | [16] |
| R | GGCATGTCGAGATAGGAAGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panera-Martínez, S.; Rodríguez-Melcón, C.; Rodríguez-Campos, D.; Pérez-Estébanez, N.; Capita, R.; Alonso-Calleja, C. Levels of Different Microbial Groups on Inert Surfaces of Poultry Slaughterhouses: Identification Using Matrix-Assisted Laser Desorption Ionization Time-of-Flight and Detection of Extended-Spectrum Beta-Lactamase- and Carbapenemase-Producing Enterobacteria. Antibiotics 2024, 13, 587. https://doi.org/10.3390/antibiotics13070587
Panera-Martínez S, Rodríguez-Melcón C, Rodríguez-Campos D, Pérez-Estébanez N, Capita R, Alonso-Calleja C. Levels of Different Microbial Groups on Inert Surfaces of Poultry Slaughterhouses: Identification Using Matrix-Assisted Laser Desorption Ionization Time-of-Flight and Detection of Extended-Spectrum Beta-Lactamase- and Carbapenemase-Producing Enterobacteria. Antibiotics. 2024; 13(7):587. https://doi.org/10.3390/antibiotics13070587
Chicago/Turabian StylePanera-Martínez, Sarah, Cristina Rodríguez-Melcón, Daniel Rodríguez-Campos, Nuria Pérez-Estébanez, Rosa Capita, and Carlos Alonso-Calleja. 2024. "Levels of Different Microbial Groups on Inert Surfaces of Poultry Slaughterhouses: Identification Using Matrix-Assisted Laser Desorption Ionization Time-of-Flight and Detection of Extended-Spectrum Beta-Lactamase- and Carbapenemase-Producing Enterobacteria" Antibiotics 13, no. 7: 587. https://doi.org/10.3390/antibiotics13070587
APA StylePanera-Martínez, S., Rodríguez-Melcón, C., Rodríguez-Campos, D., Pérez-Estébanez, N., Capita, R., & Alonso-Calleja, C. (2024). Levels of Different Microbial Groups on Inert Surfaces of Poultry Slaughterhouses: Identification Using Matrix-Assisted Laser Desorption Ionization Time-of-Flight and Detection of Extended-Spectrum Beta-Lactamase- and Carbapenemase-Producing Enterobacteria. Antibiotics, 13(7), 587. https://doi.org/10.3390/antibiotics13070587