
Long-Term Co-Circulation of Host-Specialist and Host-Generalist Lineages of Group B Streptococcus in Brazilian Dairy Cattle with Heterogeneous Antimicrobial Resistance Profiles

 , ,
, ,  and
and {kind=link}
{kind=link}
Abstract
1. Introduction
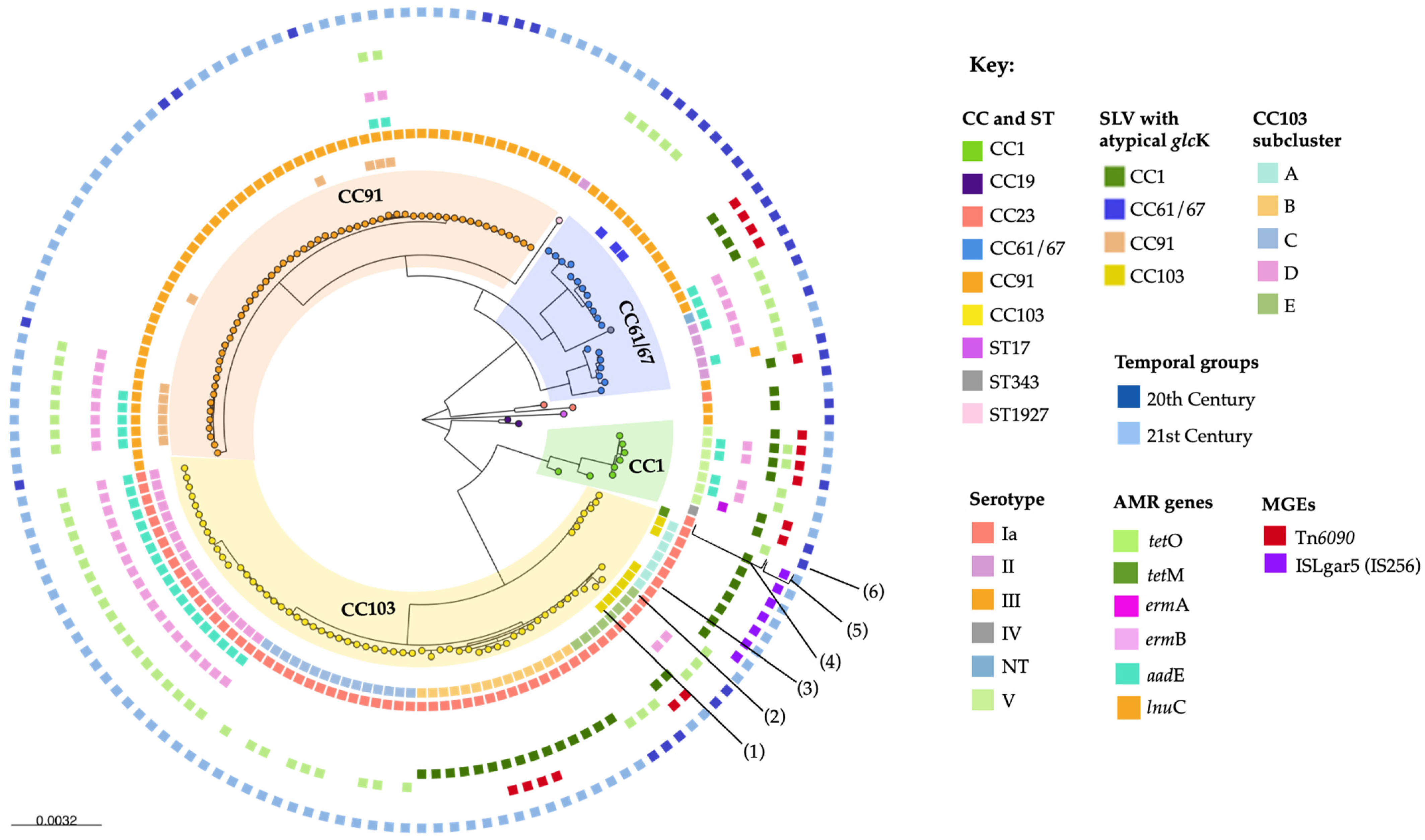
2. Results
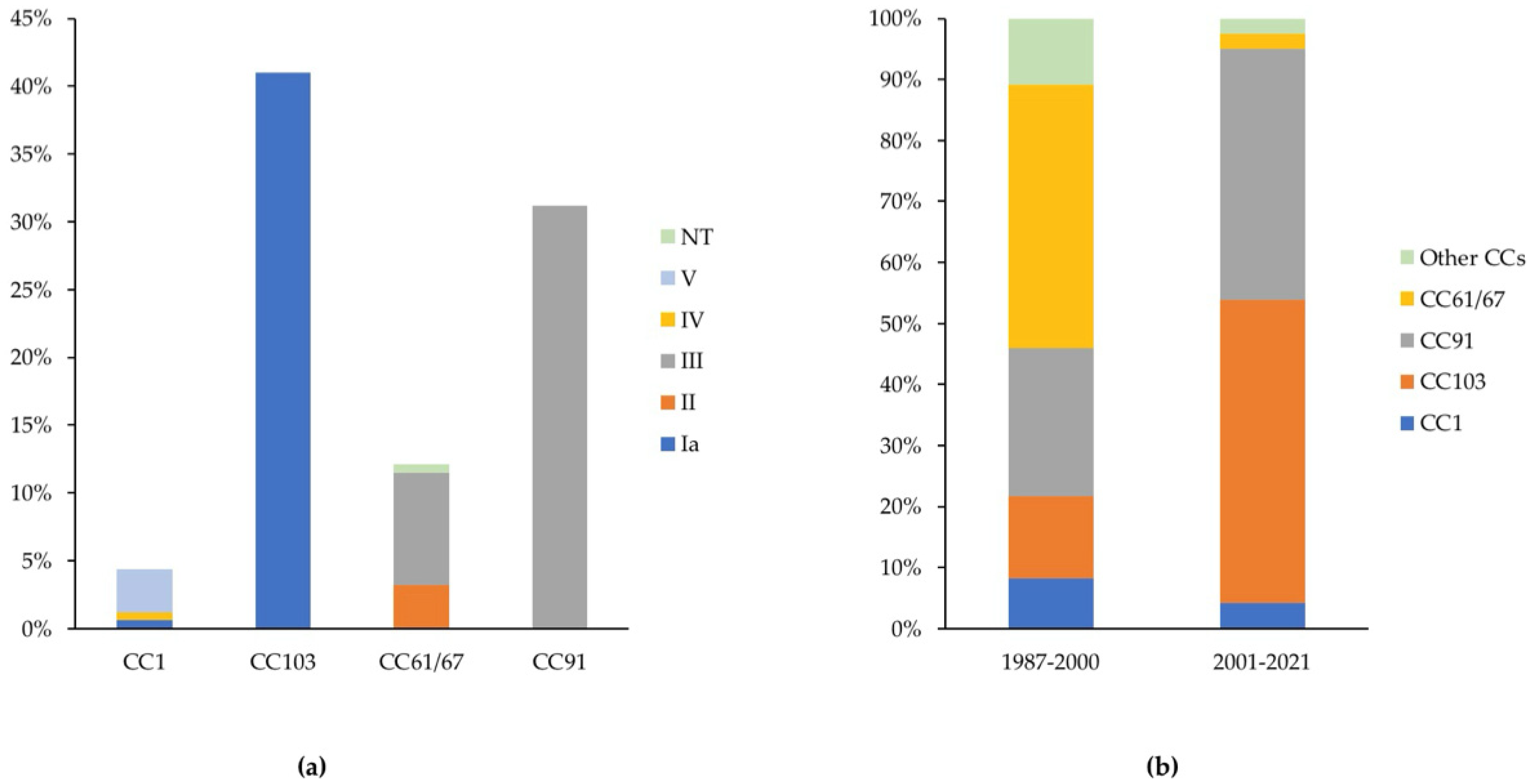
2.1. Descriptive Analysis
2.2. Within-Herd Analysis
2.3. Between-Herd and Temporal Analyses
2.4. Antimicrobial Resistance
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates
4.2. Phenotypic Analysis
4.3. Genomic and Phylogenetic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Russell, N.J.; Seale, A.C.; O’Driscoll, M.; O’Sullivan, C.; Bianchi-Jassir, F.; Gonzalez-Guarin, J.; Lawn, J.E.; Baker, C.J.; Bartlett, L.; Cutland, C.; et al. Maternal Colonization with Group B Streptococcus and Serotype Distribution Worldwide: Systematic Review and Meta-Analyses. Clin. Infect. Dis. 2017, 65, S100–S111. [Google Scholar] [CrossRef] [PubMed]
- Leal, C.A.G.; Queiroz, G.A.; Pereira, F.L.; Tavares, G.C.; Figueiredo, H.C.P. Streptococcus agalactiae Sequence Type 283 in Farmed Fish, Brazil. Emerg. Infect. Dis. 2019, 25, 776–779. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, Y.; Ding, Y.; Yi, L.; Ma, Z.; Fan, H.; Lu, C. Molecular Characterization of Streptococcus agalactiae Isolated from Bovine Mastitis in Eastern China. PLoS ONE 2013, 8, e67755. [Google Scholar] [CrossRef] [PubMed]
- Group B Streptococcus Vaccine: Full Value Vaccine Assessment. Available online: https://www.who.int/publications-detail-redirect/9789240037526 (accessed on 27 March 2024).
