
Relevance of the Adjuvant Effect between Cellular Homeostasis and Resistance to Antibiotics in Gram-Negative Bacteria with Pathogenic Capacity: A Study of Klebsiella pneumoniae

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Bacterial Homeostasis Mechanisms
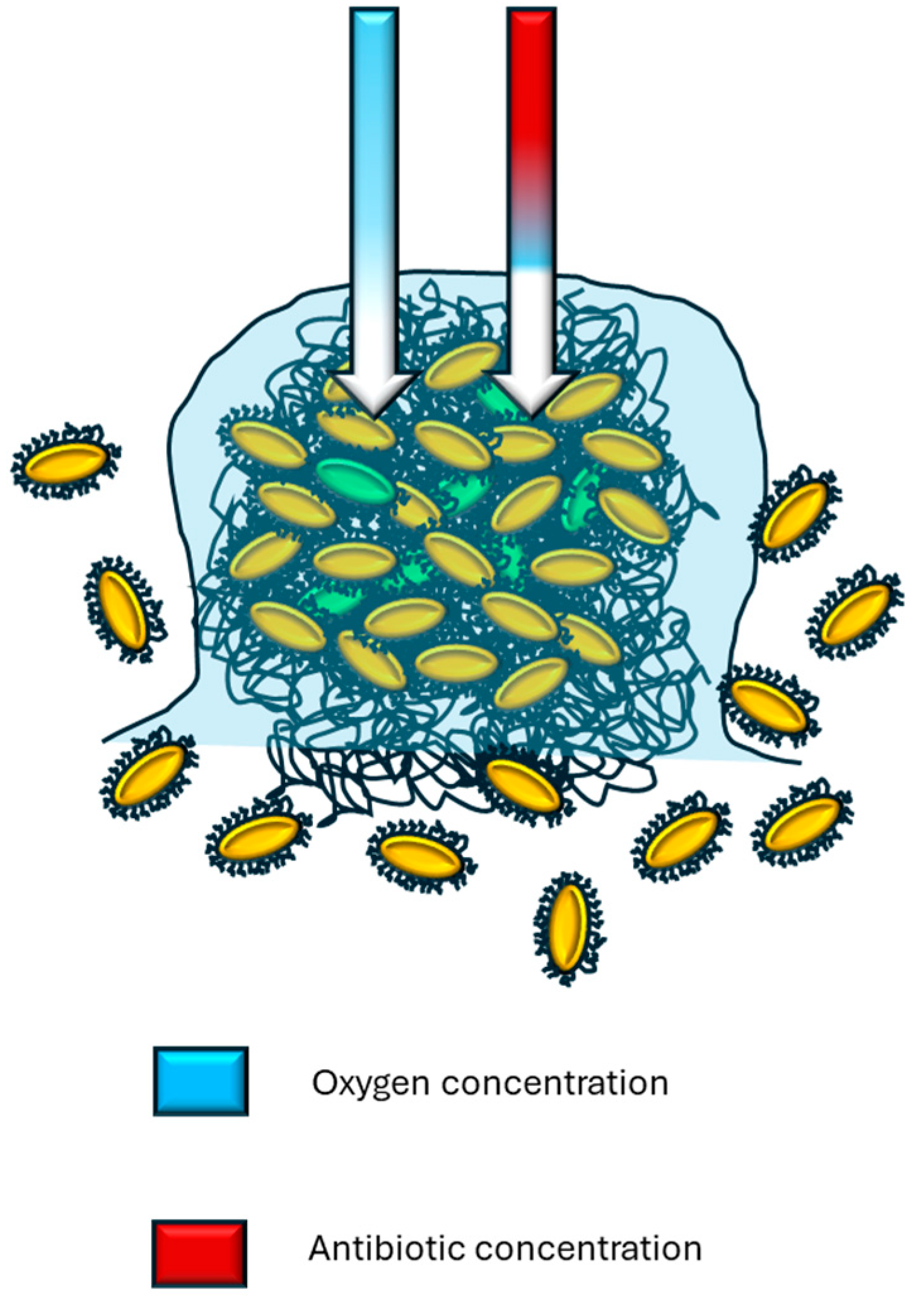
2.1. Nonspecific Mechanism and Biofilm Formation
2.2. Stressful Ecosystem Conditions and Adaptive Response
3. Klebsiella pneumoniae: Adequacy and Resistance Phenotype
4. Molecular Basis and Physiological Principles of Antimicrobial Resistance Mechanisms in K. pneumoniae
5. The Impact of Cellular Homeostasis on K. pneumoniae Bacterial Resistance
5.1. Cellular Efflux
5.2. Heat Shock Proteins
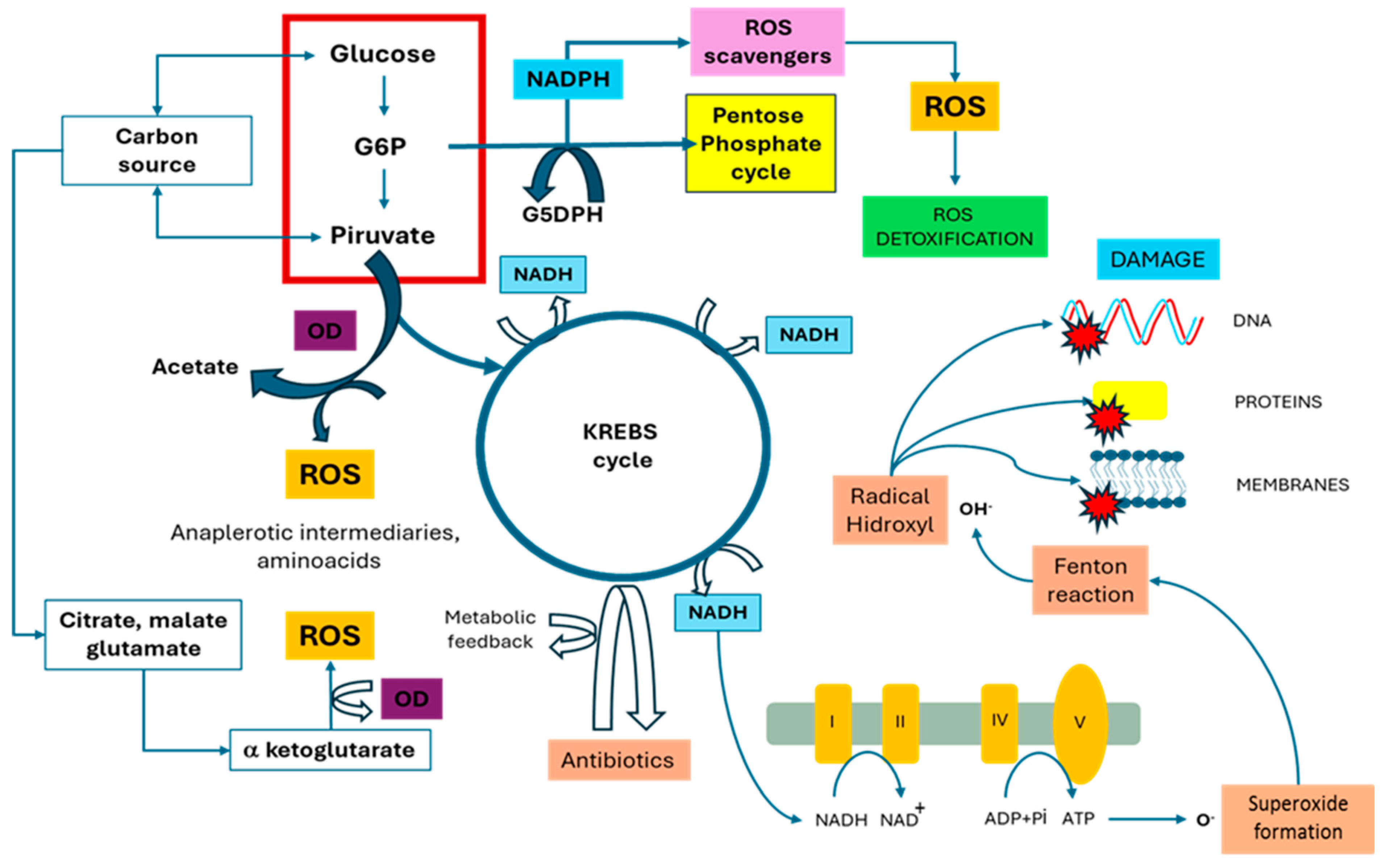
5.3. Response to O2 Availability and Reactive Oxygen Species
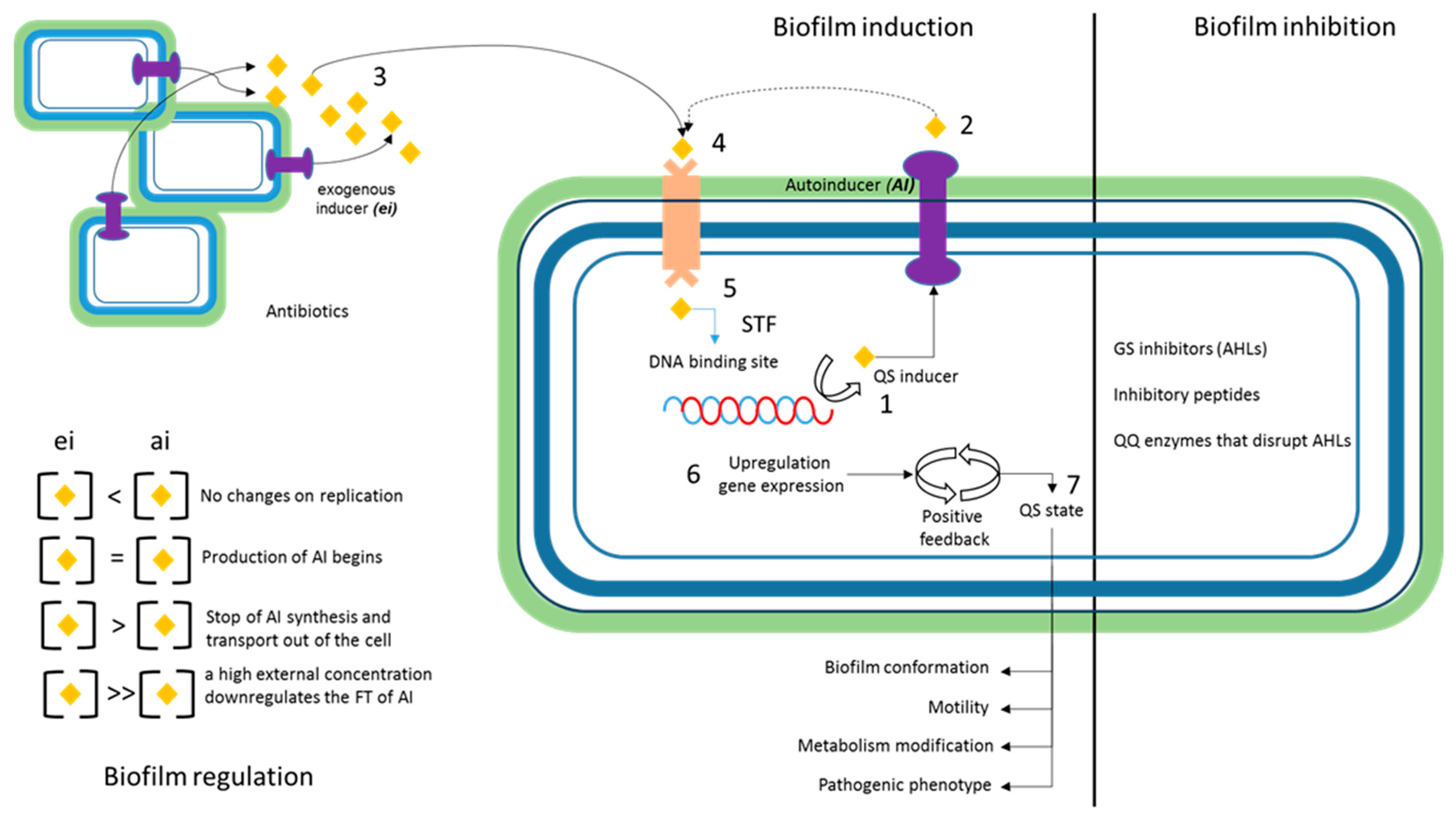
5.4. Quorum Sensing
5.5. pH Conditions Regulate Antibiotic Resistance
6. Transcriptional Regulation and Cryptic Resistance Mechanisms
7. Moonlighting Proteins Associated with Resistance
8. Conclusions
9. Recommendations for Future Research
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davies, J.; Davies, D. Origins and Evolution of Antibiotic Resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [PubMed]
- Perry, J.; Waglechner, N.; Wright, G. The Prehistory of Antibiotic Resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025197. [Google Scholar] [CrossRef] [PubMed]
- D’Costa, V.M.; King, C.E.; Kalan, L.; Morar, M.; Sung, W.W.L.; Schwarz, C.; Froese, D.; Zazula, G.; Calmels, F.; Debruyne, R.; et al. Antibiotic resistance is ancient. Nature 2011, 477, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Panta, P.R.; Doerrler, W.T. A link between pH homeostasis and colistin resistance in bacteria. Sci. Rep. 2021, 11, 13230. [Google Scholar] [CrossRef] [PubMed]
- Sułek, A.; Pucelik, B.; Kobielusz, M.; Barzowska, A.; Dąbrowski, J.M. Photodynamic Inactivation of Bacteria with Porphyrin Derivatives: Effect of Charge, Lipophilicity, ROS Generation, and Cellular Uptake on Their Biological Activity In Vitro. Int. J. Mol. Sci. 2020, 21, 8716. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Li, P.; Liu, H.; Lü, D.; Liang, H.; Dou, Y. Phenotypic and Molecular Characterization of Multidrug-Resistant Klebsiella pneumoniae Isolated from a University Teaching Hospital, China. PLoS ONE 2014, 9, e95181. [Google Scholar] [CrossRef]
