Antimicrobial Resistance in Klebsiella pneumoniae Strains: Mechanisms and Outbreaks †



Abstract
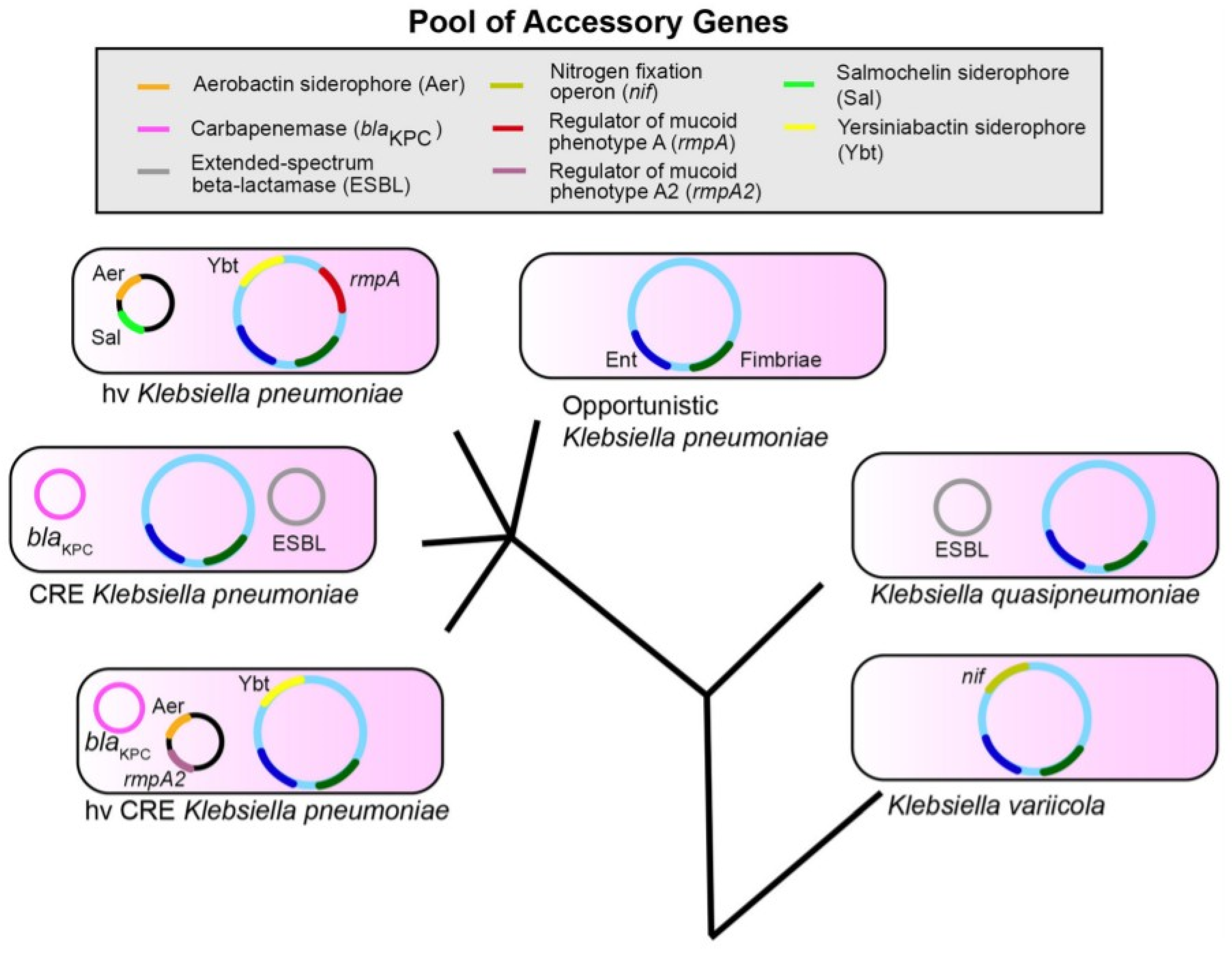
:1. Introduction
2. Main Resistance Mechanisms in K. pneumoniae
- (a)
- Mechanisms of antibiotic resistance to β-lactam antibiotics
- (b) Quinolone resistance
3. Outbreaks Caused by Resistant Strains of K. pneumoniae
4. Conclusions
Author Contributions
Conflicts of Interest
Abbreviations
| ESBL | Extended Spectrum β-Lactamase |
References
- Martin, R.; Bachman, M. Colonization, infection, and the accessory genome of Klebsiella pneumoniae. Front. Cell. Infect. Microbiol. 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.; Holt, K. Klebsiella pneumoniae as a key trafficker of drug resistance genes from environmental to clinically important bacteria. Curr. Opin. Microbiol. 2018, 45, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.; Holt, K. Klebsiella pneumoniae Population Genomics and Antimicrobial-Resistant Clones. Trends Microbiol. 2016, 24, 944–956. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Dortet, L.; Poirel, L. Carbapenem resistance in Enterobacteriaceae: Here is the storm! Trends Mol. Med. 2012, 18, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Kaczanowska, M.; Rydén-Aulin, M. Ribosome biogenesis and the translation process in Escherichia coli. Microbiol. Mol. Biol. Rev. 2007, 71, 477–494. [Google Scholar] [CrossRef] [PubMed]
- Villa, L.; Feudi, C.; Fortini, D.; Garcia-Fernandez, A.; Carattoli, A. Genomics of KPC-producing Klebsiella pneumoniae sequence type 512 clone highlights the role of RamR and ribosomal S10 protein mutations in conferring tigecycline resistance. Antimicrob. Agents Chemother. 2014, 58, 1707–1712. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Kawamura, Y.; Ezaki, T.; Balakrish Nair, G.; Iida, K.I.; Yoshida, S.I. Alteration in the GyrA subunit of DNA gyrase and the ParC subunit of topoisomerase IV in quinolone-resistant Shigella dysenteriae serotype 1 clinical isolates from Kolkata, India. Antimicrob. Agents Chemother. 2005, 49, 1660–1661. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Martinez, L.; Hernandez-Allas, S.; Alberti, S.; Tomas, J.; Benedi, V.; Jacoby, G. In vivo selection of porin-deficient mutants of Klebsiella pneumoniae with increased resistance to cefoxitin and expanded-spectrum cephalosporins. Antimicrob. Agents Chemother. 1996, 40, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.; Pons, M.; Gomes, C. Transferable mechanisms of quinolone resistance. Int. J. Antimicrob. Agents 2012, 40, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Gao, J.; Tang, W. Nosocomial infection and its molecular mechanisms of antibiotic resistance. Biosci. Trends 2016, 10, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.; Harris, P.; Henderson, A.; Paterson, D. Treatment of infections by OXA-48-producing Enterobacteriaceae. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Chen, J.; Xiao, B.; Kang, X.; Lao, X.; Zheng, H. Discovery of NDM-1 inhibitors from natural products. J. Glob. Antimicrob. Resist. 2019, 18, 80–87. [Google Scholar] [CrossRef]
- Weisenberg, S.; Morgan, D.; Espinal-Witter, R.; Larone, D. Clinical outcomes of patients with Klebsiella pneumoniae carbapenemase-producing K. pneumoniae after treatment with imipenem or meropenem. Diagn. Microbiol. Infect. Dis. 2009, 64, 233–235. [Google Scholar] [CrossRef] [PubMed]
- Gasink, L.; Edelstein, P.; Lautenbach, E.; Synnestvedt, M.; Fishman, N. Risk factors and clinical impact of Klebsiella pneumoniae carbapenemase-producing K. pneumoniae. Infect. Control Hosp. Epidemiol. 2009, 30, 1180–1185. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| β-Lactamases | bla genes conferring resistance (* intrinsic) | |
| Class A | CARB-3, PSE-1, SCO-1, SHV-1 *, TEM-1 | |
| - ESBL | CTX-M, SHV-5, TEM-10, VEB | |
| - Carbapenemase | KPC, GES-5 Class B Metallo | |
| - β-lactamase | CphA, IMP, NDM, SIM, VIM Class C | |
| (Cephalosporinase) | AmpC, CMY, DHA, FOX, MIR | |
| Class D | OXA-1, OXA-2, OXA-7, OXA-9, OXA-10, OXA-12 | |
| - ESBL | OXA-11, OXA-15 | |
| Carbapenemase | OXA-48, OXA-51, OXA-181, OXA-237 | |
| Other AMR | Genes conferring resistance (* intrinsic) | Mutations |