- Hall, J.; Adams, N.H.; Bartlett, L.; Seale, A.C.; Lamagni, T.; Bianchi-Jassir, F.; Lawn, J.E.; Baker, C.J.; Cutland, C.; Heath, P.T.; et al. Maternal Disease with Group B Streptococcus and Serotype Distribution Worldwide: Systematic Review and Meta-Analyses. Clin. Infect. Dis. 2017, 65, S112–S124. [Google Scholar] [CrossRef] [PubMed]
- Francois Watkins, L.K.; McGee, L.; Schrag, S.J.; Beall, B.; Jain, J.H.; Pondo, T.; Farley, M.M.; Harrison, L.H.; Zansky, S.M.; Baumbach, J.; et al. Epidemiology of Invasive Group B Streptococcal Infections Among Nonpregnant Adults in the United States, 2008–2016. JAMA Intern. Med. 2019, 179, 479. [Google Scholar] [CrossRef] [PubMed]
- Kalimuddin, S.; Chen, S.L.; Lim, C.T.K.; Koh, T.H.; Tan, T.Y.; Kam, M.; Wong, C.W.; Mehershahi, K.S.; Chau, M.L.; Ng, L.C.; et al. 2015 Epidemic of Severe Streptococcus agalactiae Sequence Type 283 Infections in Singapore Associated with the Consumption of Raw Freshwater Fish: A Detailed Analysis of Clinical, Epidemiological, and Bacterial Sequencing Data. Clin. Infect. Dis. 2017, 15, S145–S152. [Google Scholar] [CrossRef] [PubMed]
- Cobo-Angel, C.G.; Jaramillo-Jaramillo, A.S.; Palacio-Aguilera, M.; Jurado-Vargas, L.; Calvo-Villegas, E.A.; Ospina-Loaiza, D.A.; Rodriguez-Lecompte, J.C.; Sanchez, J.; Zadoks, R.; Ceballos-Marquez, A. Potential Group B Streptococcus Interspecies Transmission between Cattle and People in Colombian Dairy Farms. Sci. Rep. 2019, 9, 14025. [Google Scholar] [CrossRef] [PubMed]
- Zadoks, R.N.; Middleton, J.R.; McDougall, S.; Katholm, J.; Schukken, Y.H. Molecular Epidemiology of Mastitis Pathogens of Dairy Cattle and Comparative Relevance to Humans. J. Mammary Gland. Biol. Neoplasia 2011, 16, 357–372. [Google Scholar] [CrossRef]
- Botelho, A.C.N.; Ferreira, A.F.M.; Fracalanzza, S.E.L.; Teixeira, L.M.; Pinto, T.C.A. A Perspective on the Potential Zoonotic Role of Streptococcus agalactiae: Searching for a Missing Link in Alternative Transmission Routes. Front. Microbiol. 2018, 9, 608. [Google Scholar] [CrossRef]
- Crestani, C.; Forde, T.L.; Lycett, S.J.; Holmes, M.A.; Fasth, C.; Persson-Waller, K.; Zadoks, R.N. The Fall and Rise of Group B Streptococcus in Dairy Cattle: Reintroduction Due to Human-to-Cattle Host Jumps? Microb. Genom. 2021, 7, 000648. [Google Scholar] [CrossRef]
- Lyhs, U.; Kulkas, L.; Katholm, J.; Waller, K.P.; Saha, K.; Tomusk, R.J.; Zadoks, R.N. Streptococcus agalactiae Serotype IV in Humans and Cattle, Northern Europe1. Emerg. Infect. Dis. 2016, 22, 2097–2103. [Google Scholar] [CrossRef]
- Ruegg, P.L. A 100-Year Review: Mastitis Detection, Management, and Prevention. J. Dairy Sci. 2017, 100, 10381–10397. [Google Scholar] [CrossRef] [PubMed]
- Fifth OIE Annual Report on Antimicrobial Agents Intended for Use in Animals. Available online: https://www.woah.org/en/document/fifth-oie-annual-report-on-antimicrobial-agents-intended-for-use-in-animals/ (accessed on 27 March 2024).