- Kalpana, S.; Lin, W.Y.; Wang, Y.C.; Fu, Y.; Lakshmi, A.; Wang, H.Y. Antibiotic Resistance Diagnosis in ESKAPE Pathogens—A Review on Proteomic Perspective. Diagnostics 2023, 13, 1014. [Google Scholar] [CrossRef] [PubMed]
- Darby, E.M.; Trampari, E.; Siasat, P.; Gaya, M.S.; Alav, I.; Webber, M.A.; Blair, J.M. Molecular mechanisms of antibiotic resistance revisited. Nat. Rev. Microbiol. 2023, 21, 280–295. [Google Scholar] [CrossRef]
- Van Hoek, A.H.A.M.; Mevius, D.; Guerra, B.; Mullany, P.; Roberts, A.P.; Aarts, H.J.M. Acquired Antibiotic Resistance Genes: An Overview. Front. Microbiol. 2011, 2, 203. Available online: http://journal.frontiersin.org/article/10.3389/fmicb.2011.00203/abstract (accessed on 18 March 2024). [CrossRef]
- Olivares, J.; Bernardini, A.; Garcia-Leon, G.; Corona, F.; Sanchez, M.B.; Martinez, J.L. The intrinsic resistome of bacterial pathogens. Front. Microbiol. 2013, 4, 103. Available online: http://journal.frontiersin.org/article/10.3389/fmicb.2013.00103/abstract (accessed on 18 March 2024). [CrossRef]
- Huang, L.; Ahmed, S.; Gu, Y.; Huang, J.; An, B.; Wu, C.; Zhou, Y.; Cheng, G. The Effects of Natural Products and Environmental Conditions on Antimicrobial Resistance. Molecules 2021, 26, 4277. [Google Scholar] [CrossRef] [PubMed]
- Cruaud, P.; Vigneron, A.; Dorea, C.C.; Rodriguez, M.J.; Charette, S.J. Rapid Changes in Microbial Community Structures along a Meandering River. Microorganisms 2020, 8, 1631. [Google Scholar] [CrossRef] [PubMed]
- Lau, J.A.; Lennon, J.T. Rapid responses of soil microorganisms improve plant fitness in novel environments. Proc. Natl. Acad. Sci. USA 2012, 109, 14058–14062. [Google Scholar] [CrossRef]
- Rosenberg, E. Rapid acquisition of microorganisms and microbial genes can help explain punctuated evolution. Front. Ecol. Evol. 2022, 10, 957708. [Google Scholar] [CrossRef]
- Kracke, F.; Vassilev, I.; Krömer, J.O. Microbial electron transport and energy conservation—The foundation for optimizing bioelectrochemical systems. Front. Microbiol. 2015, 6, 146906. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A. Electron Transport Chain. In Sweet Biochemistry; Elsevier: Amsterdam, The Netherlands, 2018; pp. 13–16. Available online: https://linkinghub.elsevier.com/retrieve/pii/B9780128144534000030 (accessed on 18 March 2024).