| Aminoglycosides | aac, aadA, aadB, aph, armA, rmt, strAB | - |
| Carbapenems | (see carbapenemase bla genes, class A and D above) | Mutations in ompK35, ompK36 |
| Colistin | mcr-1, mcr1.2 | Inactivation of pmrB, mgrB; mutations in crrB Fluoroquinolones |
| oqxAB efflux Macrolides | ereA, ereB, ermB, mef, mph, msrE | - |
| Phenicols | catA, catB, cml, floR | Rifampin |
| Sulfonamides | folP, sul1, sul2, sul3 | - |
| Tetracycline | tet genes | - |
| Tigecycline | - | Upregulation of acrAB or oqxAB efflux; mutation in rpsJ |
| Trimethoprim | dfr genes | - |
| Enzyme | Replicons | No of Plasmids | Genbank References |
|---|---|---|---|
| CMY-8 (CTX-M-3) | HI2 | 1 | 3 |
| CMY-36 | ColE | 1 | 7 |
| CTX-M-3 (ArmA) | L/M | 73 | 11 |
| HI2 | 68 | ||
| CTX-M-9 | P | 10 | 636 |
| I1 | 8 | ||
| CTX-M-14 | FII, FIB | 13 | 11,375 |
| CTX-M-15 [TEM-1, AAC(6′)-IB-CR] | FII, FIA, FIB | 152 | 17 |
| NT | 22 | ||
| CTX-M-17 | ColE | 1 | 5 |
| CTX-M-62 | NT | 1 | 5 |
| SHV-5 | L/M | 118 | 397 |
| FII, FIB | 2 | ||
| I1 | 12 | ||
| SHV-12 | K | 5 | 1189 |
| FII, FIB | 9 | ||
| FIIK | 1 | ||
| TEM-1 | ColE | 2 | 67,442 |
| KPC-2 | N | 1 | 29,077 |
| ColE | 1 | ||
| IMP-4 (QnrB2, QnrB-4, ArmA) | L/M | 22 | 1888 |
| A/C | 15 | ||
| I1 | 17 | ||
| VIM-1 | N | 15 | 2379 |
| NT | 1 | ||
| VIM-4 (CMY-4) ArmA | A/C | 2 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moya, C.; Maicas, S. Antimicrobial Resistance in Klebsiella pneumoniae Strains: Mechanisms and Outbreaks. Proceedings 2020, 66, 11. https://doi.org/10.3390/proceedings2020066011
Moya C, Maicas S. Antimicrobial Resistance in Klebsiella pneumoniae Strains: Mechanisms and Outbreaks. Proceedings. 2020; 66(1):11. https://doi.org/10.3390/proceedings2020066011
Chicago/Turabian StyleMoya, Celeste, and Sergi Maicas. 2020. "Antimicrobial Resistance in Klebsiella pneumoniae Strains: Mechanisms and Outbreaks" Proceedings 66, no. 1: 11. https://doi.org/10.3390/proceedings2020066011
APA StyleMoya, C., & Maicas, S. (2020). Antimicrobial Resistance in Klebsiella pneumoniae Strains: Mechanisms and Outbreaks. Proceedings, 66(1), 11. https://doi.org/10.3390/proceedings2020066011