- Pinto, T.C.A.; Costa, N.S.; Vianna Souza, A.R.; Da Silva, L.G.; Corrêa, A.B.D.A.; Fernandes, F.G.; Oliveira, I.C.M.; De Mattos, M.C.; Rosado, A.S.; Benchetrit, L.C. Distribution of Serotypes and Evaluation of Antimicrobial Susceptibility among Human and Bovine Streptococcus agalactiae Strains Isolated in Brazil between 1980 and 2006. Braz. J. Infect. Dis. 2013, 17, 131–136. [Google Scholar] [CrossRef]
- Nanduri, S.A.; Petit, S.; Smelser, C.; Apostol, M.; Alden, N.B.; Harrison, L.H.; Lynfield, R.; Vagnone, P.S.; Burzlaff, K.; Spina, N.L.; et al. Epidemiology of Invasive Early-Onset and Late-Onset Group B Streptococcal Disease in the United States, 2006 to 2015: Multistate Laboratory and Population-Based Surveillance. JAMA Pediatr. 2019, 173, 224. [Google Scholar] [CrossRef]
- Tomazi, T.; De Souza Filho, A.F.; Heinemann, M.B.; Santos, M.V.D. Molecular Characterization and Antimicrobial Susceptibility Pattern of Streptococcus agalactiae Isolated from Clinical Mastitis in Dairy Cattle. PLoS ONE 2018, 13, e0199561. [Google Scholar] [CrossRef]
- CDC. The Biggest Antibiotic-Resistant Threats in the U.S. Available online: https://www.cdc.gov/drugresistance/biggest-threats.html (accessed on 27 March 2024).
- Hernando-Amado, S.; Coque, T.M.; Baquero, F.; Martínez, J.L. Antibiotic Resistance: Moving From Individual Health Norms to Social Norms in One Health and Global Health. Front. Microbiol. 2020, 11, 1914. [Google Scholar] [CrossRef]
- Rainard, P.; Gilbert, F.B.; Germon, P.; Foucras, G. Invited Review: A Critical Appraisal of Mastitis Vaccines for Dairy Cows. J. Dairy Sci. 2021, 104, 10427–10448. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.; Alves-Barroco, C.; Sauvage, E.; Bexiga, R.; Albuquerque, P.; Tavares, F.; Santos-Sanches, I.; Glaser, P. Persistence of a Dominant Bovine Lineage of Group B Streptococcus Reveals Genomic Signatures of Host Adaptation. Environ. Microbiol. 2016, 18, 4216–4229. [Google Scholar] [CrossRef]
- Bisharat, N.; Crook, D.W.; Leigh, J.; Harding, R.M.; Ward, P.N.; Coffey, T.J.; Maiden, M.C.; Peto, T.; Jones, N. Hyperinvasive Neonatal Group B Streptococcus Has Arisen from a Bovine Ancestor. J. Clin. Microbiol. 2004, 42, 2161–2167. [Google Scholar] [CrossRef]
- Jones, N.; Bohnsack, J.F.; Takahashi, S.; Oliver, K.A.; Chan, M.-S.; Kunst, F.; Glaser, P.; Rusniok, C.; Crook, D.W.M.; Harding, R.M.; et al. Multilocus Sequence Typing System for Group B Streptococcus. J. Clin. Microbiol. 2003, 41, 2530–2536. [Google Scholar] [CrossRef]
- Van Belkum, A.; Tassios, P.T.; Dijkshoorn, L.; Haeggman, S.; Cookson, B.; Fry, N.K.; Fussing, V.; Green, J.; Feil, E.; Gerner-Smidt, P.; et al. Guidelines for the Validation and Application of Typing Methods for Use in Bacterial Epidemiology. Clin. Microbiol. Infect. 2007, 13, 1–46. [Google Scholar] [CrossRef]
- M100 Ed34|Performance Standards for Antimicrobial Susceptibility Testing, 34th Edition. Available online: https://clsi.org/standards/products/microbiology/documents/m100/ (accessed on 27 March 2024).