- Booth, J.A.; Špírek, M.; Lobie, T.A.; Skarstad, K.; Krejci, L.; Bjørås, M. Antibiotic-induced DNA damage results in a controlled loss of pH homeostasis and genome instability. Sci. Rep. 2020, 10, 19422. [Google Scholar] [CrossRef] [PubMed]
- Choby, J.E.; Howard-Anderson, J.; Weiss, D.S. Hypervirulent Klebsiella pneumoniae—Clinical and molecular perspectives. J. Intern. Med. 2020, 287, 283–300. [Google Scholar] [CrossRef] [PubMed]
- Germain, E.; Castro-Roa, D.; Zenkin, N.; Gerdes, K. Molecular Mechanism of Bacterial Persistence by HipA. Mol. Cell 2013, 52, 248–254. [Google Scholar] [CrossRef]
- Martínez, J.L.; Rojo, F. Metabolic regulation of antibiotic resistance. FEMS Microbiol. Rev. 2011, 35, 768–789. [Google Scholar] [CrossRef]
- O’Toole, G.A.; Gibbs, K.A.; Hager, P.W.; Phibbs, P.V.; Kolter, R. The Global Carbon Metabolism Regulator Crc Is a Component of a Signal Transduction Pathway Required for Biofilm Development by Pseudomonas aeruginosa. J. Bacteriol. 2000, 182, 425–431. [Google Scholar] [CrossRef]
- Rittershaus, E.S.C.; Baek, S.H.; Sassetti, C.M. The Normalcy of Dormancy: Common Themes in Microbial Quiescence. Cell Host Microbe 2013, 13, 643–651. [Google Scholar] [CrossRef]
- Sachidanandham, R.; Yew-Hoong Gin, K. A dormancy state in nonspore-forming bacteria. Appl. Microbiol. Biotechnol. 2009, 81, 927–941. [Google Scholar] [CrossRef] [PubMed]
- Vuotto, C.; Longo, F.; Pascolini, C.; Donelli, G.; Balice, M.P.; Libori, M.F.; Tiracchia, V.; Salvia, A.; Varaldo, P.E. Biofilm formation and antibiotic resistance in Klebsiella pneumoniae urinary strains. J. Appl. Microbiol. 2017, 123, 1003–1018. [Google Scholar] [CrossRef]
- Fazeli-Nasab, B.; Sayyed, R.Z.; Mojahed, L.S.; Rahmani, A.F.; Ghafari, M.; Antonius, S. Biofilm production: A strategic mechanism for survival of microbes under stress conditions. Biocatal. Agric. Biotechnol. 2022, 42, 102337. [Google Scholar] [CrossRef]
- Moya, C.; Maicas, S. Antimicrobial Resistance in Klebsiella pneumoniae Strains: Mechanisms and Outbreaks. In The 1st International Electronic Conference on Microbiology; MDPI: Basel, Switzerland, 2020; p. 11. Available online: https://www.mdpi.com/2504-3900/66/1/11 (accessed on 18 March 2024).
- Mirghani, R.; Saba, T.; Khaliq, H.; Mitchell, J.; Do, L.; Chambi, L.; Diazm, K.; Kenedy, T.; Alkassab, K.; Juynh, T.; et al. Biofilms: Formation, drug resistance and alternatives to conventional approaches. AIMS Microbiol. 2022, 8, 239–277. [Google Scholar] [CrossRef] [PubMed]
- López-Pérez, M.; Viniegra-González, G. Production of protein and metabolites by yeast grown in solid state fermentation: Present status and perspectives. J. Chem. Technol. Biotechnol. 2016, 91, 1224–1231. [Google Scholar] [CrossRef]
- Garcia Mendez, D.F.; Rengifo Herrera, J.A.; Sanabria, J.; Wist, J. Analysis of the Metabolic Response of Planktonic Cells and Biofilms of Klebsiella pneumoniae to Sublethal Disinfection with Sodium Hypochlorite Measured by NMR. Microorganisms 2022, 10, 1323. [Google Scholar] [CrossRef]
- Van Laar, T.A.; Chen, T.; You, T.; Leung, K.P. Sublethal Concentrations of Carbapenems Alter Cell Morphology and Genomic Expression of Klebsiella pneumoniae Biofilms. Antimicrob. Agents Chemother. 2015, 59, 1707–1717. [Google Scholar] [CrossRef]
- Li, Y.; Xia, L.; Chen, J.; Lian, Y.; Dandekar, A.A.; Xu, F.; Wang, M. Resistance elicited by sub-lethal concentrations of ampicillin is partially mediated by quorum sensing in Pseudomonas aeruginosa. Environ. Int. 2021, 156, 106619. [Google Scholar] [CrossRef]
- Wesche, A.M.; Gurtler, J.B.; Marks, B.P.; Ryser, E.T. Stress, Sublethal Injury, Resuscitation, and Virulence of Bacterial Foodborne Pathogens. J. Food Prot. 2009, 72, 1121–1138. [Google Scholar] [CrossRef]
- Yin, W.; Wang, Y.; Liu, L.; He, J. Biofilms: The Microbial “Protective Clothing” in Extreme Environments. Int. J. Mol. Sci. 2019, 20, 3423. [Google Scholar] [CrossRef] [PubMed]
- Nirwati, H.; Sinanjung, K.; Fahrunissa, F.; Wijaya, F.; Napitupulu, S.; Hati, V.P.; Hakim, M.S.; Meliala, A.; Aman, A.T.; Nuryastuti, T. Biofilm formation and antibiotic resistance of Klebsiella pneumoniae isolated from clinical samples in a tertiary care hospital, Klaten, Indonesia. BMC Proc. 2019, 13, 20. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Rose, J.; Huang, S.; Hu, Y.; Wu, Q.; Wang, D.; Li, C.; Liu, M.; Zhou, P.; Jiang, L. A pH-gated conformational switch regulates the phosphatase activity of bifunctional HisKA-family histidine kinases. Nat. Commun. 2017, 8, 2104. [Google Scholar] [CrossRef] [PubMed]
- Purvis, J.E.; Yomano, L.P.; Ingram, L.O. Enhanced Trehalose Production Improves Growth of Escherichia coli under Osmotic Stress. Appl. Environ. Microbiol. 2005, 71, 3761–3769. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Zhang, K.; Fan, Y.; Fang, H.; Nie, Y.; Wu, X.L. The Bacterial MtrAB Two-Component System Regulates the Cell Wall Homeostasis Responding to Environmental Alkaline Stress. Microbiol. Spectr. 2022, 10, e02311-22. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.T.; Peng, H.L. Regulation of the Homologous Two-Component Systems KvgAS and KvhAS in Klebsiella pneumoniae CG43. J. Biochem. 2006, 140, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Pinto, N.A.; Kim, C.Y.; Yang, S.; D’Souza, R.; Yong, D.; Lee, I. Network Integrative Genomic and Transcriptomic Analysis of Carbapenem-Resistant Klebsiella pneumoniae Strains Identifies Genes for Antibiotic Resistance and Virulence. mSystems 2019, 4, e00202-19. [Google Scholar] [CrossRef] [PubMed]
- Krulwich, T.A.; Sachs, G.; Padan, E. Molecular aspects of bacterial pH sensing and homeostasis. Nat. Rev. Microbiol. 2011, 9, 330–343. [Google Scholar] [CrossRef]
- Poolman, B. Physicochemical homeostasis in bacteria. FEMS Microbiol. Rev. 2023, 47, fuad033. [Google Scholar] [CrossRef]
- Herrmann, H.A.; Dyson, B.C.; Vass, L.; Johnson, G.N.; Schwartz, J.M. Flux sampling is a powerful tool to study metabolism under changing environmental conditions. Npj Syst. Biol. Appl. 2019, 5, 32. [Google Scholar] [CrossRef]
- Galperin, M.Y. Bacterial signal transduction network in a genomic perspective. Environ. Microbiol. 2004, 6, 552–567. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- Harada, K.; Asai, T. Role of Antimicrobial Selective Pressure and Secondary Factors on Antimicrobial Resistance Prevalence in Escherichia coli from Food-Producing Animals in Japan. J. Biomed. Biotechnol. 2010, 2010, 180682. [Google Scholar] [CrossRef] [PubMed]
- Tello, A.; Austin, B.; Telfer, T.C. Selective Pressure of Antibiotic Pollution on Bacteria of Importance to Public Health. Environ. Health Perspect. 2012, 120, 1100–1106. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yu, Z.; Ding, T. Quorum-Sensing Regulation of Antimicrobial Resistance in Bacteria. Microorganisms 2020, 8, 425. [Google Scholar] [CrossRef] [PubMed]
- Effah, C.Y.; Sun, T.; Liu, S.; Wu, Y. Klebsiella pneumoniae: An increasing threat to public health. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 1. [Google Scholar] [CrossRef]
- Lery, L.M.; Frangeul, L.; Tomas, A.; Passet, V.; Almeida, A.S.; Bialek-Davenet, S.; Barbe, V.; Bengoechea, J.A.; Sansonetti, P.; Brisse, S.; et al. Comparative analysis of Klebsiella pneumoniae genomes identifies a phospholipase D family protein as a novel virulence factor. BMC Biol. 2014, 12, 41. [Google Scholar] [CrossRef]
- Bengoechea, J.A.; Sa Pessoa, J. Klebsiella pneumoniae infection biology: Living to counteract host defences. FEMS Microbiol. Rev. 2019, 42, 123–144. [Google Scholar] [CrossRef]
- Brisse, S.; Fevre, C.; Passet, V.; Issenhuth-Jeanjean, S.; Tournebize, R.; Diancourt, L.; Grimont, P. Virulent Clones of Klebsiella pneumoniae: Identification and Evolutionary Scenario Based on Genomic and Phenotypic Characterization. PLoS ONE 2009, 4, e4982. [Google Scholar] [CrossRef]
- Gorrie, C.L.; Mirčeta, M.; Wick, R.R.; Judd, L.M.; Lam, M.M.C.; Gomi, R.; Abbott, I.J.; Thomson, N.R.; Strugnell, R.A.; Pratt, N.F.; et al. Genomic dissection of Klebsiella pneumoniae infections in hospital patients reveals insights into an opportunistic pathogen. Nat. Commun. 2022, 13, 3017. [Google Scholar] [CrossRef]
- Holt, K.E.; Wertheim, H.; Zadoks, R.N.; Baker, S.; Whitehouse, C.A.; Dance, D.; Jenney, A.; Connor, T.R.; Hsu, L.Y.; Severin, J.; et al. Genomic analysis of diversity, population structure, virulence, and antimicrobial resistance in Klebsiella pneumoniae, an urgent threat to public health. Proc. Natl. Acad. Sci. USA 2015, 112, E3574–E3581. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, R.A.; Klopprogge, K.; Grabbe, R. Regulation of nitrogen fixation in Klebsiella pneumoniae and Azotobacter vinelandii: NifL, transducing two environmental signals to the nif transcriptional activator NifA. J. Mol. Microbiol. Biotechnol. 2002, 4, 235–242. [Google Scholar]
- Xiao, Y.; Zhang, X.; Zhu, M.; Tan, W. Effect of the culture media optimization, pH and temperature on the biohydrogen production and the hydrogenase activities by Klebsiella pneumoniae ECU-15. Bioresour. Technol. 2013, 137, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Torres, A.J.; Prieto, L.; Hazen, T.C. Comparison of the in situ survival and activity of Klebsiella pneumoniae and Escherichia coli in tropical marine environments. Microb. Ecol. 1988, 15, 41–57. [Google Scholar] [CrossRef]
- Raffelsberger, N.; Hetland, M.A.K.; Svendsen, K.; Småbrekke, L.; Löhr, I.H.; Andreassen, L.L.E.; Brisse, S.; Holt, K.E.; Sundsfjord, A.; Samuelsen, Ø.; et al. Gastrointestinal carriage of Klebsiella pneumoniae in a general adult population: A cross-sectional study of risk factors and bacterial genomic diversity. Gut Microbes 2021, 13, 1939599. [Google Scholar] [CrossRef] [PubMed]
- Ochońska, D.; Ścibik, Ł.; Brzychczy-Włoch, M. Biofilm Formation of Clinical Klebsiella pneumoniae Strains Isolated from Tracheostomy Tubes and Their Association with Antimicrobial Resistance, Virulence and Genetic Diversity. Pathogens 2021, 10, 1345. [Google Scholar] [CrossRef] [PubMed]
- Walker, K.A.; Miller, V.L. The intersection of capsule gene expression, hypermucoviscosity and hypervirulence in Klebsiella pneumoniae. Curr. Opin. Microbiol. 2020, 54, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kumar, S.; Zhang, L.; Wu, H.; Wu, H. Characteristics of antibiotic resistance mechanisms and genes of Klebsiella pneumoniae. Open Med. 2023, 18, 20230707. [Google Scholar] [CrossRef] [PubMed]
- Padilla, E.; Llobet, E.; Doménech-Sánchez, A.; Martínez-Martínez, L.; Bengoechea, J.A.; Albertí, S. Klebsiella pneumoniae AcrAB Efflux Pump Contributes to Antimicrobial Resistance and Virulence. Antimicrob. Agents Chemother. 2010, 54, 177–183. [Google Scholar] [CrossRef]
- Xu, Q.; Sheng, Z.; Hao, M.; Jiang, J.; Ye, M.; Chen, Y.; Xu, X.; Guo, Q.; Wang, M. RamA upregulates multidrug resistance efflux pumps AcrAB and OqxAB in Klebsiella pneumoniae. Int. J. Antimicrob. Agents 2021, 57, 106251. [Google Scholar] [CrossRef]
- Badger-Emeka, L.I.; Emeka, P.M. Genetic fingerprinting and profile analysis of virulence genes in XDR clinical isolates of Klebsiella Pneumoniae. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 5233–5244. [Google Scholar] [CrossRef] [PubMed]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Nisar, A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; et al. Antibiotic resistance: A rundown of a global crisis. Infect. Drug Resist. 2018, 11, 1645–1658. [Google Scholar] [CrossRef]
- Horng, Y.T.; Panjaitan, N.S.D.; Tsai, Y.J.; Su, P.W.; Yang, H.C.; Soo, P.C. The role of EII complex in the bacterial responses to the glucose-survey in clinical Klebsiella pneumoniae isolates. Afzal M, editor. PLoS ONE 2023, 18, e0289759. [Google Scholar] [CrossRef]
- Zhao, Q.; Su, X.; Wang, Y.; Liu, R.; Bartlam, M. Structural analysis of molybdate binding protein ModA from Klebsiella pneumoniae. Biochem. Biophys. Res. Commun. 2023, 681, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Cesur, M.F.; Siraj, B.; Uddin, R.; Durmuş, S.; Çakır, T. Network-Based Metabolism-Centered Screening of Potential Drug Targets in Klebsiella pneumoniae at Genome Scale. Front. Cell. Infect. Microbiol. 2020, 9, 447. [Google Scholar] [CrossRef] [PubMed]
- Shebl, R.I.; Elkhatib, W.F.; Badawy, M.S.E.M. Modulating the transcriptomic profile of multidrug-resistant Klebsiella pneumoniae biofilm formation by antibiotics in combination with zinc sulfate. Ann. Clin. Microbiol. Antimicrob. 2023, 22, 84. [Google Scholar] [CrossRef]
- Li, Y.; Kumar, S.; Zhang, L.; Wu, H. Klebsiella pneumonia and Its Antibiotic Resistance: A Bibliometric Analysis. BioMed Res. Int. 2022, 2022, 1668789. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, K.; Ding, C.; Wang, S.; Wu, W.; Liu, X. Exploring multidrug-resistant Klebsiella pneumoniae antimicrobial resistance mechanisms through whole genome sequencing analysis. BMC Microbiol. 2023, 23, 245. [Google Scholar] [CrossRef]
- Rice, L.B.; Carias, L.L.; Hujer, A.M.; Bonafede, M.; Hutton, R.; Hoyen, C.; Bonomo, R.A. High-Level Expression of Chromosomally Encoded SHV-1 β-Lactamase and an Outer Membrane Protein Change Confer Resistance to Ceftazidime and Piperacillin-Tazobactam in a Clinical Isolate of Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2000, 44, 362–367. [Google Scholar] [CrossRef]