- VET01SEd7|Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated From Animals, 7th Edition. Available online: https://clsi.org/standards/products/veterinary-medicine/documents/vet01s/ (accessed on 27 March 2024).
- Sørensen, U.B.S.; Klaas, I.C.; Boes, J.; Farre, M. The Distribution of Clones of Streptococcus agalactiae (Group B Streptococci) among Herdspersons and Dairy Cows Demonstrates Lack of Host Specificity for Some Lineages. Vet. Microbiol. 2019, 235, 71–79. [Google Scholar] [CrossRef]
- Da Cunha, V.; Davies, M.R.; Douarre, P.-E.; Rosinski-Chupin, I.; Margarit, I.; Spinali, S.; Perkins, T.; Lechat, P.; Dmytruk, N.; Sauvage, E.; et al. Streptococcus agalactiae Clones Infecting Humans Were Selected and Fixed through the Extensive Use of Tetracycline. Nat. Commun. 2014, 5, 4544. [Google Scholar] [CrossRef]
- Dogan, B.; Schukken, Y.H.; Santisteban, C.; Boor, K.J. Distribution of Serotypes and Antimicrobial Resistance Genes among Streptococcus agalactiae Isolates from Bovine and Human Hosts. J. Clin. Microbiol. 2005, 43, 5899–5906. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, L.; Bottini, E.; Cadona, J.; Cacciato, C.; Monteavaro, C.; Bustamante, A.; Sanso, A.M. Multidrug Resistance and Molecular Characterization of Streptococcus agalactiae Isolates From Dairy Cattle with Mastitis. Front. Cell. Infect. Microbiol. 2021, 11, 647324. [Google Scholar] [CrossRef]
- Tian, X.Y.; Zheng, N.; Han, R.W.; Ho, H.; Wang, J.; Wang, Y.T.; Wang, S.Q.; Li, H.G.; Liu, H.W.; Yu, Z.N. Antimicrobial Resistance and Virulence Genes of Streptococcus Isolated from Dairy Cows with Mastitis in China. Microb. Pathog. 2019, 131, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Shao, W.; Wang, F.; Ma, J.; Chen, H.; Wang, S.; Wu, Y.; Wang, C.; Zheng, N.; Wang, J.; et al. Antimicrobial Resistance and Virulence Genes of Streptococcus agalactiae Isolated from Mastitis Milk Samples in China. J. Vet. Res. 2022, 66, 581–590. [Google Scholar] [CrossRef]
- Khan, U.B.; Portal, E.A.R.; Sands, K.; Lo, S.; Chalker, V.J.; Jauneikaite, E.; Spiller, O.B. Genomic Analysis Reveals New Integrative Conjugal Elements and Transposons in GBS Conferring Antimicrobial Resistance. Antibiotics 2023, 12, 544. [Google Scholar] [CrossRef]
- Zhi, Y.; Ji, H.J.; Jung, J.H.; Byun, E.B.; Kim, W.S.; Lin, S.M.; Lim, S.; Jang, A.-Y.; Choi, M.J.; Ahn, K.B.; et al. Molecular Characteristics of IS 1216 Carrying Multidrug Resistance Gene Cluster in Serotype III/Sequence Type 19 Group B Streptococcus. mSphere 2021, 6, e00543-21. [Google Scholar] [CrossRef]
- Zhou, K.; Zhu, D.; Tao, Y.; Xie, L.; Han, L.; Zhang, Y.; Sun, J. New Genetic Context of Lnu(B) Composed of Two Multi-Resistance Gene Clusters in Clinical Streptococcus agalactiae ST-19 Strains. Antimicrob. Resist. Infect. Control 2019, 8, 117. [Google Scholar] [CrossRef]
- Naranjo-Lucena, A.; Slowey, R. Invited Review: Antimicrobial Resistance in Bovine Mastitis Pathogens: A Review of Genetic Determinants and Prevalence of Resistance in European Countries. J. Dairy Sci. 2023, 106, 1–23. [Google Scholar] [CrossRef]
- Gao, J.; Yu, F.-Q.; Luo, L.-P.; He, J.-Z.; Hou, R.-G.; Zhang, H.-Q.; Li, S.-M.; Su, J.-L.; Han, B. Antibiotic Resistance of Streptococcus agalactiae from Cows with Mastitis. Vet. J. 2012, 194, 423–424. [Google Scholar] [CrossRef]