- Singh, P. The incidence of AmpC β-lactamases producing Klebsiella pneumoniae subspecies pneumoniae. Int. J. Res. Med. Sci. 2018, 6, 1169. [Google Scholar] [CrossRef]
- Li, J.; Liu, F.; Wang, Q.; Ge, P.; Woo, P.C.Y.; Yan, J.; Zhao, Y.; Gao, G.F.; Liu, C.H.; Liu, C. Genomic and transcriptomic analysis of NDM-1 Klebsiella pneumoniae in spaceflight reveal mechanisms underlying environmental adaptability. Sci. Rep. 2014, 4, 6216. [Google Scholar] [CrossRef] [PubMed]
- De Majumdar, S.; Yu, J.; Fookes, M.; McAteer, S.P.; Llobet, E.; Finn, S.; Spence, S.; Monaghan, A.; Kissenpfennig, A.; Ingram, R.J.; et al. Elucidation of the RamA Regulon in Klebsiella pneumoniae Reveals a Role in LPS Regulation. PLoS Pathog. 2015, 11, e1004627. [Google Scholar] [CrossRef] [PubMed]
- Anes, J.; Dever, K.; Eshwar, A.; Nguyen, S.; Cao, Y.; Sivasankaran, S.K.; Sakalauskaitė, S.; Lehner, A.; Devineau, S.; Daugelavičius, R.; et al. Analysis of the Oxidative Stress Regulon Identifies soxS as a Genetic Target for Resistance Reversal in Multidrug-Resistant Klebsiella pneumoniae. mBio 2021, 12, e00867-21. [Google Scholar] [CrossRef] [PubMed]
- Chubiz, L.M.; Rao, C.V. Role of the mar-sox-rob Regulon in Regulating Outer Membrane Porin Expression. J. Bacteriol. 2011, 193, 2252–2260. [Google Scholar] [CrossRef] [PubMed]
- Albarri, O.; AlMatar, M.; Var, I.; Köksal, F. Antimicrobial Resistance of Clinical Klebsiella pneumoniae Isolates: Involvement of AcrAB and OqxAB Efflux Pumps. Curr. Mol. Pharmacol. 2023, 17, e310323215266. [Google Scholar] [CrossRef] [PubMed]
- Dersch, P.; Khan, M.A.; Mühlen, S.; Görke, B. Roles of Regulatory RNAs for Antibiotic Resistance in Bacteria and Their Potential Value as Novel Drug Targets. Front. Microbiol. 2017, 8, 803. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Sun, T.; Jia, P.; Li, S.; Li, X.; Shi, Y.; Li, X.; Gao, H.; Yin, H.; Jia, X.-M.; et al. A-to-I RNA Editing in Klebsiella pneumoniae Regulates Quorum Sensing and Affects Cell Growth and Virulence. Adv. Sci. 2023, 10, 2206056. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Zhang, H.; Lu, S.; Lai, C.; Liu, H.; Zhu, H. The metabolic flux regulation of Klebsiella pneumoniae based on quorum sensing system. Sci. Rep. 2016, 6, 38725. [Google Scholar] [CrossRef]
- Sun, L.; Rasmussen, P.K.; Bai, Y.; Chen, X.; Cai, T.; Wang, J.; Guo, X.; Xie, Z.; Ding, X.; Niu, L.; et al. Proteomic Changes of Klebsiella pneumoniae in Response to Colistin Treatment and crrB Mutation-Mediated Colistin Resistance. Antimicrob. Agents Chemother. 2020, 64, e02200-19. [Google Scholar] [CrossRef]
- Rocha, J.; Henriques, I.; Gomila, M.; Manaia, C.M. Common and distinctive genomic features of Klebsiella pneumoniae thriving in the natural environment or in clinical settings. Sci. Rep. 2022, 12, 10441. [Google Scholar] [CrossRef]
- Sengupta, S.; Chattopadhyay, M.K.; Grossart, H.P. The multifaceted roles of antibiotics and antibiotic resistance in nature. Front. Microbiol. 2013, 4, 47. Available online: http://journal.frontiersin.org/article/10.3389/fmicb.2013.00047/abstract (accessed on 18 March 2024). [CrossRef] [PubMed]
- Nikaido, H.; Pagès, J.M. Broad-specificity efflux pumps and their role in multidrug resistance of Gram-negative bacteria. FEMS Microbiol. Rev. 2012, 36, 340–363. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.; Ma, J.; Rothnie, A.; Biggin, P.C.; Kerr, I.D. Towards understanding promiscuity in multidrug efflux pumps. Trends Biochem. Sci. 2014, 39, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Ozu, M.; Alvear-Arias, J.J.; Fernandez, M.; Caviglia, A.; Peña-Pichicoi, A.; Carrillo, C.; Carmona, E.; Otero-Gonzalez, A.; Garate, J.A.; Amodeo, G.; et al. Aquaporin Gating: A New Twist to Unravel Permeation through Water Channels. Int. J. Mol. Sci. 2022, 23, 12317. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, R.; Mori, M.; Segota, I.; Zhang, Z.; Aebersold, R.; Ludwig, C.; Hwa, T. Principles of gene regulation quantitatively connect DNA to RNA and proteins in bacteria. Science 2022, 378, eabk2066. [Google Scholar] [CrossRef] [PubMed]
- Koşar, Z.; Erbaş, A. Can the Concentration of a Transcription Factor Affect Gene Expression? Front. Soft Matter 2022, 2, 914494. [Google Scholar] [CrossRef]
- Li, Y.; Cross, T.S.; Dörr, T. Analysis of AcrB in Klebsiella pneumoniae reveals natural variants promoting enhanced multidrug resistance. Res. Microbiol. 2022, 173, 103901. [Google Scholar] [CrossRef]
- Al-Ruobayiee, M.R.; Ibrahim, A.H. The Relationship Between OqxAB Efflux Pump and Drug Resistance in Klebsiella pneumoniae Isolated from Clinical Sources. Al-Rafidain J. Med. Sci. 2023, 5, S106–S112. [Google Scholar] [CrossRef]
- Amereh, F.; Arabestani, M.R.; Shokoohizadeh, L. Relationship of OqxAB efflux pump to antibiotic resistance, mainly fluoroquinolones in Klebsiella pneumoniae, isolated from hospitalized patients. Iran J. Basic Med. Sci. 2022, 26, 93. [Google Scholar] [CrossRef]
- Ni, R.T.; Onishi, M.; Mizusawa, M.; Kitagawa, R.; Kishino, T.; Matsubara, F.; Tsuchiya, T.; Kuroda, T.; Ogawa, W. The role of RND-type efflux pumps in multidrug-resistant mutants of Klebsiella pneumoniae. Sci. Rep. 2020, 10, 10876. [Google Scholar] [CrossRef]
- Srinivasan, V.B.; Singh, B.B.; Priyadarshi, N.; Chauhan, N.K.; Rajamohan, G. Role of Novel Multidrug Efflux Pump Involved in Drug Resistance in Klebsiella pneumoniae. Forestier C, editor. PLoS ONE 2014, 9, e96288. [Google Scholar] [CrossRef] [PubMed]
- Akinpelu, S.; Ajayi, A.; Smith, S.I.; Adeleye, A.I. Efflux pump activity, biofilm formation and antibiotic resistance profile of Klebsiella spp. isolated from clinical samples at Lagos University Teaching Hospital. BMC Res. Notes 2020, 13, 258. [Google Scholar] [CrossRef] [PubMed]
- Ts, A.; Sg, B. Survival of Klebsiella pneumoniae strain borkar in pomegranate orchard soil and its tolerance to temperature and pH. J. Appl. Biotechnol. Bioeng. 2018, 5, 299–301. Available online: https://medcraveonline.com/JABB/survival-of-klebsiella-pneumoniae-strain-borkar-in-pomegranate-orchard-soil-and-its-tolerance-to-temperature-and-ph.html (accessed on 18 March 2024). [CrossRef]
- Tsuji, A.; Kaneko, Y.; Takahashi, K.; Ogawa, M.; Goto, S. The Effects of Temperature and pH on the Growth of Eight Enteric and Nine Glucose Non-Fermenting Species of Gram-Negative Rods. Microbiol. Immunol. 1982, 26, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Ortega, Y.; García-Latorre, E.A.; Lezama, R.A.; Luna-Herrera, J.; Vega-López, A.; Domínguez-López, M.L. Heat shock protein 60 from Klebsiella pneumoniae protects mononuclear cells from apoptotic cell death induced by dexamethasone. Microb. Pathog. 2011, 51, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Yang, J.; Qi, Z.; Wu, H.; Wang, B.; Zou, F.; Mei, H.; Liu, J.; Wang, W.; Liu, Q. Heat shock proteins: Biological functions, pathological roles, and therapeutic opportunities. MedComm 2022, 3, e161. [Google Scholar] [CrossRef]
- Cruz-Loya, M.; Kang, T.M.; Lozano, N.A.; Watanabe, R.; Tekin, E.; Damoiseaux, R.; Savage, V.M.; Yeh, P. Stressor interaction networks suggest antibiotic resistance co-opted from stress responses to temperature. ISME J. 2019, 13, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.D.H.; Kwon, H.Y.; Kim, E.H.; Kim, K.W.; Briles, D.E.; Pyo, S.; Rhee, D.K. Decrease in Penicillin Susceptibility Due to Heat Shock Protein ClpL in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2011, 55, 2714–2728. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Loya, M.; Tekin, E.; Kang, T.M.; Cardona, N.; Lozano-Huntelman, N.; Rodriguez-Verdugo, A.; Savage, V.M.; Yeh, P.J. Antibiotics Shift the Temperature Response Curve of Escherichia coli Growth. mSystems 2021, 6, e00228-21. [Google Scholar] [CrossRef]
- Fuqua, S.A.W.; Oesterreich, S.; Hilsenbeck, S.G.; Von Hoff, D.D.; Eckardt, J.; Osborne, C.K. Heat shock proteins and drug resistance. Breast Cancer Res. Treat. 1994, 32, 67–71. [Google Scholar] [CrossRef]
- Peinado-Ruiz, I.C.; Burgos-Molina, A.M.; Sendra-Portero, F.; Ruiz-Gómez, M.J. Relationship between heat shock proteins and cellular resistance to drugs and ageing. Exp. Gerontol. 2022, 167, 111896. [Google Scholar] [CrossRef]
- Gato, E.; Vázquez-Ucha, J.C.; Rumbo-Feal, S.; Álvarez-Fraga, L.; Vallejo, J.A.; Martínez-Guitián, M.; Beceiro, A.; Vivas, J.R.; Campoy, P.J.S.; Perez-Vazquez, M.; et al. Kpi, a chaperone-usher pili system associated with the worldwide-disseminated high-risk clone Klebsiella pneumoniae ST-15. Proc. Natl. Acad. Sci. USA 2020, 117, 17249–17259. [Google Scholar] [CrossRef] [PubMed]
- Lv, B.; Huang, X.; Lijia, C.; Ma, Y.; Bian, M.; Li, Z.; Duan, J.; Zhou, F.; Yang, B.; Qie, X.; et al. Heat shock potentiates aminoglycosides against gram-negative bacteria by enhancing antibiotic uptake, protein aggregation, and ROS. Proc. Natl. Acad. Sci. USA 2023, 120, e2217254120. [Google Scholar] [CrossRef]
- Ali, S.; Alam, M.; Hasan, G.M.; Hassan, M.I. Potential therapeutic targets of Klebsiella pneumoniae: A multi-omics review perspective. Brief. Funct. Genomics 2022, 21, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Grabbe, R.; Kuhn, A.; Schmitz, R.A. Cloning, sequencing and characterization of Fnr from Klebsiella pneumoniae. Antonie Van Leeuwenhoek 2001, 79, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, F.; Xu, L.; Wang, J.; Li, M.; Yuan, J.; Wang, H.; Yang, R.; Li, B. Cyclic AMP-CRP Modulates the Cell Morphology of Klebsiella pneumoniae in High-Glucose Environment. Front. Microbiol. 2020, 10, 2984. [Google Scholar] [CrossRef]
- Panjaitan, N.S.D.; Horng, Y.T.; Cheng, S.W.; Chung, W.T.; Soo, P.C. EtcABC, a Putative EII Complex, Regulates Type 3 Fimbriae via CRP-cAMP Signaling in Klebsiella pneumoniae. Front. Microbiol. 2019, 10, 1558. [Google Scholar] [CrossRef] [PubMed]
- Baez, A.; Shiloach, J. Effect of elevated oxygen concentration on bacteria, yeasts, and cells propagated for production of biological compounds. Microb. Cell Factories 2014, 13, 181. [Google Scholar] [CrossRef]
- Ray, P.D.; Huang, B.W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell. Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef]
- Addorisio, S.R.; Shteynberg, R.M.; Dasilva, M.S.; Mixon, J.M.; Mucciarone, K.; Vu, L.; Arsenault, K.L.; Briand, V.; Parker, S.; Smith, S.L.; et al. Oxidative Stress Response in Bacteria: A Review. Fine Focus 2022, 8, 36–46. [Google Scholar] [CrossRef]
- Carvajal-Garcia, J.; Samadpour, A.N.; Hernandez Viera, A.J.; Merrikh, H. Oxidative stress drives mutagenesis through transcription-coupled repair in bacteria. Proc. Natl. Acad. Sci. USA 2023, 120, e2300761120. [Google Scholar] [CrossRef]
- Liu, W.; Chen, G.; Dou, K.; Yi, B.; Wang, D.; Zhou, Q.; Sun, Y. Eugenol eliminates carbapenem-resistant Klebsiella pneumoniae via reactive oxygen species mechanism. Front. Microbiol. 2023, 14, 1090787. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, S.T.; Bassler, B.L. Bacterial Quorum Sensing: Its Role in Virulence and Possibilities for Its Control. Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef]
- Guerra, M.E.S.; Destro, G.; Vieira, B.; Lima, A.S.; Ferraz, L.F.C.; Hakansson, A.P.; Darrieux, M.; Converso, T.R. Klebsiella pneumoniae Biofilms and Their Role in Disease Pathogenesis. Front. Cell Infect. Microbiol. 2022, 12, 877995. [Google Scholar] [CrossRef] [PubMed]
- Balestrino, D.; Haagensen, J.A.J.; Rich, C.; Forestier, C. Characterization of Type 2 Quorum Sensing in Klebsiella pneumoniae and Relationship with Biofilm Formation. J. Bacteriol. 2005, 187, 2870–2880. [Google Scholar] [CrossRef]
- Naga, N.G.; El-Badan, D.E.; Ghanem, K.M.; Shaaban, M.I. It is the time for quorum sensing inhibition as alternative strategy of antimicrobial therapy. Cell Commun. Signal. 2023, 21, 133. [Google Scholar] [CrossRef]
- Skindersoe, M.E.; Alhede, M.; Phipps, R.; Yang, L.; Jensen, P.O.; Rasmussen, T.B.; Bjarnsholt, T.; Tolker-Nielsen, T.; Høiby, N.; Givskov, M. Effects of Antibiotics on Quorum Sensing in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2008, 52, 3648–3663. [Google Scholar] [CrossRef]
- Carvalho, A.; Krin, E.; Korlowski, C.; Mazel, D.; Baharoglu, Z. Interplay between Sublethal Aminoglycosides and Quorum Sensing: Consequences on Survival in V. cholerae. Cells 2021, 10, 3227. [Google Scholar] [CrossRef] [PubMed]
- De Araujo, C.; Balestrino, D.; Roth, L.; Charbonnel, N.; Forestier, C. Quorum sensing affects biofilm formation through lipopolysaccharide synthesis in Klebsiella pneumoniae. Res. Microbiol. 2010, 161, 595–603. [Google Scholar] [CrossRef]
- Brisse, S.; Passet, V.; Haugaard, A.B.; Babosan, A.; Kassis-Chikhani, N.; Struve, C.; Decré, D. wzi Gene Sequencing, a Rapid Method for Determination of Capsular Type for Klebsiella Strains. J. Clin. Microbiol. 2013, 51, 4073–4078. [Google Scholar] [CrossRef]