- Carvalho-Castro, G.A.; Silva, J.R.; Paiva, L.V.; Custódio, D.A.C.; Moreira, R.O.; Mian, G.F.; Prado, I.A.; Chalfun-Junior, A.; Costa, G.M. Molecular Epidemiology of Streptococcus agalactiae Isolated from Mastitis in Brazilian Dairy Herds. Braz. J. Microbiol. 2017, 48, 551–559. [Google Scholar] [CrossRef]
- Trauner, A.; Banaei-Esfahani, A.; Gygli, S.M.; Warmer, P.; Feldmann, J.; Zampieri, M.; Borrell, S.; Collins, B.C.; Beisel, C.; Aebersold, R.; et al. Expression Dysregulation as a Mediator of Fitness Costs in Antibiotic Resistance. Antimicrob. Agents Chemother. 2021, 65, e0050421. [Google Scholar] [CrossRef]
- Barsi, F.; Carra, E.; Ricchi, M.; Gnali, G.; Pisoni, G.; Russo, S.; Filippi, A.; Arrigoni, N.; Zadoks, R.N.; Garbarino, C. Circulation of Streptococcus agalactiae ST103 in a Free Stall Italian Dairy Farm. Appl. Environ. Microbiol. 2022, 88, e00383-22. [Google Scholar] [CrossRef]
- Mahmmod, Y.S.; Klaas, I.C.; Katholm, J.; Lutton, M.; Zadoks, R.N. Molecular Epidemiology and Strain-Specific Characteristics of Streptococcus agalactiae at the Herd and Cow Level. J. Dairy Sci. 2015, 98, 6913–6924. [Google Scholar] [CrossRef]
- Reyes, J.; Chaffer, M.; Sanchez, J.; Torres, G.; Macias, D.; Jaramillo, M.; Duque, P.C.; Ceballos, A.; Keefe, G.P. Evaluation of the Efficacy of Intramuscular versus Intramammary Treatment of Subclinical Streptococcus agalactiae Mastitis in Dairy Cows in Colombia. J. Dairy Sci. 2015, 98, 5294–5303. [Google Scholar] [CrossRef]
- Bianchi-Jassir, F.; Paul, P.; To, K.-N.; Carreras-Abad, C.; Seale, A.C.; Jauneikaite, E.; Madhi, S.A.; Russell, N.J.; Hall, J.; Madrid, L.; et al. Systematic Review of Group B Streptococcal Capsular Types, Sequence Types and Surface Proteins as Potential Vaccine Candidates. Vaccine 2020, 38, 6682–6694. [Google Scholar] [CrossRef]
- Neemuchwala, A.; Teatero, S.; Athey, T.B.T.; McGeer, A.; Fittipaldi, N. Capsular Switching and Other Large-Scale Recombination Events in Invasive Sequence Type 1 Group B Streptococcus. Emerg. Infect. Dis. 2016, 22, 1941–1944. [Google Scholar] [CrossRef]
- McGee, L.; Chochua, S.; Li, Z.; Mathis, S.; Rivers, J.; Metcalf, B.; Ryan, A.; Alden, N.; Farley, M.M.; Harrison, L.H.; et al. Multistate, Population-Based Distributions of Candidate Vaccine Targets, Clonal Complexes, and Resistance Features of Invasive Group B Streptococci within the United States, 2015–2017. Clin. Infect. Dis. 2021, 72, 1004–1013. [Google Scholar] [CrossRef]
- Martins, E.R.; Melo-Cristino, J.; Ramirez, M. Reevaluating the Serotype II Capsular Locus of Streptococcus agalactiae. J. Clin. Microbiol. 2007, 45, 3384–3386. [Google Scholar] [CrossRef]
- Usein, C.-R.; Militaru, M.; Cristea, V.; Străuţ, M. Genetic Diversity and Antimicrobial Resistance in Streptococcus agalactiae Strains Recovered from Female Carriers in the Bucharest Area. Mem. Inst. Oswaldo Cruz 2014, 109, 189–196. [Google Scholar] [CrossRef]
- Barony, G.M.; Tavares, G.C.; Pereira, F.L.; Carvalho, A.F.; Dorella, F.A.; Leal, C.A.G.; Figueiredo, H.C.P. Large-Scale Genomic Analyses Reveal the Population Structure and Evolutionary Trends of Streptococcus agalactiae Strains in Brazilian Fish Farms. Sci. Rep. 2017, 7, 13538. [Google Scholar] [CrossRef]