- Pacheco, T.; Gomes, A.É.I.; Siqueira, N.M.G.; Assoni, L.; Darrieux, M.; Venter, H.; Ferraz, L.F.C. SdiA, a Quorum-Sensing Regulator, Suppresses Fimbriae Expression, Biofilm Formation, and Quorum-Sensing Signaling Molecules Production in Klebsiella pneumoniae. Front. Microbiol. 2021, 12, 597735. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Chen, J.; Yang, C.; Yin, Y.; Yao, K. Quorum Sensing: A Prospective Therapeutic Target for Bacterial Diseases. BioMed Res. Int. 2019, 2019, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Cadavid, E.; Echeverri, F. The Search for Natural Inhibitors of Biofilm Formation and the Activity of the Autoinductor C6-AHL in Klebsiella pneumoniae ATCC 13884. Biomolecules 2019, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lin, Y.; Zhang, X.; Chen, L.; Xu, C.; Liu, S.; Cao, J.; Zheng, X.; Jia, H.; Chen, L.; et al. Combining Colistin with Furanone C-30 Rescues Colistin Resistance of Gram-Negative Bacteria in Vitro and in Vivo. Microbiol. Spectr. 2021, 9, e01231-21. [Google Scholar] [CrossRef] [PubMed]
- Helmy, Y.A.; Taha-Abdelaziz, K.; Hawwas, H.A.E.H.; Ghosh, S.; AlKafaas, S.S.; Moawad, M.M.M.; Saied, E.M.; Kassem, I.; Mawad, A.M. Antimicrobial Resistance and Recent Alternatives to Antibiotics for the Control of Bacterial Pathogens with an Emphasis on Foodborne Pathogens. Antibiotics 2023, 12, 274. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Gao, Y.; Chen, X.; Yu, Z.; Li, X. Quorum Quenching Enzymes and Their Application in Degrading Signal Molecules to Block Quorum Sensing-Dependent Infection. Int. J. Mol. Sci. 2013, 14, 17477–17500. [Google Scholar] [CrossRef] [PubMed]
- Fong, J.; Zhang, C.; Yang, R.; Boo, Z.Z.; Tan, S.K.; Nielsen, T.E.; Givskov, M.; Liu, X.-W.; Bin, W.; Su, H.; et al. Combination Therapy Strategy of Quorum Quenching Enzyme and Quorum Sensing Inhibitor in Suppressing Multiple Quorum Sensing Pathways of P. aeruginosa. Sci. Rep. 2018, 8, 1155. [Google Scholar] [CrossRef]
- Murugayah, S.A.; Gerth, M.L. Engineering quorum quenching enzymes: Progress and perspectives. Biochem. Soc. Trans. 2019, 47, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Rehman, Z.U.; Momin, A.A.; Aldehaiman, A.; Irum, T.; Grünberg, R.; Arold, S.T. The exceptionally efficient quorum quenching enzyme LrsL suppresses Pseudomonas aeruginosa biofilm production. Front. Microbiol. 2022, 13, 977673. [Google Scholar] [CrossRef]
- Rémy, B.; Plener, L.; Decloquement, P.; Armstrong, N.; Elias, M.; Daudé, D.; Chabrière, É. Lactonase Specificity Is Key to Quorum Quenching in Pseudomonas aeruginosa. Front. Microbiol. 2020, 11, 762. [Google Scholar] [CrossRef]
- Case, R.J.; Labbate, M.; Kjelleberg, S. AHL-driven quorum-sensing circuits: Their frequency and function among the Proteobacteria. ISME J. 2008, 2, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Lade, H.; Paul, D.; Kweon, J.H. N-Acyl Homoserine Lactone-Mediated Quorum Sensing with Special Reference to Use of Quorum Quenching Bacteria in Membrane Biofouling Control. BioMed Res. Int. 2014, 2014, 162584. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zeng, X.; Zheng, J.; Zou, Y.; Qiu, S.; Dai, Y. AHL-mediated quorum sensing to regulate bacterial substance and energy metabolism: A review. Microbiol. Res. 2022, 262, 127102. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, B.; Grenier, D.; Yi, L. Regulatory Mechanisms of the LuxS/AI-2 System and Bacterial Resistance. Antimicrob. Agents Chemother. 2019, 63, e01186-19. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Xu, Y.; Yuan, B.; Yue, Y.; Zhao, M.; Luo, R.; Wu, H.; Wang, L.; Zhang, Y.; Xiao, J.; et al. Effect of Autoinducer-2 Quorum Sensing Inhibitor on Interspecies Quorum Sensing. Front. Microbiol. 2022, 13, 791802. [Google Scholar] [CrossRef] [PubMed]
- Farah, C.; Vera, M.; Morin, D.; Haras, D.; Jerez, C.A.; Guiliani, N. Evidence for a Functional Quorum-Sensing Type AI-1 System in the Extremophilic Bacterium Acidithiobacillus ferrooxidans. Appl. Environ. Microbiol. 2005, 71, 7033–7040. [Google Scholar] [CrossRef]
- Boon, E.M.; Dai, Z.; Liu, N.; Henares, B.; Arora, D.; Lahiri, T. Nitric oxide regulated cyclic di-GMP signaling. BMC Pharmacol. Toxicol. 2013, 14, O37. [Google Scholar] [CrossRef]
- Ramírez-Mata, A.; Fernández-Domínguez, I.J.; Nuñez-Reza, K.J.; Xiqui-Vázquez, M.L.; Baca, B.E. Redes de señalización en la producción de biopelículas en bacterias: Quorum sensing, di-GMPc y óxido nítrico. [Networks involving quorum sensing, cyclic-di-GMP and nitric oxide on biofilm production in bacteria]. Rev. Argent. Microbiol. 2014, 16, 242–255. [Google Scholar]
- Kincses, A.; Rácz, B.; Baaity, Z.; Vásárhelyi, O.; Kristóf, E.; Somogyvári, F.; Spengler, G. The Relationship between Antibiotic Sus-ceptibility and pH in the Case of Uropathogenic Bacteria. Antibiotics 2021, 10, 1431. [Google Scholar] [CrossRef]
- Herrera-Espejo, S.; Fontserè, S.; Infante, C.; Suárez-Benjumea, A.; Carretero-Ledesma, M.; Suñer-Poblet, M.; González-Corvillo, G.; Bernal, G.; Martín-Gutiérrez, G.; Pérez-Cáceres, G.A.; et al. Acidic Urine pH and Clinical Outcome of Lower Urinary Tract Infection in Kidney Transplant Recipients Treated with Ciprofloxa-cin and Fosfomycin. Antibiotics 2024, 13, 116. [Google Scholar] [CrossRef]
- Sorbara, M.T.; Dubin, K.; Littmann, E.R.; Moody, T.U.; Fontana, E.; Seok, R.; Leiner, I.M.; Taur, Y.; Peled, J.U.; van den Brink, M.E.M.; et al. Inhibiting antibiotic-resistant Enterobacte-riaceae by microbiota-mediated intracellular acidification. J. Exp. Med. 2019, 216, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Zarzecka, U.; Chajęcka-Wierzchowska, W.; Zakrzewski, A.; Zadernowska, A.; Fraqueza, M.J. High pressure processing, acidic and osmotic stress increased resistance to aminoglycosides and tetracyclines and the frequency of gene transfer among strains from commercial starter and protective cultures. Food Microbiol. 2022, 107, 104090. [Google Scholar] [CrossRef] [PubMed]
- Deekshit, V.K.; Srikumar, S. ‘To be, or not to be’—The dilemma of ‘silent’ antimicrobial resistance genes in bacteria. J. Appl. Microbiol. 2022, 133, 2902–2914. [Google Scholar] [CrossRef] [PubMed]
- Olaitan, A.O.; Dureja, C.; Youngblom, M.A.; Topf, M.A.; Shen, W.J.; Gonzales-Luna, A.J.; Deshpande, A.; Hevener, K.E.; Freeman, J.; Wilcox, M.H.; et al. Decoding a cryptic mechanism of metronidazole resistance among globally disseminated fluoroquinolone-resistant Clostridioidium difficile. Nat. Commun. 2023, 14, 4130. [Google Scholar] [CrossRef] [PubMed]
- Maddocks, S.E.; Oyston, P.C.F. Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins. Microbiology 2008, 154, 3609–3623. [Google Scholar] [CrossRef] [PubMed]
- Baugh, A.C.; Momany, C.; Neidle, E.L. Versatility and Complexity: Common and Uncommon Facets of LysR-Type Transcriptional Regulators. Annu. Rev. Microbiol. 2023, 77, 317–339. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, V.A.A.; Lery, L.M.S. Insights into Klebsiella pneumoniae type VI secretion system transcriptional regulation. BMC Genom. 2019, 20, 506. [Google Scholar] [CrossRef] [PubMed]
- Cornacchia, A.; Janowicz, A.; Centorotola, G.; Saletti, M.A.; Ranieri, S.C.; Ancora, M.; Ripà, P.; Cammà, C.; Pomilio, F.; Chiaverini, A.; et al. Multi-approach methods to pre-dict cryptic carbapenem resistance mechanisms in Klebsiella pneumoniae detected in Central Italy. Front. Microbiol. 2023, 14, 1242693. [Google Scholar] [CrossRef] [PubMed]
- Henderson, B.; Martin, A. Bacterial Virulence in the Moonlight: Multitasking Bacterial Moonlighting Proteins Are Virulence Determinants in Infectious Disease. Infect. Immun. 2011, 79, 3476–3491. [Google Scholar] [CrossRef]
- Kupper, M.; Gupta, S.K.; Feldhaar, H.; Gross, R. Versatile roles of the chaperonin GroEL in microorganism-insect interac-tions. FEMS Microbiol. Lett. 2014, 353, 1–10. [Google Scholar] [CrossRef]
- Lee, I.P.A.; Eldakar, O.T.; Gogarten, J.P.; Andam, C.P. Bacterial cooperation through horizontal gene transfer. Trends Ecol. Evol. 2022, 37, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Woods, L.C.; Gorrell, R.J.; Taylor, F.; Connallon, T.; Kwok, T.; McDonald, M.J. Horizontal gene transfer potentiates adaptation by reducing selective constraints on the spread of genetic variation. Proc. Natl. Acad. Sci. USA 2020, 117, 26868–26875. [Google Scholar] [CrossRef] [PubMed]
- Ombada, M.; Perwez, T.; Campitelli, M.; Zeineddine, N. Hypermucoviscous Klebsiella pneumoniae: A Hypervirulent Strain Masquerading as Metastasis. Cureus 2023, 15, e39561. Available online: https://www.cureus.com/articles/150203-hypermucoviscous-klebsiella-pneumoniae-a-hypervirulent-strain-masquerading-as-metastasis (accessed on 27 May 2023). [CrossRef] [PubMed]
- Shon, A.S.; Bajwa, R.P.S.; Russo, T.A. Hypervirulent (hypermucoviscous) Klebsiella pneumoniae: A new and dangerous breed. Virulence 2013, 4, 107–118. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Inhibitor | Function | Reference |
|---|---|---|
| Furanones | They inhibit the production of auto-inducers by interrupting signaling, which prevents bacteria from producing and detecting the auto-inducing molecules necessary to coordinate their behavior, reducing cell density-dependent gene expression and that related to virulence. | [125,126] |
| (AHL-ase) | Enzyme that is responsible for degrading autoinduction molecules (AHLs) produced by Gram-negative bacteria and, therefore, their communication. | [125,126,127] |
| Mechanism that involves the enzymatic degradation of signaling molecules used by bacteria to synchronize their behavior within communities. It prevents the accumulation of signaling molecules in the medium, which inhibits QS. It is classified as follows: class I includes lactonases, acylases and paraoxonases. Class II refers to oxidoreductases. | [127,128,129,130,131,132] | |
| Inductor | ||
| (AHLs) | Acyl-homoserine lactones (AHLs) are signaling molecules in the quorum sensing system, which play a role in the coordination of various bacterial processes; Inside the cell, AHLs bind to LuxR-type receptor proteins, forming a transcription activating complex. | [133,134,135] |
| AlQS System | The Auto-Inducer Quorum Sensing System is an intercellular communication mechanism that allows bacteria to coordinate their behavior based on population density. This system involves the synthesis and detection of signaling molecules (auto-inducers), which bind to specific receptors on the bacterial surface when they reach a critical concentration, activating the expression of specific genes that regulate processes such as biofilm production, virulence, etc | [133,134,135] |
| Lux regulators | The Lux-type regulator family is a group of proteins essential for cellular communication and the coordination of events depending on the density of the bacterial population. They are classified based on their structure and function, with LuxR being the main regulator.
| [125,135,136,137,138,139] |
| SdiA | Transcriptional regulator that belongs to the family of LuxR-type regulators, acting by suppressing the expression of fimbriae and biofilm formation, in addition to regulating the expression of genes related to virulence. | [136,139] |
| di-GMPc | Second intracellular messenger that plays a crucial role in regulating biofilm formation. This molecule is involved in signal transduction and controls the expression of genes related to the production of exopolysaccharides and biofilm formation in bacteria. | [125,139,140] |
| Nitric oxide | Signaling molecule that participates in intercellular communication and in the regulation of various physiological processes in bacteria. In the context of quorum sensing, nitric oxide can act as an environmental signal that modulates biofilm formation and bacterial motility in response to specific conditions. | [125,139,140] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivera-Galindo, M.A.; Aguirre-Garrido, F.; Garza-Ramos, U.; Villavicencio-Pulido, J.G.; Fernández Perrino, F.J.; López-Pérez, M. Relevance of the Adjuvant Effect between Cellular Homeostasis and Resistance to Antibiotics in Gram-Negative Bacteria with Pathogenic Capacity: A Study of Klebsiella pneumoniae. Antibiotics 2024, 13, 490. https://doi.org/10.3390/antibiotics13060490
Rivera-Galindo MA, Aguirre-Garrido F, Garza-Ramos U, Villavicencio-Pulido JG, Fernández Perrino FJ, López-Pérez M. Relevance of the Adjuvant Effect between Cellular Homeostasis and Resistance to Antibiotics in Gram-Negative Bacteria with Pathogenic Capacity: A Study of Klebsiella pneumoniae. Antibiotics. 2024; 13(6):490. https://doi.org/10.3390/antibiotics13060490
Chicago/Turabian StyleRivera-Galindo, Mildred Azucena, Félix Aguirre-Garrido, Ulises Garza-Ramos, José Geiser Villavicencio-Pulido, Francisco José Fernández Perrino, and Marcos López-Pérez. 2024. "Relevance of the Adjuvant Effect between Cellular Homeostasis and Resistance to Antibiotics in Gram-Negative Bacteria with Pathogenic Capacity: A Study of Klebsiella pneumoniae" Antibiotics 13, no. 6: 490. https://doi.org/10.3390/antibiotics13060490
APA StyleRivera-Galindo, M. A., Aguirre-Garrido, F., Garza-Ramos, U., Villavicencio-Pulido, J. G., Fernández Perrino, F. J., & López-Pérez, M. (2024). Relevance of the Adjuvant Effect between Cellular Homeostasis and Resistance to Antibiotics in Gram-Negative Bacteria with Pathogenic Capacity: A Study of Klebsiella pneumoniae. Antibiotics, 13(6), 490. https://doi.org/10.3390/antibiotics13060490