- Delannoy, C.M.; Crumlish, M.; Fontaine, M.C.; Pollock, J.; Foster, G.; Dagleish, M.P.; Turnbull, J.F.; Zadoks, R.N. Human Streptococcus agalactiae Strains in Aquatic Mammals and Fish. BMC Microbiol. 2013, 13, 41. [Google Scholar] [CrossRef]
- Spellerberg, B.; Brandt, C. Streptococcus. In Manual of Clinical Microbiology; American Society for Microbiology: Washington, DC, USA, 2023; Volume 1, pp. 383–402. [Google Scholar]
- Clark, A.E.; Kaleta, E.J.; Arora, A.; Wolk, D.M. Matrix-assisted laser desorption ionization-time of flight mass spectrometry: A fundamental shift in the routine practice of clinical microbiology. Clin. Microbiol. Rev. 2013, 26, 547–603. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Katoh, K. MAFFT: A Novel Method for Rapid Multiple Sequence Alignment Based on Fast Fourier Transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Tonkin-Hill, G.; MacAlasdair, N.; Ruis, C.; Weimann, A.; Horesh, G.; Lees, J.A.; Gladstone, R.A.; Lo, S.; Beaudoin, C.; Floto, R.A.; et al. Producing Polished Prokaryotic Pangenomes with the Panaroo Pipeline. Genome Biol. 2020, 21, 180. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Argimón, S.; Abudahab, K.; Goater, R.J.E.; Fedosejev, A.; Bhai, J.; Glasner, C.; Feil, E.J.; Holden, M.T.G.; Yeats, C.A.; Grundmann, H.; et al. Microreact: Visualizing and Sharing Data for Genomic Epidemiology and Phylogeography. Microb. Genom. 2016, 2, 1–11. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, L.M.A.d.; Simões, L.C.; Crestani, C.; Costa, N.S.; Pantoja, J.C.d.F.; Rabello, R.F.; Teixeira, L.M.; Khan, U.B.; Bentley, S.; Jamrozy, D.; et al. Long-Term Co-Circulation of Host-Specialist and Host-Generalist Lineages of Group B Streptococcus in Brazilian Dairy Cattle with Heterogeneous Antimicrobial Resistance Profiles. Antibiotics 2024, 13, 389. https://doi.org/10.3390/antibiotics13050389
Oliveira LMAd, Simões LC, Crestani C, Costa NS, Pantoja JCdF, Rabello RF, Teixeira LM, Khan UB, Bentley S, Jamrozy D, et al. Long-Term Co-Circulation of Host-Specialist and Host-Generalist Lineages of Group B Streptococcus in Brazilian Dairy Cattle with Heterogeneous Antimicrobial Resistance Profiles. Antibiotics. 2024; 13(5):389. https://doi.org/10.3390/antibiotics13050389
Chicago/Turabian StyleOliveira, Laura Maria Andrade de, Leandro Correia Simões, Chiara Crestani, Natália Silva Costa, José Carlos de Figueiredo Pantoja, Renata Fernandes Rabello, Lucia Martins Teixeira, Uzma Basit Khan, Stephen Bentley, Dorota Jamrozy, and et al. 2024. "Long-Term Co-Circulation of Host-Specialist and Host-Generalist Lineages of Group B Streptococcus in Brazilian Dairy Cattle with Heterogeneous Antimicrobial Resistance Profiles" Antibiotics 13, no. 5: 389. https://doi.org/10.3390/antibiotics13050389
APA StyleOliveira, L. M. A. d., Simões, L. C., Crestani, C., Costa, N. S., Pantoja, J. C. d. F., Rabello, R. F., Teixeira, L. M., Khan, U. B., Bentley, S., Jamrozy, D., Pinto, T. d. C. A., & Zadoks, R. N. (2024). Long-Term Co-Circulation of Host-Specialist and Host-Generalist Lineages of Group B Streptococcus in Brazilian Dairy Cattle with Heterogeneous Antimicrobial Resistance Profiles. Antibiotics, 13(5), 389. https://doi.org/10.3390/antibiotics13050389